


Effects of Phage Cocktail, Probiotics, and Their Combination on Growth Performance and Gut Microbiota of Broiler Chickens

 , , ,
, , ,
Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Probiotics and Phage Preparation
2.2. Chicken Management
2.3. Experimental Design, Animals, and Diets
2.4. Chemical Analysis of Experimental Diets
2.5. Sampling
2.6. Growth Performance
2.7. Chicken Gut Microbiota Study
2.7.1. DNA Extraction
2.7.2. Illumina Sequencing of the V3–V4 Region of the 16S rRNA Gene
2.7.3. Bioinformatics Analysis
2.7.4. Microbial Predicted Functional Metagenomes Based on PICRUSt and STAMP
2.7.5. Nucleotide Sequence Accession Numbers
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
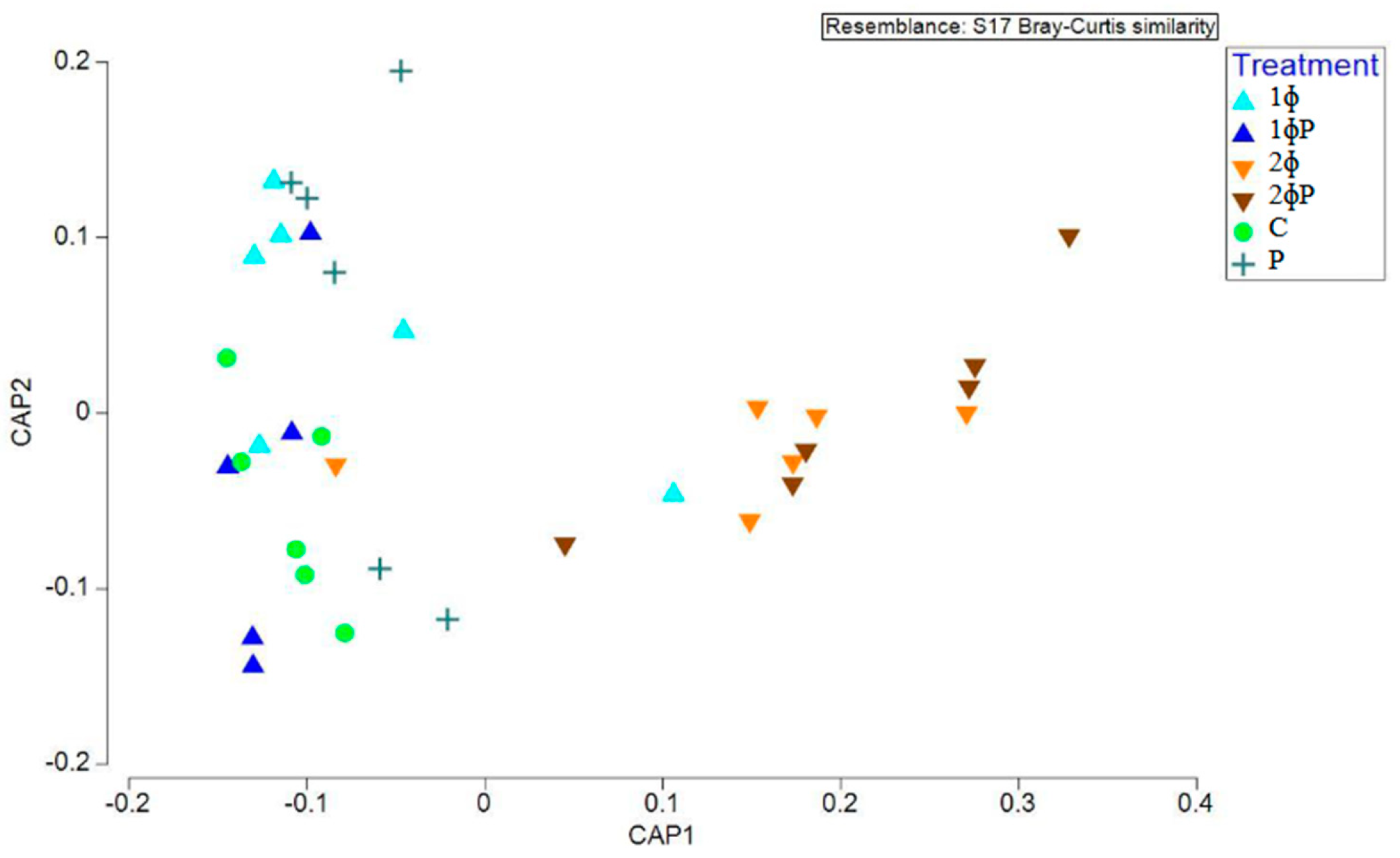
3.2. Multivariate Analysis on Gut Microbiota
3.3. Significant OTUs Present in the ϕP Compared to the Non-ϕP Groups
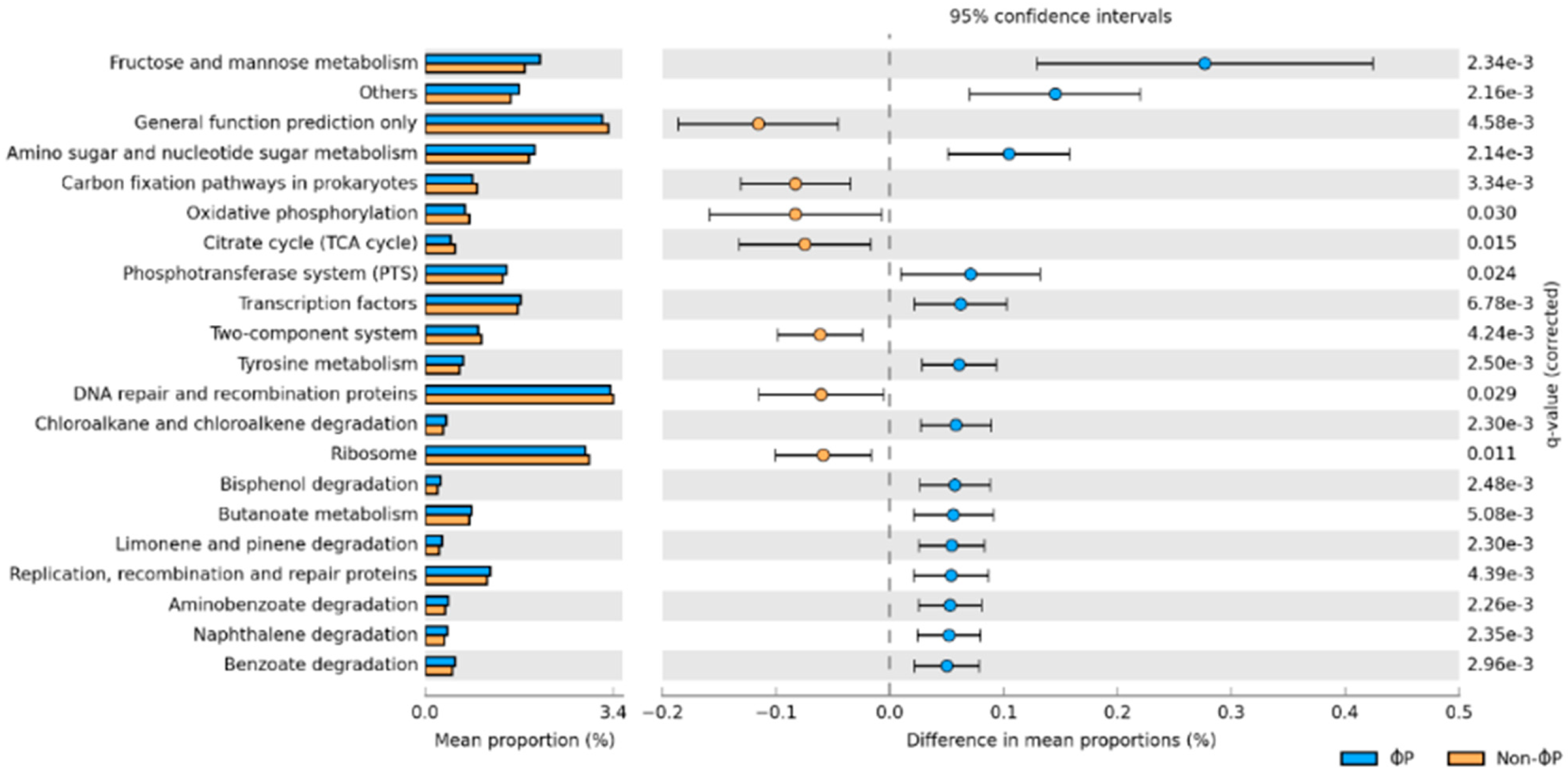
3.4. Microbial Predicted Functional Metagenomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leeson, S. Predictions for Commercial Poultry Nutrition. J. Appl. Poult. Res. 2008, 17, 315–322. [Google Scholar] [CrossRef]
- Dibner, J.J.; Richards, J.D. Antibiotic growth promoters in agriculture: History and mode of action. Poult. Sci. 2005, 84, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, Y.; Letourneau-Montminy, M.P.; Gaucher, M.L.; Chorfi, Y.; Suresh, G.; Rouissi, T.; Brar, S.K.; Cote, C.; Ramirez, A.A.; Godbout, S. Use of antibiotics in broiler production: Global impacts and alternatives. Anim. Nutr. 2018, 4, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Dunshea, F.R.; D’Souza, D.N.; Jensen, B.B.; Engberg, R.M. Meat, Animal, Poultry and Fish Production and Management|Antibiotic Growth Promotants. In Encyclopedia of Meat Sciences; Academic Press: Oxford, UK, 2014; pp. 172–176. [Google Scholar]
- Nhung, N.T.; Chansiripornchai, N.; Carrique-Mas, J.J. Antimicrobial Resistance in Bacterial Poultry Pathogens: A Review. Front. Vet. Sci. 2017, 4, 126. [Google Scholar] [CrossRef] [PubMed]
- Flynn, D. South Korea Bans Antibiotics in Animal Feed. Seattle Food Safety News, 7 June 2011. Available online: https://www.foodsafetynews.com/2011/06/south-korea-bans-antibiotics-in-animal-feed/ (accessed on 2 April 2023).
- Maron, D.F.; Smith, T.J.; Nachman, K.E. Restrictions on antimicrobial use in food animal production: An international regulatory and economic survey. Glob. Health 2013, 9, 1–11. [Google Scholar] [CrossRef]
- McEwen, S.A.; Angulo, F.J.; Collignon, P.J.; Conly, J. Potential unintended consequences associated with restrictions on antimicrobial use in food-producing animals. In WHO Guidelines on Use of Medically Important Antimicrobials in Food-Producing Animals; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- M’Sadeq, S.A.; Wu, S.; Swick, R.A.; Choct, M. Towards the control of necrotic enteritis in broiler chickens with in-feed antibiotics phasing-out worldwide. Anim. Nutr. 2015, 1, 1–11. [Google Scholar] [CrossRef]
- Karavolias, J.; Salois, M.J.; Baker, K.T.; Watkins, K. Raised without antibiotics: Impact on animal welfare and implications for food policy. Transl. Anim. Sci. 2018, 2, 337–348. [Google Scholar] [CrossRef]
- Kim, K.H.; Ingale, S.L.; Kim, J.S.; Lee, S.H.; Lee, J.H.; Kwon, I.K.; Chae, B.J. Bacteriophage and probiotics both enhance the performance of growing pigs but bacteriophage are more effective. Anim. Feed Sci. Technol. 2014, 196, 88–95. [Google Scholar] [CrossRef]
- Zhao, P.Y.; Baek, H.Y.; Kim, I.H. Effects of bacteriophage supplementation on egg performance, egg quality, excreta microflora, and moisture content in laying hens. Asian-Aust. J. Anim. Sci. 2012, 25, 1015–1020. [Google Scholar] [CrossRef]
- Yan, L.; Hong, S.M.; Kim, I.H. Effect of bacteriophage supplementation on the growth performance, nutrient digestibility, blood characteristics, and fecal microbial shedding in growing pigs. Asian-Aust. J. Anim. Sci. 2012, 25, 1451–1456. [Google Scholar] [CrossRef]
- Wang, J.P.; Yan, L.; Lee, J.H.; Kim, I.H. Evaluation of bacteriophage supplementation on growth performance, blood characteristics, relative organ weight, breast muscle characteristics and excreta microbial shedding in broilers. Asian-Aust. J. Anim. Sci. 2013, 26, 573–578. [Google Scholar] [CrossRef]
- Kim, J.S.; Hosseindoust, A.; Lee, S.H.; Choi, Y.H.; Kim, M.J.; Lee, J.H.; Kwon, I.K.; Chae, B.J. Bacteriophage cocktail and multi-strain probiotics in the feed for weanling pigs: Effects on intestine morphology and targeted intestinal coliforms and Clostridium. Animal 2017, 11, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Ngu, N.; Phuong, L.; Anh, L.; Loc, H.; Tam, N.; Huan, P.; Diep, T.; Kamei, K. The Efficiency of Bacteriophages Against Salmonella Typhimurium Infection in Native Noi Broilers. Braz. J. Poult. Sci. 2022, 24, 3. [Google Scholar] [CrossRef]
- Kabir, S. The role of probiotics in the poultry industry. Int. J. Mol. Sci. 2009, 10, 3531–3546. [Google Scholar] [CrossRef] [PubMed]
- De Vuyst, L.; Avonts, L.; Makras, L. Probiotics, prebiotics and gut health. In Woodhead Publishing Series in Food Science, Technology and Nutrition, Functional Foods, Ageing and Degenerative Disease; Woodhead Publishing: Cambridge, UK, 2004; pp. 416–482. [Google Scholar]
- Mountzouris, K.C.; Tsitrsikos, P.; Palamidi, I.; Arvaniti, A.; Mohnl, M.; Schatzmayr, G.; Fegeros, K. Effects of probiotic inclusion levels in broiler nutrition on growth performance, nutrient digestibility, plasma immunoglobulins, and cecal microflora composition. Poult. Sci. 2010, 89, 58–67. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Li, Q.; Zeng, X.; Xu, Y.; Jin, K.; Liu, J.; Cao, G. Effects of Probiotics on the Growth Performance, Antioxidant Functions, Immune Responses, and Caecal Microbiota of Broilers Challenged by Lipopolysaccharide. Front. Vet. Sci. 2022, 9, 846649. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Li, Z.; Wang, P.; Zhu, B.; Zhao, M.; Huang, D.; Ye, Y.; Ding, Z.; Li, L.; Wan, G.; et al. Effects of probiotic supplements on growth performance and intestinal microbiota of partridge shank broiler chicks. PeerJ 2021, 9, e12538. [Google Scholar] [CrossRef]
- Blajman, J.E.; Frizzo, L.S.; Zbrun, M.V.; Astesana, D.M.; Fusari, M.L.; Soto, L.P.; Rosmini, M.R.; Signorini, M.L. Probiotics and broiler growth performance: A meta-analysis of randomised controlled trials. Br. Poult. Sci. 2014, 55, 483–494. [Google Scholar] [CrossRef]
- Liu, D.; Van Belleghem, J.D.; de Vries, C.R.; Burgener, E.; Chen, Q.; Manasherob, R.; Aronson, J.R.; Amanatullah, D.F.; Tamma, P.D.; Suh, G.A. The safety and toxicity of phage therapy: A review of animal and clinical studies. Viruses 2021, 13, 1268. [Google Scholar] [CrossRef]
- Steele, A.; Stacey, H.J.; de Soir, S.; Jones, J.D. The Safety and Efficacy of Phage Therapy for Superficial Bacterial Infections: A Systematic Review. Antibiotics 2020, 9, 754. [Google Scholar] [CrossRef]
- Mohd Shaufi, M.A.; Sieo, C.C.; Chong, C.W.; Gan, H.M.; Ho, Y.W. Deciphering chicken gut microbial dynamics based on high-throughput 16S rRNA metagenomics analyses. Gut Pathog. 2015, 7, 4. [Google Scholar] [CrossRef]
- Shaufi, M.A.M.; Sieo, C.C.; Cheah, Y.-K.; Chong, C.W.; Omar, A.R.; Ho, Y.W.; Tan, G.H. Discrimination of Escherichia coli isolates recovered from mucosal contents of chicken intestines and different age by repetitive elements sequence-based PCR. J. Biochem. Microbiol. Biotechnol. 2017, 5, 7–12. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Poultry; The National Academies Press: Washington, DC, USA, 1994; p. 176. [Google Scholar]
- Naidoo, V.; McGaw, L.J.; Bisschop, S.P.; Duncan, N.; Eloff, J.N. The value of plant extracts with antioxidant activity in attenuating coccidiosis in broiler chickens. Vet. Parasitol. 2008, 153, 214–219. [Google Scholar] [CrossRef]
- Wang, Y.-B.; Xu, B.-H. Effect of different selenium source (sodium selenite and selenium yeast) on broiler chickens. Anim. Feed Sci. Technol. 2008, 144, 306–314. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glockner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Paulson, J.N. metagenomeSeq: Statistical analysis for sparse high-throughput sequencing. Bioconductor Package 2014, 1, 191. [Google Scholar]
- Clarke, K.; Gorley, R. PRIMER-6. 6.1. 2007. Available online: https://www.bioconductor.org/packages/devel/bioc/vignettes/metagenomeSeq/inst/doc/metagenomeSeq.pdf (accessed on 13 March 2023).
- Langille, M.G.I.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Tyson, G.W.; Hugenholtz, P.; Beiko, R.G. STAMP: Statistical analysis of taxonomic and functional profiles. Bioinformatics 2014, 30, 3123–3124. [Google Scholar] [CrossRef]
- Duncan, D.B. Multiple range and multiple F tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Upadhaya, S.D.; Ahn, J.M.; Cho, J.H.; Kim, J.Y.; Kang, D.K.; Kim, S.W.; Kim, H.B.; Kim, I.H. Bacteriophage cocktail supplementation improves growth performance, gut microbiome and production traits in broiler chickens. J. Anim. Sci. Biotechnol. 2021, 12, 49. [Google Scholar] [CrossRef]
- Lu, J.; Idris, U.; Harmon, B.; Hofacre, C.; Maurer, J.J.; Lee, M.D. Diversity and succession of the intestinal bacterial community of the maturing broiler chicken. Appl. Environ. Microbiol. 2003, 69, 6816–6824. [Google Scholar] [CrossRef] [PubMed]
- Torok, V.A.; Ophel-Keller, K.; Loo, M.; Hughes, R.J. Application of methods for identifying broiler chicken gut bacterial species linked with increased energy metabolism. Appl. Environ. Microbiol. 2008, 74, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Forster, R.J.; Yu, H.; Chambers, J.R.; Wheatcroft, R.; Sabour, P.M.; Chen, S. Molecular analysis of bacterial populations in the ileum of broiler chickens and comparison with bacteria in the cecum. FEMS Microbiol. Ecol. 2002, 41, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.; Sodhi, N.; Chenu, J.; Cox, J.; Riordan, S.; Mitchell, H. The interplay between Campylobacter and Helicobacter species and other gastrointestinal microbiota of commercial broiler chickens. Gut Pathog. 2014, 6, 18. [Google Scholar] [CrossRef]
- Eeckhaut, V.; Van Immerseel, F.; Croubels, S.; De Baere, S.; Haesebrouck, F.; Ducatelle, R.; Louis, P.; Vandamme, P. Butyrate production in phylogenetically diverse Firmicutes isolated from the chicken caecum. Microb. Biotechnol. 2011, 4, 503–512. [Google Scholar] [CrossRef]
- Polansky, O.; Sekelova, Z.; Faldynova, M.; Sebkova, A.; Sisak, F.; Rychlik, I. Important Metabolic Pathways and Biological Processes Expressed by Chicken Cecal Microbiota. Appl. Environ. Microbiol. 2016, 82, 1569–1576. [Google Scholar] [CrossRef]
- Oakley, B.B.; Lillehoj, H.S.; Kogut, M.H.; Kim, W.K.; Maurer, J.J.; Pedroso, A.; Lee, M.D.; Collett, S.R.; Johnson, T.J.; Cox, N.A. The chicken gastrointestinal microbiome. FEMS Microbiol. Lett. 2014, 360, 100–112. [Google Scholar] [CrossRef]
- Scheppach, W.; Weiler, F. The butyrate story: Old wine in new bottles? Curr. Opin. Clin. Nutr. Metab. Care 2004, 7, 563–567. [Google Scholar] [CrossRef]
- Sergeant, M.J.; Constantinidou, C.; Cogan, T.A.; Bedford, M.R.; Penn, C.W.; Pallen, M.J. Extensive microbial and functional diversity within the chicken cecal microbiome. PLoS ONE 2014, 9, e91941. [Google Scholar] [CrossRef]
- Hu, Z.; Guo, Y. Effects of dietary sodium butyrate supplementation on the intestinal morphological structure, absorptive function and gut flora in chickens. Anim. Feed Sci. Technol. 2007, 132, 240–249. [Google Scholar] [CrossRef]
- Timbermont, L.; Lanckriet, A.; Dewulf, J.; Nollet, N.; Schwarzer, K.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Control of Clostridium perfringens-induced necrotic enteritis in broilers by target-released butyric acid, fatty acids and essential oils. Avian Pathol. 2010, 39, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Bjerrum, L.; Engberg, R.M.; Leser, T.D.; Jensen, B.B.; Finster, K.; Pedersen, K. Microbial community composition of the ileum and cecum of broiler chickens as revealed by molecular and culture-based techniques. Poult. Sci. 2006, 85, 1151–1164. [Google Scholar] [CrossRef]
- Waite, D.W.; Taylor, M.W. Characterizing the avian gut microbiota: Membership, driving influences, and potential function. Front. Microbiol. 2014, 5, 223. [Google Scholar] [CrossRef]
- Ercolini, D. High-throughput sequencing and metagenomics: Moving forward in the culture-independent analysis of food microbial ecology. Appl. Environ. Microbiol. 2013, 79, 3148–3155. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Basal Diet (C) | BD + 1 g/kg Phage Cocktail (1ϕ) | BD + 2 g/kg Phage Cocktail (2ϕ) | BD + 1 g/kg Probiotic (P) | BD + 1 g/kg Phage Cocktail + 1 g/kg Probiotic (1ϕP) | BD + 2 g/kg Phage Cocktail + 1 g/kg Probiotic (2ϕP) | |
|---|---|---|---|---|---|---|
| Ingredient (g/kg) | ||||||
| Corn | 538.60 | 537.60 | 536.60 | 537.60 | 536.60 | 535.60 |
| Soybean meal, 48% Cp | 361.90 | 361.90 | 361.90 | 361.90 | 361.90 | 361.90 |
| Fish meal | 30.00 | 30.00 | 30.00 | 30.00 | 30.00 | 30.00 |
| Palm oil | 37.40 | 37.40 | 37.40 | 37.40 | 37.40 | 37.40 |
| 60% choline chloride | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 |
| Vitamin premix † | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 |
| Mineral premix ‡ | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Salt (NaCl) | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| DL-Methionine | 1.80 | 1.80 | 1.80 | 1.80 | 1.80 | 1.80 |
| Limestone | 13.00 | 13.00 | 13.00 | 13.00 | 13.00 | 13.00 |
| Dicalcium phosphate | 11.50 | 11.50 | 11.50 | 11.50 | 11.50 | 11.50 |
| Phage cocktail | 0.00 | 1.00 | 2.00 | 0.00 | 1.00 | 2.00 |
| Probiotic | 0.00 | 0.00 | 0.00 | 1.00 | 1.00 | 1.00 |
| Total | 1000.00 | 1000.00 | 1000.00 | 1000.00 | 1000.00 | 1000.00 |
| Calculated analysis | ||||||
| Metabolisable energy (MJ/kg) | 13.06 | 13.06 | 13.06 | 13.06 | 13.06 | 13.06 |
| Crude protein | 220.00 | 220.00 | 220.00 | 220.00 | 220.00 | 220.00 |
| Crude fat | 63.10 | 63.10 | 63.10 | 63.10 | 63.10 | 63.10 |
| Crude fibre | 38.00 | 38.00 | 38.00 | 38.00 | 38.00 | 38.00 |
| Digestible methionine + cysteine | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 | 9.50 |
| Digestible lysine | 13.70 | 13.70 | 13.70 | 13.70 | 13.70 | 13.70 |
| Calcium | 10.20 | 10.20 | 10.20 | 10.20 | 10.20 | 10.20 |
| Available phosphorus | 4.50 | 4.50 | 4.50 | 4.50 | 4.50 | 4.50 |
| Chemical analysis | ||||||
| Crude protein | 217.70 | 217.70 | 217.70 | 217.70 | 217.70 | 217.70 |
| Crude fat | 54.80 | 54.80 | 54.80 | 54.80 | 54.80 | 54.80 |
| Crude fibre | 38.80 | 38.80 | 38.80 | 39.01 | 39.02 | 39.02 |
| Calcium | 6.50 | 6.55 | 6.61 | 6.66 | 6.71 | 6.77 |
| Phosphorus | 5.16 | 5.16 | 5.16 | 5.16 | 5.16 | 5.16 |
| Sodium | 6.70 | 6.71 | 6.72 | 6.70 | 6.71 | 6.72 |
| Basal Diet (C) | BD + 1 g/kg Phage Cocktail (1ϕ) | BD + 2 g/kg Phage Cocktail (2ϕ) | BD + 1 g/kg Probiotic (P) | BD + 1 g/kg Phage Cocktail + 1 g/kg Probiotic (1ϕP) | BD + 2 g/kg Phage Cocktail + 1 g/kg Probiotic (2ϕP) | |
|---|---|---|---|---|---|---|
| Ingredient (g/kg) | ||||||
| Corn | 602.70 | 601.70 | 600.70 | 601.70 | 600.70 | 599.70 |
| Soybean meal, 48% Cp | 318.60 | 318.60 | 318.60 | 318.60 | 318.60 | 318.60 |
| Fish meal | 30.00 | 30.00 | 30.00 | 30.00 | 30.00 | 30.00 |
| Palm Oil | 24.50 | 24.50 | 24.50 | 24.50 | 24.50 | 24.50 |
| 60% choline chloride | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 | 2.00 |
| Vitamin premix † | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 |
| Mineral premix ‡ | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Salt (NaCl) | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| DL-Methionine | 0.40 | 0.40 | 0.40 | 0.40 | 0.40 | 0.40 |
| Limestone | 13.00 | 13.00 | 13.00 | 13.00 | 13.00 | 13.00 |
| Dicalcium phosphate | 6.50 | 6.50 | 6.50 | 6.50 | 6.50 | 6.50 |
| Phage cocktail | 0.00 | 1.00 | 2.00 | 0.00 | 1.00 | 2.00 |
| Probiotic | 0.00 | 0.00 | 0.00 | 1.00 | 1.00 | 1.00 |
| Total | 1000.00 | 1000.00 | 1000.00 | 1000.00 | 1000.00 | 1000.00 |
| Calculated analysis | ||||||
| Metabolisable energy (MJ/kg) | 13.06 | 13.06 | 13.06 | 13.06 | 13.06 | 13.06 |
| Crude protein | 199.90 | 199.90 | 199.90 | 199.90 | 199.90 | 199.90 |
| Crude fat | 52.20 | 52.20 | 52.20 | 52.20 | 52.20 | 52.20 |
| Crude fibre | 36.50 | 36.50 | 36.50 | 36.50 | 36.50 | 36.50 |
| Digestible methionine + cysteine | 8.50 | 8.50 | 8.50 | 8.50 | 8.50 | 8.50 |
| Digestible lysine | 12.00 | 12.00 | 12.00 | 12.00 | 12.00 | 12.00 |
| Calcium | 9.00 | 9.00 | 9.00 | 9.00 | 9.00 | 9.00 |
| Available phosphorus | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 | 3.50 |
| Chemical analysis | ||||||
| Crude protein | 212.05 | 212.50 | 212.50 | 212.50 | 212.50 | 212.50 |
| Crude fat | 45.00 | 45.00 | 45.00 | 45.00 | 45.00 | 45.00 |
| Crude fibre | 40.50 | 40.50 | 40.50 | 40.71 | 40.71 | 40.71 |
| Calcium | 6.80 | 6.85 | 6.91 | 6.96 | 7.01 | 7.07 |
| Phosphorus | 4.50 | 4.50 | 4.50 | 4.50 | 4.50 | 4.50 |
| Sodium | 1.90 | 1.91 | 1.92 | 1.90 | 1.91 | 1.92 |
| Item † | Age (days) | C | 1ϕ | 2ϕ | P | 1ϕP | 2ϕP |
|---|---|---|---|---|---|---|---|
| BW (g) | 1 | 43.00 ± 1.10 | 42.78 ± 1.09 | 42.98 ± 0.78 | 42.82 ± 0.96 | 42.78 ± 1.05 | 43.32 ± 0.38 |
| 21 | 859.58 ± 14.20 | 874.07 ± 8.85 | 864.67 ± 11.05 | 871.78 ± 6.09 | 874.02 ± 12.31 | 855.10 ± 13.74 | |
| 35 | 1605.30 ± 39.45 a | 1734.17 ± 47.55 ab | 1741.27 ± 56.58 ab | 1690.43 ± 40.05 ab | 1785.75 ± 31.93 b | 1733.63 ± 56.00 ab | |
| BWG (g/bird) | 1–21 | 816.63 ± 13.97 | 861.32 ± 9.47 | 821.70 ± 11.09 | 828.98 ± 6.04 | 831.25 ± 11.91 | 811.80 ± 12.95 |
| 22–35 | 745.72 ± 40.37 a | 860.10 ± 50.69 ab | 876.60 ± 46.20 ab | 818.65 ± 41.87 ab | 911.73 ± 32.82 b | 878.53 ± 47.67 ab | |
| 1–35 | 1562.35 ± 40.42 a | 1691.42 ± 47.67 ab | 1698.30 ± 56.55 ab | 1647.63 ± 39.82 ab | 1742.98 ± 32.76 b | 1690.33 ± 55.75 ab | |
| FI (g/bird) | 1–21 | 1169.88 ± 18.03 | 1153.10 ± 10.46 | 1111.35 ± 30.52 | 1104.87 ± 37.66 | 1160.58 ± 20.14 | 1131.58 ± 10.33 |
| 22–35 | 1748.38 ± 24.66 | 1761.45 ± 51.10 | 1650.32 ± 104.62 | 1653.68 ± 40.06 | 1740.27 ± 63.75 | 1760.85 ± 44.20 | |
| 1–35 | 2918.27 ± 38.40 | 2914.55 ± 55.02 | 2922.88 ± 158.00 | 2758.53 ± 51.98 | 2900.85 ± 70.40 | 2892.42 ± 53.92 | |
| FCR (feed/gain) | 1–21 | 1.43 ± 0.17 c | 1.35 ± 0.17 a | 1.37 ± 0.15 ab | 1.40 ± 0.10 bc | 1.37 ± 0.11 ab | 1.34 ± 0.15 a |
| 22–35 | 2.53 ± 0.12 b | 2.10 ± 0.08 a | 2.06 ± 0.06 a | 1.97 ± 0.06 a | 1.89 ± 0.04 a | 2.01 ± 0.07 a | |
| 1–35 | 1.87 ± 0.04 b | 1.68 ± 0.03 a | 1.65 ± 0.03 a | 1.61 ± 0.03 a | 1.60 ± 0.03 a | 1.61 ± 0.03 a | |
| Mortality rate (%) | 1–21 | 2.08 | 2.08 | 2.08 | NIL | NIL | 2.08 |
| 22–35 | 2.08 | NIL | 2.08 | NIL | 2.08 | NIL | |
| 1–35 | 4.17 | 2.08 | 4.17 | NIL | 2.08 | 2.08 |
| a. Marginal Test | |||||
|---|---|---|---|---|---|
| Source | Df | SS | MS | Pseudo-F | pMC |
| Treatment | 5 | 28,910 | 5782.1 | 4.0189 | 0.001 |
| Residual | 29 | 41,723 | 1438.7 | ||
| Total | 34 | 70,634 | |||
| b. Pairwise Test | |||||
| Groups † | t | Unique Perms | pMC | ||
| 1ϕ, 1ϕP | 2.2179 | 403 | 0.011 | ||
| 1ϕ, 2ϕ | 0.99346 | 405 | 0.419 | ||
| 1ϕ, 2ϕP | 2.7819 | 405 | 0.002 | ||
| 1ϕ, C | 0.97549 | 408 | 0.437 | ||
| 1ϕ, P | 0.78783 | 413 | 0.672 | ||
| 1ϕP, 2ϕ | 2.6116 | 401 | 0.002 | ||
| 1ϕP,2ϕP | 1.179 | 418 | 0.234 | ||
| 1ϕP, C | 2.4102 | 403 | 0.006 | ||
| 1ϕP, P | 2.2141 | 410 | 0.005 | ||
| 2ϕ, 2ϕP | 3.2675 | 401 | 0.002 | ||
| 2ϕ, C | 0.80525 | 408 | 0.651 | ||
| 2ϕ, P | 0.98786 | 397 | 0.393 | ||
| 2ϕP, C | 3.167 | 399 | 0.001 | ||
| 2ϕP, P | 2.7201 | 400 | 0.002 | ||
| C, P | 1.0209 | 394 | 0.3850 | ||
| OTUs | Taxonomy | LogFC | Standard Error (SE) | p-Values | Adjusted p-Values |
|---|---|---|---|---|---|
| Otu000006 | Bacteroides uniformis | 3.238460356 | 0.513486 | 2.85 ×10−10 | 1.64 ×10−8 |
| Otu000001 | Bacteroides | 3.082429046 | 0.511617 | 1.69 ×10−9 | 6.49 ×10−8 |
| Otu000585 | Odoribacter | 2.884646375 | 1.034084 | 0.005278 | 0.02529 |
| Otu000009 | Alistipes | 2.764961394 | 0.432556 | 1.64 ×10−10 | 1.64 ×10−8 |
| Otu000139 | Alistipes finegoldii | 2.723944494 | 0.680663 | 6.28 ×10−5 | 0.000723 |
| Otu000269 | Ruminococcaceae_unclassified | 2.720698332 | 1.02679 | 0.008056 | 0.034313 |
| Otu000070 | Alistipes | 2.647180277 | 0.526343 | 4.92 ×10−7 | 1.41 ×10−5 |
| Otu000624 | Ruminococcaceae UCG-014 | 2.618442967 | 1.056871 | 0.013229 | 0.04612 |
| Otu000111 | Anaerotruncus | 2.539148885 | 0.862369 | 0.003236 | 0.018607 |
| Otu000014 | Ruminococcus | 2.495897314 | 0.525048 | 2.00 ×10−6 | 4.59 ×10−5 |
| Otu000031 | Lachnospiraceae_unclassified | 2.470502349 | 0.558793 | 9.82 ×10−6 | 0.000188 |
| Otu000117 | Ruminococcaceae UCG-005 | 2.445622307 | 0.891623 | 0.00609 | 0.028015 |
| Otu000048 | Bacillaceae_unclassified | 2.308772376 | 0.862055 | 0.007401 | 0.032737 |
| Otu000170 | Bacteroides | 2.245170744 | 0.57669 | 9.89 ×10−5 | 0.001034 |
| Otu000319 | Rhodospirillaceae | 2.162299849 | 0.88805 | 0.014897 | 0.047586 |
| Otu000005 | Alistipes onderdonkii | 2.121163741 | 0.521993 | 4.83 ×10−5 | 0.000695 |
| Otu000210 | Anaerotruncus | 2.025946607 | 0.945538 | 0.032142 | 0.080355 |
| Otu000015 | Clostridium X1Vb | 2.019893556 | 0.617525 | 0.001072 | 0.007704 |
| Otu000337 | Desulfovibrio | 2.014867537 | 0.841755 | 0.016682 | 0.049954 |
| Otu000675 | Anaerostipes | 2.013275931 | 1.087486 | 0.064125 | 0.122906 |
| Otu000032 | Clostridium X1Va | 1.982675782 | 0.536143 | 0.000217 | 0.002082 |
| Otu000643 | Vampirovibrio | 1.967914419 | 1.072608 | 0.06655 | 0.125463 |
| Otu000353 | Alistipes putredinis | 1.899173634 | 0.770005 | 0.013646 | 0.046157 |
| Otu000204 | Ruminococcaceae UCG-014 | 1.859164848 | 0.750237 | 0.013208 | 0.04612 |
| Otu000157 | Ruminococcaceae_unclassified | 1.841791976 | 0.706923 | 0.009178 | 0.036394 |
| Otu000866 | Clostridium | 1.840566564 | 0.885046 | 0.03756 | 0.086387 |
| Otu000367 | Coprobacillus | 1.822128533 | 1.039682 | 0.079674 | 0.143164 |
| Otu000694 | Lachnospiraceae_unclassified | 1.815009851 | 1.076386 | 0.091756 | 0.158407 |
| Otu000066 | Bacteroides fragilis | 1.803001647 | 0.592086 | 0.002325 | 0.014857 |
| Otu000025 | Lachnospiraceae_unclassified | 1.784599464 | 0.409758 | 1.33 ×10−5 | 0.000218 |
| Otu000426 | Ruminococcaceae UCG-014 | 1.782185523 | 0.793533 | 0.024711 | 0.063151 |
| Otu000027 | Eisenbergiella | 1.766953157 | 0.495082 | 0.000358 | 0.00317 |
| Otu000067 | Butyricimonas | 1.753087241 | 0.611499 | 0.004146 | 0.02165 |
| Otu000075 | Lachnospiraceae_unclassified | 1.747367694 | 0.612453 | 0.00433 | 0.02165 |
| Otu000247 | Bacteria_unclassified | 1.741509274 | 0.770742 | 0.023851 | 0.062338 |
| Otu000022 | Faecalibacterium prausnitzii | 1.738699859 | 0.433282 | 6.00 ×10−5 | 0.000723 |
| Otu000104 | Lachnospiraceae_unclassified | 1.715040783 | 0.493786 | 0.000514 | 0.004224 |
| Otu002794 | Lachnospiraceae_unclassified | 1.702047151 | 1.080272 | 0.115124 | 0.194695 |
| Otu000095 | Oscillospira | 1.686705907 | 0.725797 | 0.020129 | 0.055115 |
| Otu000338 | Ruminococcaceae UCG-014 | 1.677289282 | 0.969175 | 0.083517 | 0.14776 |
| Otu000186 | Anaerotruncus | 1.668325958 | 0.655373 | 0.010909 | 0.041816 |
| Otu000148 | Oscillospira | 1.665393273 | 0.699204 | 0.017226 | 0.049954 |
| Otu000947 | Clostridiales | 1.659154189 | 1.306282 | 0.204037 | 0.312856 |
| Otu000802 | Clostridium IV | 1.654944242 | 0.813506 | 0.041918 | 0.090954 |
| Otu000041 | Clostridium X1Va | 1.644739429 | 0.517848 | 0.001493 | 0.010098 |
| Otu000069 | Ruminococcus | 1.601694881 | 0.542143 | 0.003133 | 0.018607 |
| Otu000044 | Lachnospiraceae_unclassified | 1.575177456 | 0.551342 | 0.004277 | 0.02165 |
| Otu000573 | Ruminococcus | 1.546199668 | 1.1055 | 0.16192 | 0.258623 |
| Otu000057 | Ruminococcus | 1.517227727 | 0.458007 | 0.000924 | 0.007085 |
| Otu000007 | Phascolarctobacterium | 1.505713845 | 0.607781 | 0.013234 | 0.04612 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaufi, M.A.M.; Sieo, C.C.; Chong, C.W.; Geok Hun, T.; Omar, A.R.; Han Ming, G.; Wan Ho, Y. Effects of Phage Cocktail, Probiotics, and Their Combination on Growth Performance and Gut Microbiota of Broiler Chickens. Animals 2023, 13, 1328. https://doi.org/10.3390/ani13081328
Shaufi MAM, Sieo CC, Chong CW, Geok Hun T, Omar AR, Han Ming G, Wan Ho Y. Effects of Phage Cocktail, Probiotics, and Their Combination on Growth Performance and Gut Microbiota of Broiler Chickens. Animals. 2023; 13(8):1328. https://doi.org/10.3390/ani13081328
Chicago/Turabian StyleShaufi, Mohd Asrore Mohd, Chin Chin Sieo, Chun Wie Chong, Tan Geok Hun, Abdul Rahman Omar, Gan Han Ming, and Yin Wan Ho. 2023. "Effects of Phage Cocktail, Probiotics, and Their Combination on Growth Performance and Gut Microbiota of Broiler Chickens" Animals 13, no. 8: 1328. https://doi.org/10.3390/ani13081328
APA StyleShaufi, M. A. M., Sieo, C. C., Chong, C. W., Geok Hun, T., Omar, A. R., Han Ming, G., & Wan Ho, Y. (2023). Effects of Phage Cocktail, Probiotics, and Their Combination on Growth Performance and Gut Microbiota of Broiler Chickens. Animals, 13(8), 1328. https://doi.org/10.3390/ani13081328