
Effects of Natural and Synthetic Astaxanthin on Growth, Body Color, and Transcriptome and Metabolome Profiles in the Leopard Coralgrouper (Plectropomus leopardus)


 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Experimental Diets
2.3. Experimental Fish and Sample Collection
2.4. Colorimetric Evaluation
2.5. RNA Extraction and Transcriptome Sequencing Analysis
2.6. Metabolome Analysis
2.7. Quantitative Real-Time PCR (qPCR)
2.8. Data Calculation and Statistical Analysis
3. Results
3.1. Growth
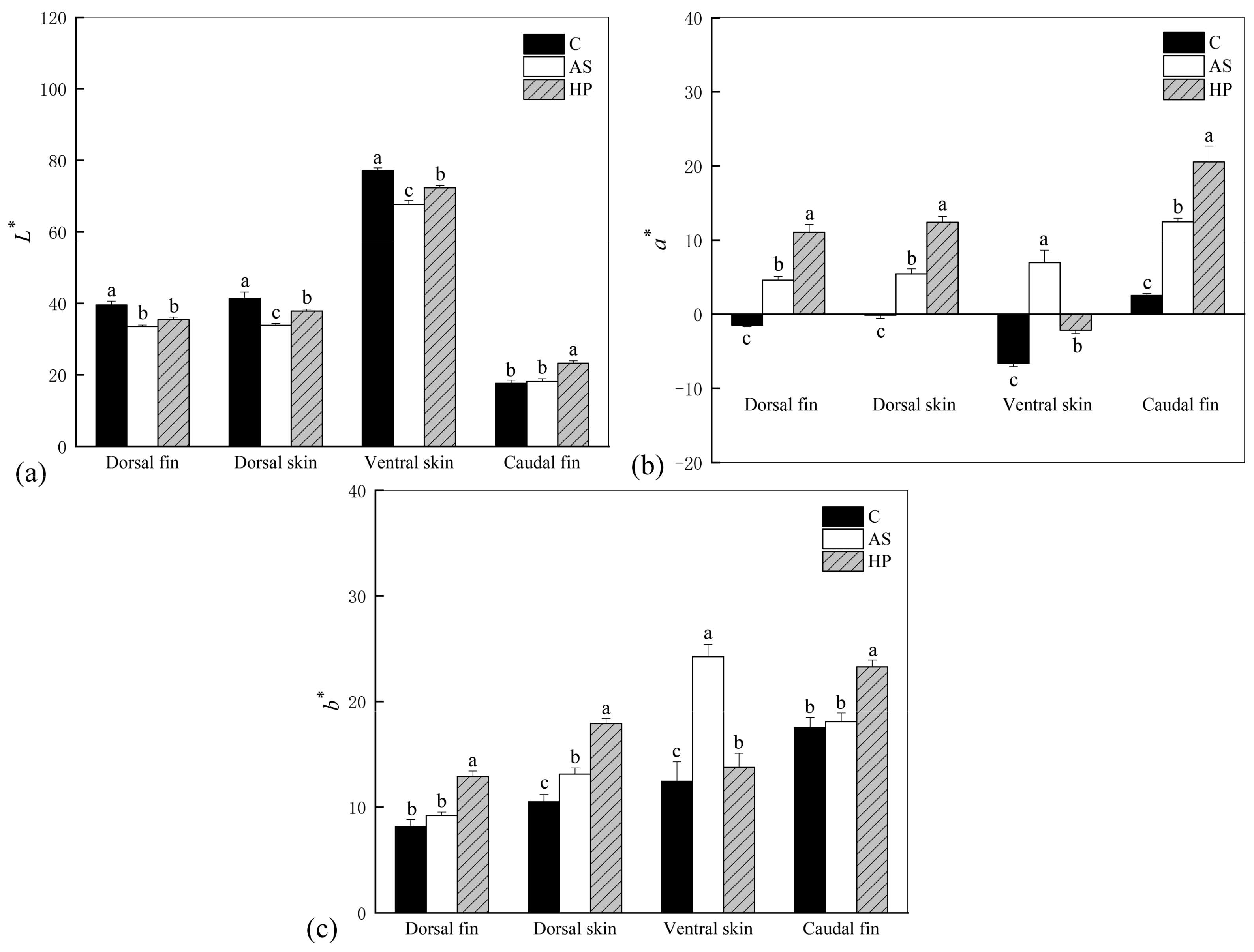
3.2. Colorimetric Evaluation
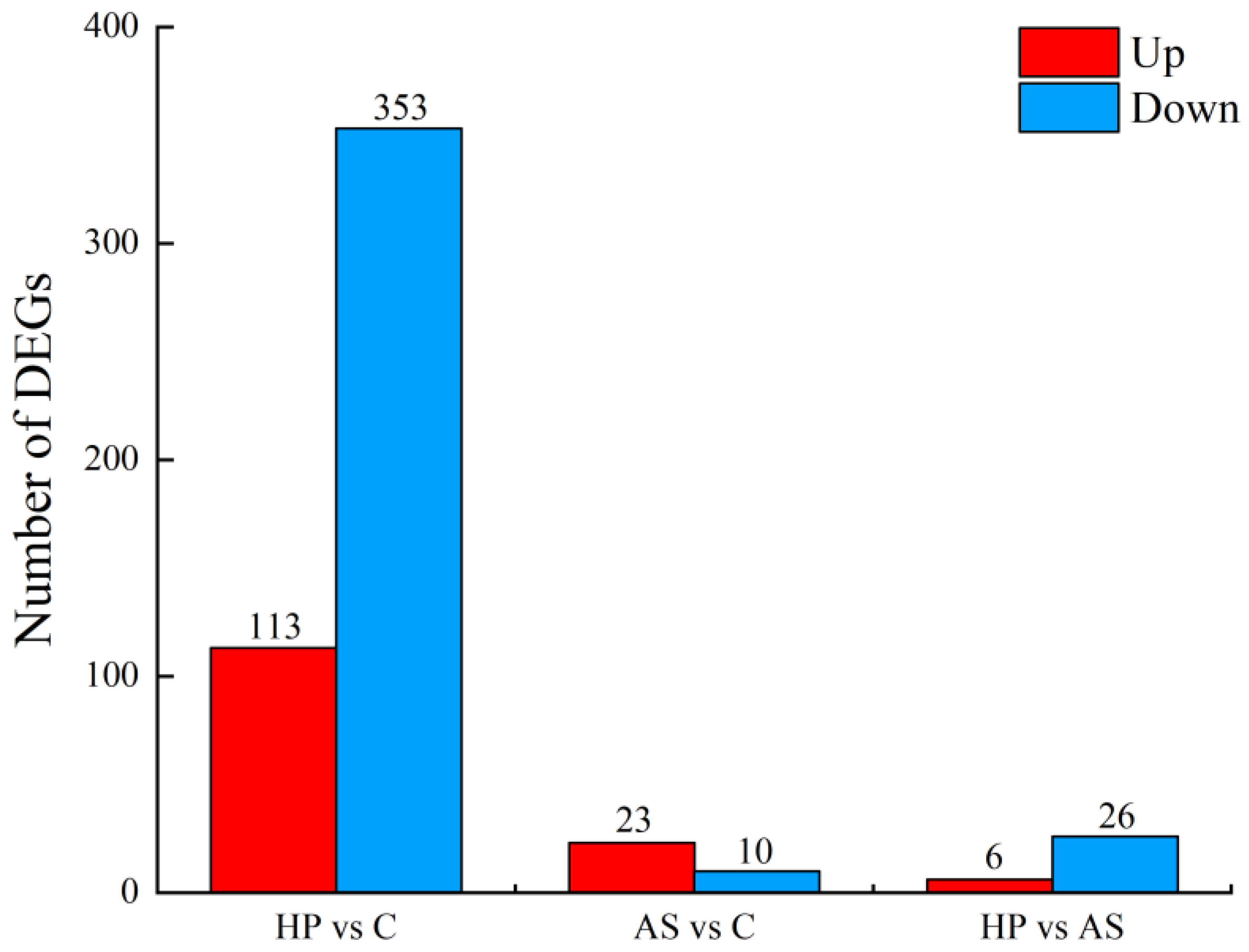
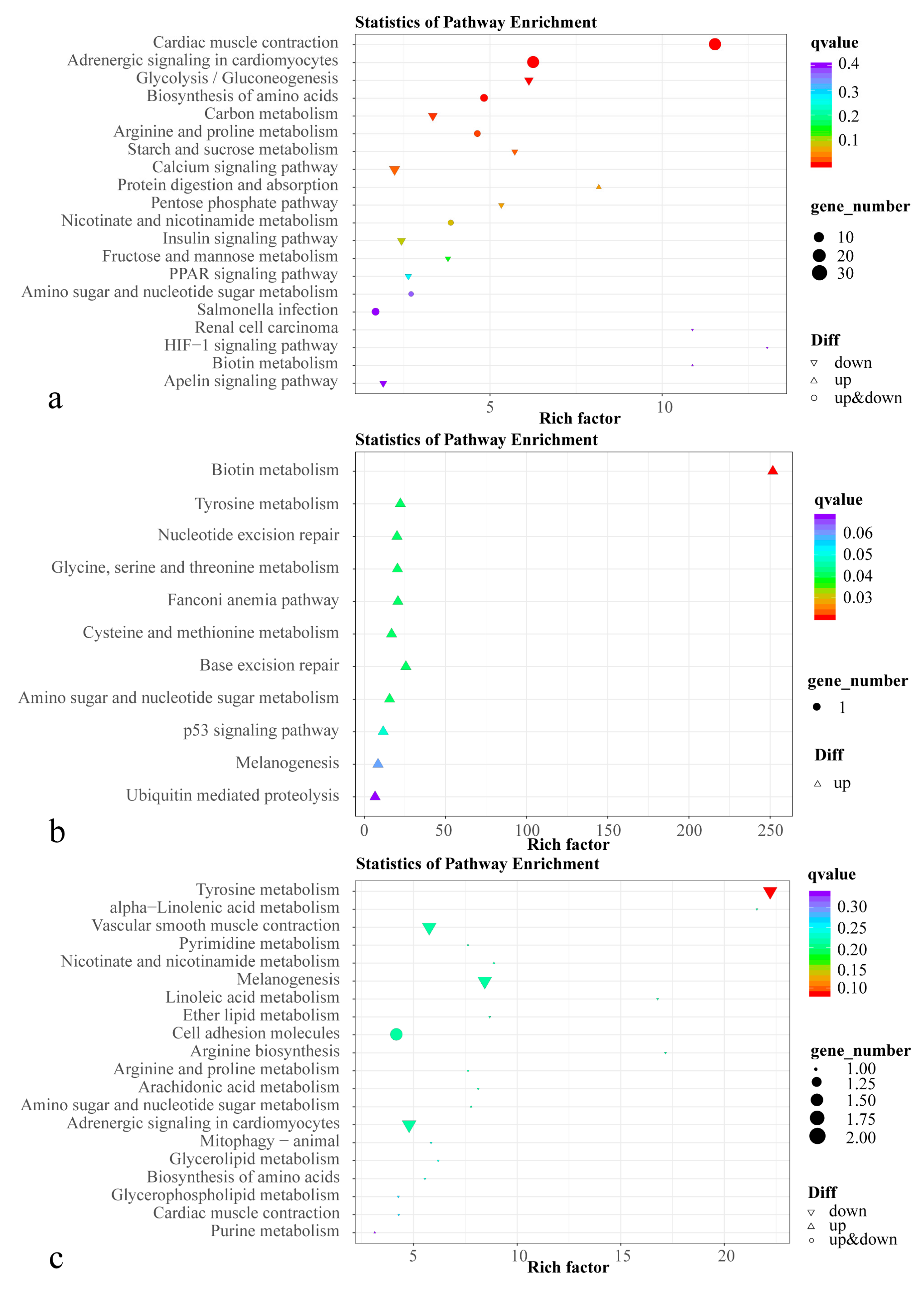
3.3. Transcriptome Analysis
3.4. Metabolome Analysis
3.5. Integrated Transcriptome and Metabolome Analysis
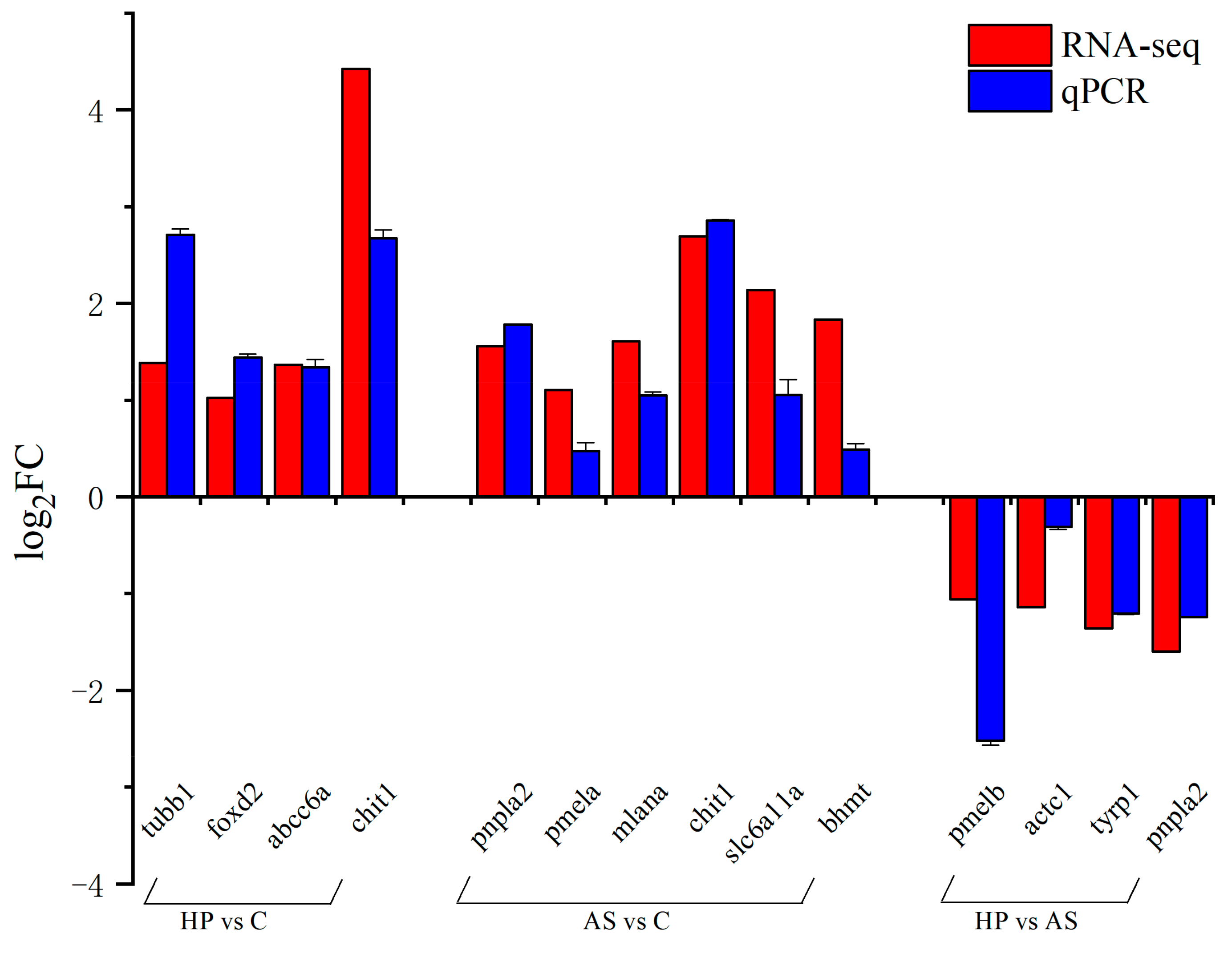
3.6. Validation of Gene Expression Levels by qPCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Protas, M.E.; Patel, N.H. Evolution of coloration patterns. Annu. Rev. Cell Dev. Biol. 2008, 24, 425–446. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, G.M.; Kelley, J.L.; Morrell, L.J. Colour change and assortment in the western rainbowfish. Anim. Behav. 2010, 79, 1025–1030. [Google Scholar] [CrossRef]
- Irion, U.; Nüsslein-Volhard, C. The identification of genes involved in the evolution of color patterns in fish. Curr. Opin. Genet. Dev. 2019, 57, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Parichy, D.M. Pigment patterns: Fish in stripes and spots. Curr. Biol. 2003, 13, R947–R950. [Google Scholar] [CrossRef] [PubMed]
- Henning, F.; Jones, J.C.; Franchini, P.; Meyer, A. Transcriptomics of morphological color change in polychromatic Midas cichlids. BMC Genom. 2013, 14, 171. [Google Scholar] [CrossRef]
- Goda, M.; Fujiyoshi, Y.; Sugimoto, M.; Fujii, R. Novel Dichromatic Chromatophores in the Integument of the Mandarin Fish Synchiropus splendidus. Biol. Bull. 2013, 224, 14–17. [Google Scholar] [CrossRef]
- Singh, A.P.; Nüsslein-Volhard, C. Zebrafish Stripes as a Model for Vertebrate Colour Pattern Formation. Curr. Biol. 2015, 25, R81–R92. [Google Scholar] [CrossRef]
- Hirata, M.; Nakamura, K.; Kanemaru, T.; Shibata, Y.; Kondo, S. Pigment cell organization in the hypodermis of zebrafish. Dev. Dyn. 2003, 227, 497–503. [Google Scholar] [CrossRef]
- Nüsslein-Volhard, C.; Singh, A.P. How fish color their skin: A paradigm for development and evolution of adult patterns. BioEssays 2017, 39, 1600231. [Google Scholar] [CrossRef]
- Braasch, I.; Schartl, M.; Volff, J. Evolution of pigment synthesis pathways by gene and genome duplication in fish. BMC Evol. Biol. 2007, 7, 74. [Google Scholar] [CrossRef]
- Bar, I.; Kaddar, E.; Velan, A.; David, L. Melanocortin receptor 1 and black pigmentation in the Japanese ornamental carp (Cyprinus carpio var. Koi). Front. Genet. 2013, 4, 6. [Google Scholar] [CrossRef]
- Borel, P.; Lietz, G.; Goncalves, A.; Szabo De Edelenyi, F.; Lecompte, S.; Curtis, P.; Goumidi, L.; Caslake, M.J.; Miles, E.A.; Packard, C.; et al. CD36 and SR-BI Are Involved in Cellular Uptake of Provitamin A Carotenoids by Caco-2 and HEK Cells, and Some of Their Genetic Variants Are Associated with Plasma Concentrations of These Micronutrients in Humans. J. Nutr. 2013, 143, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Chen, H.; Mandal, B.K.; Wang, J.; Shi, Z.; Lu, G.; Wang, C. HDL receptor/Scavenger receptor B1-Scarb1 and Scarb1-like mediate the carotenoid-based red coloration in fish. Aquaculture 2021, 545, 737208. [Google Scholar] [CrossRef]
- Ahi, E.P.; Lecaudey, L.A.; Ziegelbecker, A.; Steiner, O.; Goessler, W.; Sefc, K.M. Expression levels of the tetratricopeptide repeat protein gene ttc39b covary with carotenoid-based skin colour in cichlid fish. Biol. Lett. 2020, 16, 20200629. [Google Scholar] [CrossRef]
- Tian, X.; Pang, X.; Wang, L.; Li, M.; Dong, C.; Ma, X.; Wang, L.; Song, D.; Feng, J.; Xu, P.; et al. Dynamic regulation of mRNA and miRNA associated with the developmental stages of skin pigmentation in Japanese ornamental carp. Gene 2018, 666, 32–43. [Google Scholar] [CrossRef]
- Parichy, D.M. Evolution of pigment cells and patterns: Recent insights from teleost fishes. Curr. Opin. Genet. Dev. 2021, 69, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, J.; Li, Y.; Zhao, L.; Liu, Z. Analysis of yellow mutant rainbow trout transcriptomes at different developmental stages reveals dynamic regulation of skin pigmentation genes. Sci. Rep. 2022, 12, 256. [Google Scholar] [CrossRef]
- Lobo, G.P.; Amengual, J.; Palczewski, G.; Babino, D.; von Lintig, J. Mammalian Carotenoid-oxygenases: Key players for carotenoid function and homeostasis. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2012, 1821, 78–87. [Google Scholar] [CrossRef]
- Lopes, R.J.; Johnson, J.D.; Toomey, M.B.; Ferreira, M.S.; Araujo, P.M.; Melo-Ferreira, J.; Andersson, L.; Hill, G.E.; Corbo, J.C.; Carneiro, M. Genetic Basis for Red Coloration in Birds. Curr. Biol. 2016, 26, 1427–1434. [Google Scholar] [CrossRef]
- Twyman, H.; Valenzuela, N.; Literman, R.; Andersson, S.; Mundy, N.I. Seeing red to being red: Conserved genetic mechanism for red cone oil droplets and co-option for red coloration in birds and turtles. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161208. [Google Scholar] [CrossRef]
- Huang, D.; Lewis, V.M.; Foster, T.N.; Toomey, M.B.; Corbo, J.C.; Parichy, D.M.; Whitfield, T.T.; White, R.M.; Kelsh, R. Development and genetics of red coloration in the zebrafish relative Danio albolineatus. eLife 2021, 10, e70253. [Google Scholar] [CrossRef]
- Rashidian, G.; Rainis, S.; Prokić, M.D.; Faggio, C. Effects of different levels of carotenoids and light sources on swordtail fish (Xiphophorus helleri) growth, survival rate and reproductive parameters. Nat. Prod. Res. 2021, 35, 3675–3686. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, C.T.; Robaina, L.E.; Fernández-Palacios, H.; Schuchardt, D.; Izquierdo, M.S. Effect of different carotenoid sources and their dietary levels on red porgy (Pagrus pagrus) growth and skin colour. Aquaculture 2005, 244, 223–231. [Google Scholar] [CrossRef]
- Díaz Jiménez, L.; Hernández Vergara, M.P.; Pérez Rostro, C.I.; Olvera Novoa, M.Á. The effect of two carotenoid sources, background colour and light spectrum on the body pigmentation of the clownfish Amphiprion ocellaris. Aquac. Res. 2021, 52, 3052–3061. [Google Scholar] [CrossRef]
- Capelli, B.; Bagchi, D.; Cysewski, G.R. Synthetic astaxanthin is significantly inferior to algal-based astaxanthin as an antioxidant and may not be suitable as a human nutraceutical supplement. Nutrafoods 2013, 12, 145–152. [Google Scholar] [CrossRef]
- Zhu, X.; Hao, R.; Zhang, J.; Tian, C.; Hong, Y.; Zhu, C.; Li, G. Dietary astaxanthin improves the antioxidant capacity, immunity and disease resistance of coral trout (Plectropomus leopardus). Fish Shellfish Immunol. 2022, 122, 38–47. [Google Scholar] [CrossRef]
- Page, G.I.; Davies, S.J. Tissue astaxanthin and canthaxanthin distribution in rainbow trout (Oncorhynchus mykiss) and Atlantic salmon (Salmo salar). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2006, 143, 125–132. [Google Scholar] [CrossRef]
- Moretti, V.M.; Mentasti, T.; Bellagamba, F.; Luzzana, U.; Caprino, F.; Turchini, G.M.; Giani, I.; Valfrè, F. Determination of astaxanthin stereoisomers and colour attributes in flesh of rainbow trout (Oncorhynchus mykiss) as a tool to distinguish the dietary pigmentation source. Food Addit. Contam. 2006, 23, 1056–1063. [Google Scholar] [CrossRef]
- Chekanov, K.; Litvinov, D.; Fedorenko, T.; Chivkunova, O.; Lobakova, E. Combined production of astaxanthin and β-carotene in a new strain of the microalga Bracteacoccus aggregatus BM5/15 (IPPAS C-2045) cultivated in photobioreactor. Biology 2021, 10, 643. [Google Scholar] [CrossRef]
- Ambati, R.; Phang, S.; Ravi, S.; Aswathanarayana, R. Astaxanthin: Sources, Extraction, Stability, Biological Activities and Its Commercial Applications—A Review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef]
- Chekanov, K. Diversity and Distribution of Carotenogenic Algae in Europe: A Review. Mar. Drugs 2023, 21, 108. [Google Scholar] [CrossRef] [PubMed]
- Holtin, K.; Kuehnle, M.; Rehbein, J.; Schuler, P.; Nicholson, G.; Albert, K. Determination of astaxanthin and astaxanthin esters in the microalgae Haematococcus pluvialis by LC-(APCI)MS and characterization of predominant carotenoid isomers by NMR spectroscopy. Anal. Bioanal. Chem. 2009, 395, 1613–1622. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Armstrong, D.W.; Chang, C. Rapid baseline separation of enantiomers and a mesoform of all-trans-astaxanthin, 13-cis-astaxanthin, adonirubin, and adonixanthin in standards and commercial supplements. J. Chromatogr. A 2008, 1194, 172–177. [Google Scholar] [CrossRef] [PubMed]
- Choubert, G.; Mendes-Pinto, M.M.; Morais, R. Pigmenting efficacy of astaxanthin fed to rainbow trout Oncorhynchus mykiss: Effect of dietary astaxanthin and lipid sources. Aquaculture 2006, 257, 429–436. [Google Scholar] [CrossRef]
- Gouveia, L.; Rema, P.; Pereira, O.; Empis, J. Colouring ornamental fish (Cyprinus carpio and Carassius auratus) with microalgal biomass. Aquac. Nutr. 2003, 9, 123–129. [Google Scholar] [CrossRef]
- Jiang, J.; Nuez-Ortin, W.; Angell, A.; Zeng, C.; de Nys, R.; Vucko, M.J. Enhancing the colouration of the marine ornamental fish Pseudochromis fridmani using natural and synthetic sources of astaxanthin. Algal Res. 2019, 42, 101596. [Google Scholar] [CrossRef]
- Cai, X.; Qu, M.; Ding, S.; Wang, H.; Wang, H.; Hu, L.; Su, Y. Differentiation of coral trout (Plectropomus leopardus) based on an analysis of morphology and complete mitochondrial DNA: Are cryptic species present? Acta Oceanol. Sin. 2013, 32, 40–46. [Google Scholar] [CrossRef]
- Shimose, T.; Kanaiwa, M. Influence of the body color and size on the market value of wild captured coralgroupers (Serranidae, Plectropomus): Implications for fisheries management. Fish. Res. 2022, 248, 106223. [Google Scholar] [CrossRef]
- Guan, X.; Wu, H.; Ma, J.; Shan, L.; Lin, S. Effects of natural additives on growth and body color of leopard coralgrouper (Plectropomus leopardus). J. Mar. Sci. 2018, 36, 80–91. [Google Scholar] [CrossRef]
- Chen, C.; Wu, L.; Li, Y.; Song, Z.; Liang, Y.; Zhai, J.; Wang, L.; Pang, Z. Morphology of the Early Age and the Pigment Occurrence and the Effects of Different Feed Additives on the Body Color of Plectropomus leopardus. Prog. Fish. Sci. 2014, 35, 83–90. [Google Scholar] [CrossRef]
- Leary, S.; Underwood, W.; Anthony, R.; Cartner, S.; Grandin, T.; Greenacre, C.; Gwaltney-Brant, S.; McCrackin, M.A.; Meyer, R. AVMA Guidelines for the Euthanasia of Animals, 2020th ed.; American Veterinary Medical Association: Schaumburg, IL, USA, 2020; Volume 2013, pp. 2020–2021. [Google Scholar]
- Korifi, R.; Le Dréau, Y.; Antinelli, J.; Valls, R.; Dupuy, N. CIEL*a*b* color space predictive models for colorimetry devices—Analysisof perfume quality. Talanta 2013, 104, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.; Dillies, M. SARTools: A DESeq2- and EdgeR-Based R Pipeline for Comprehensive Differential Analysis of RNA-Seq Data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Micah, A.D.; Wen, B.; Wang, Q.; Zhang, Y.; Yusuf, A.; Thierry, N.N.B.; Tokpanou, O.S.; Onimisi, M.M.; Adeyemi, S.O.; Gao, J.; et al. Effect of dietary astaxanthin on growth, body color, biochemical parameters and transcriptome profiling of juvenile blood parrotfish (Vieja melanurus ♀ × Amphilophus citrinellus ♂). Aquacult. Rep. 2022, 24, 101142. [Google Scholar] [CrossRef]
- Christiansen, R.; Torrissen, O.J. Growth and survival of Atlantic salmon, Salmo salar L. fed different dietary levels of astaxanthin. Juveniles. Aquac. Nutr. 1996, 2, 55–62. [Google Scholar] [CrossRef]
- Kalinowski, C.T.; Robaina, L.E.; Izquierdo, M.S. Effect of dietary astaxanthin on the growth performance, lipid composition and post-mortem skin colouration of red porgy Pagrus pagrus. Aquac. Int. 2011, 19, 811–823. [Google Scholar] [CrossRef]
- Li, M.; Wu, W.; Zhou, P.; Xie, F.; Zhou, Q.; Mai, K. Comparison effect of dietary astaxanthin and Haematococcus pluvialis on growth performance, antioxidant status and immune response of large yellow croaker Pseudosciaena crocea. Aquaculture 2014, 434, 227–232. [Google Scholar] [CrossRef]
- Amar, E.C.; Kiron, V.; Satoh, S.; Watanabe, T. Influence of various dietary synthetic carotenoids on bio-defence mechanisms in rainbow trout, Oncorhynchus mykiss (Walbaum). Aquac. Res. 2001, 32, 162–173. [Google Scholar] [CrossRef]
- Baron, M.; Davies, S.; Alexander, L.; Snellgrove, D.; Sloman, K.A. The effect of dietary pigments on the coloration and behaviour of flame-red dwarf gourami, Colisa lalia. Anim. Behav. 2008, 75, 1041–1051. [Google Scholar] [CrossRef]
- Angell, A.; de Nys, R.; Mangott, A.; Vucko, M.J. The effects of concentration and supplementation time of natural and synthetic sources of astaxanthin on the colouration of the prawn Penaeus monodon. Algal Res. 2018, 35, 577–585. [Google Scholar] [CrossRef]
- Zhang, C.; Jin, Y.; Yu, Y.; Xiang, J.; Li, F. Effects of natural astaxanthin from microalgae and chemically synthetic astaxanthin supplementation on two different varieties of the ridgetail white prawn (Exopalaemon carinicauda). Algal Res. 2021, 57, 102347. [Google Scholar] [CrossRef]
- Maoka, T.; Sato, W.; Nagai, H.; Takahashi, T. Carotenoids of Red, Brown, and Black Specimens of Plectropomus leopardus, the Coral Trout (Suziara in Japanese). J. Oleo Sci. 2017, 66, ess16179. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; He, C.; Xing, W.; Ma, Z.; Jiang, N.; Li, W.; Sun, X.; Luo, L. Effects of different carotenoids on pigmentation of blood parrot (Cichlasoma synspilum × Cichlasoma citrinellum). J. Aquac. 2016, 7, 414. [Google Scholar] [CrossRef]
- Walsh, N.; Dale, J.; McGraw, K.J.; Pointer, M.A.; Mundy, N.I. Candidate genes for carotenoid coloration in vertebrates and their expression profiles in the carotenoid-containing plumage and bill of a wild bird. Proc. R. Soc. B Biol. Sci. 2012, 279, 58–66. [Google Scholar] [CrossRef]
- Toomey, M.B.; Lopes, R.J.; Araújo, P.M.; Johnson, J.D.; Gazda, M.A.; Afonso, S.; Mota, P.G.; Koch, R.E.; Hill, G.E.; Corbo, J.C.; et al. High-density lipoprotein receptor SCARB1 is required for carotenoid coloration in birds. Proc. Natl. Acad. Sci. USA 2017, 114, 5219–5224. [Google Scholar] [CrossRef]
- Rodríguez-Bernaldo De Quirós, A.; Costa, H.S. Analysis of carotenoids in vegetable and plasma samples: A review. J. Food Compos. Anal. 2006, 19, 97–111. [Google Scholar] [CrossRef]
- Sakudoh, T.; Kuwazaki, S.; Iizuka, T.; Narukawa, J.; Yamamoto, K.; Uchino, K.; Sezutsu, H.; Banno, Y.; Tsuchida, K. CD36 homolog divergence is responsible for the selectivity of carotenoid species migration to the silk gland of the silkworm Bombyx mori[s]. J. Lipid Res. 2012, 54, 482–495. [Google Scholar] [CrossRef]
- Paul, D.S.; Grevengoed, T.J.; Pascual, F.; Ellis, J.M.; Willis, M.S.; Coleman, R.A. Deficiency of cardiac Acyl-CoA synthetase-1 induces diastolic dysfunction, but pathologic hypertrophy is reversed by rapamycin. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2014, 1841, 880–887. [Google Scholar] [CrossRef]
- Chiu, H.C.; Kovacs, A.; Ford, D.A.; Hsu, F.F.; Garcia, R.; Herrero, P.; Saffitz, J.E.; Schaffer, J.E. A novel mouse model of lipotoxic cardiomyopathy. J. Clin. Investig. 2001, 107, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Richards, M.R.; Harp, J.D.; Ory, D.S.; Schaffer, J.E. Fatty acid transport protein 1 and long-chain acyl coenzyme A synthetase 1 interact in adipocytes. J. Lipid Res. 2006, 47, 665–672. [Google Scholar] [CrossRef]
- Parkes, H.A.; Preston, E.; Wilks, D.; Ballesteros, M.; Carpenter, L.; Wood, L.; Kraegen, E.W.; Furler, S.M.; Cooney, G.J. Overexpression of acyl-CoA synthetase-1 increases lipid deposition in hepatic (HepG2) cells and rodent liver in vivo. Am. J. Physiol.-Endocrinol. Metab. 2006, 291, E737–E744. [Google Scholar] [CrossRef] [PubMed]
- Bruce, C.R.; Hoy, A.J.; Turner, N.; Watt, M.J.; Allen, T.L.; Carpenter, K.; Cooney, G.J.; Febbraio, M.A.; Kraegen, E.W. Overexpression of Carnitine Palmitoyltransferase-1 in Skeletal Muscle Is Sufficient to Enhance Fatty Acid Oxidation and Improve High-Fat Diet–Induced Insulin Resistance. Diabetes 2009, 58, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Storch, J.; Corsico, B. The emerging functions and mechanisms of mammalian fatty acid-binding proteins. Annu. Rev. Nutr. 2008, 28, 73–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, H.; Li, N.; Leng, L.; Wang, Y. Tissue Expression and Association with Fatness Traits of Liver Fatty Acid-Binding Protein Gene in Chicken1. Poult. Sci. 2006, 85, 1890–1895. [Google Scholar] [CrossRef]
- Samulin, J.; Berget, I.; Lien, S.; Sundvold, H. Differential gene expression of fatty acid binding proteins during porcine adipogenesis. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 151, 147–152. [Google Scholar] [CrossRef]
- Gill, J.L.; Bishop, S.C.; McCorquodale, C.; Williams, J.L.; Wiener, P. Identification of polymorphisms in the malic enzyme 1, NADP(+)-dependent, cytosolic and nuclear receptor subfamily 0, group B, member 2 genes and their associations with meat and carcass quality traits in commercial Angus cattle. Anim. Genet. 2012, 43, 88–92. [Google Scholar] [CrossRef]
- Ahi, E.P.; Lecaudey, L.A.; Ziegelbecker, A.; Steiner, O.; Glabonjat, R.; Goessler, W.; Hois, V.; Wagner, C.; Lass, A.; Sefc, K.M. Comparative transcriptomics reveals candidate carotenoid color genes in an East African cichlid fish. BMC Genom. 2020, 21, 54. [Google Scholar] [CrossRef]
- Copeman, L.A.; Parrish, C.C.; Brown, J.A.; Harel, M. Effects of docosahexaenoic, eicosapentaenoic, and arachidonic acids on the early growth, survival, lipid composition and pigmentation of yellowtail flounder (Limanda ferruginea): A live food enrichment experiment. Aquaculture 2002, 210, 285–304. [Google Scholar] [CrossRef]
- Mosadeghi, S.; Furnes, B.; Matsuo, A.Y.O.; Schlenk, D. Expression and characterization of cytochrome P450 2X1 in channel catfish (Ictalurus punctatus). Biochim. Biophys. Acta (BBA)-Gen. Subj. 2007, 1770, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Buhler, D.R.; Wang-Buhler, J. Rainbow trout cytochrome P450s: Purification, molecular aspects, metabolic activity, induction and role in environmental monitoring1. Comp. Biochem. Physiol. Part C Pharmacol. Toxicol. Endocrinol. 1998, 121, 107–137. [Google Scholar] [CrossRef]
- Tan, G.; Zhang, G.; Xu, J.; Kang, C.; Yan, Z.; Lei, M.; Pu, X.; Dong, C. PLA2G10 facilitates the cell-cycle progression of soft tissue leiomyosarcoma cells at least by elevating cyclin E1/CDK2 expression. Biochem. Biophys. Res. Commun. 2020, 527, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Taketomi, Y.; Isogai, Y.; Miki, Y.; Yamamoto, K.; Masuda, S.; Hosono, T.; Arata, S.; Ishikawa, Y.; Ishii, T.; et al. Group III secreted phospholipase A2 regulates epididymal sperm maturation and fertility in mice. J. Clin. Investig. 2010, 120, 1400–1414. [Google Scholar] [CrossRef] [PubMed]
- Samuelsson, B. An Elucidation of the Arachidonic Acid Cascade. Drugs 1987, 33, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Hao, R.; Tian, C.; Zhang, J.; Zhu, C.; Li, G. Integrative Transcriptomics and Metabolomics Analysis of Body Color Formation in the Leopard Coralgrouper (Plectropomus leopardus). Front. Mar. Sci. 2021, 8, 726102. [Google Scholar] [CrossRef]
- Zhang, X.T.; Wei, K.J.; Chen, Y.Y.; Shi, Z.C.; Liu, L.K.; Li, J.; Zhang, G.R.; Ji, W. Molecular cloning and expression analysis of tyr and tyrp1 genes in normal and albino yellow catfish Tachysurus fulvidraco. J. Fish. Biol. 2018, 92, 979–998. [Google Scholar] [CrossRef]
- Lin, X.; Tian, C.; Huang, Y.; Shi, H.; Li, G. Comparative Transcriptome Analysis Identifies Candidate Genes Related to Black-Spotted Pattern Formation in Spotted Scat (Scatophagus argus). Animals 2021, 11, 765. [Google Scholar] [CrossRef]
- Berson, J.F.; Harper, D.C.; Tenza, D.; Raposo, G.; Marks, M.S. Pmel17 initiates premelanosome morphogenesis within multivesicular bodies. Mol. Biol. Cell. 2001, 12, 3451–3464. [Google Scholar] [CrossRef]
- Wang, C.; Xu, J.; Kocher, T.D.; Li, M.; Wang, D. CRISPR Knockouts of pmela and pmelb Engineered a Golden Tilapia by Regulating Relative Pigment Cell Abundance. J. Hered. 2022, 113, 398–413. [Google Scholar] [CrossRef]
- Price, E.R.; Horstmann, M.A.; Wells, A.G.; Weilbaecher, K.N.; Takemoto, C.M.; Landis, M.W.; Fisher, D.E. α-Melanocyte-stimulating Hormone Signaling Regulates Expression of microphthalmia, a Gene Deficient in Waardenburg Syndrome. J. Biol. Chem. 1998, 273, 33042–33047. [Google Scholar] [CrossRef] [PubMed]
- Mei, D.A.; Gross, G.J.; Nithipatikom, K. Simultaneous determination of adenosine, inosine, hypoxanthine, xanthine, and uric acid in microdialysis samples using microbore column high-performance liquid chromatography with a diode array detector. Anal. Biochem. 1996, 238, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.F.; Bruce-Keller, A.J.; Goodman, Y.; Mattson, M.P. Uric Acid Protects Neurons Against Excitotoxic and Metabolic Insults in Cell Culture, and Against Focal Ischemic Brain Injury In Vivo. J. Neurosci. Res. 1998, 53, 613–625. [Google Scholar] [CrossRef]
- Fu, W.; Wu, Z.; Zheng, R.; Yin, N.; Han, F.; Zhao, Z.; Dai, M.; Han, D.; Wang, W.; Niu, L. Inhibition mechanism of melanin formation based on antioxidant scavenging of reactive oxygen species. Analyst 2022, 147, 2703–2711. [Google Scholar] [CrossRef] [PubMed]
- Chandler, C.J.; Wang, T.T.; Halsted, C. Pteroylpolyglutamate Hydrolase from Human Jejunal Brush Borders. J. Biol. Chem. 1986, 261, 928–933. [Google Scholar] [CrossRef]
- Ziegler, I.; McDonaldo, T.; Hesslinger, C.; Pelletier, I.; Boyle, P. Development of the Pteridine Pathway in the Zebrafish, Danio rerio. J. Biol. Chem. 2000, 275, 18926–18932. [Google Scholar] [CrossRef] [PubMed]
- Ronau, J.A.; Paul, L.N.; Fuchs, J.E.; Liedl, K.R.; Abu-Omar, M.M.; Das, C. A Conserved Acidic Residue in Phenylalanine Hydroxylase Contributes to Cofactor Affinity and Catalysis. Biochemistry. 2014, 53, 6834–6848. [Google Scholar] [CrossRef]
- Ichinose, H.; Nomura, T.; Sumi-Ichinose, C. Metabolism of tetrahydrobiopterin: Its relevance in monoaminergic neurons and neurological disorders. Chem. Rec. 2008, 8, 378–385. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets | |||
|---|---|---|---|
| Ingredients (%) | C | AS | HP |
| Fish meal | 60 | 60 | 60 |
| Wheat gluten flour | 5 | 5 | 5 |
| Soybean meal | 5 | 5 | 5 |
| Seaweed powder | 3 | 3 | 3 |
| Wheat meal | 14.5 | 14.5 | 14.5 |
| Soybean lecithin | 1 | 1 | 1 |
| Fish oil | 4 | 4 | 4 |
| Ca(H2PO4)2 | 1.5 | 1.5 | 1.5 |
| Vitamin premix and mineral premix | 1 | 1 | 1 |
| Microcrystalline cellulose | 5 | 4.8 | 4.333 |
| Carophyll® pink powder | 0 | 0.2 | 0 |
| Haematococcus lacustris powder | 0 | 0 | 0.667 |
| Total | 100 | 100 | 100 |
| Proximate composition (%, dry matter) | |||
| Moisture | 7.3 | 7.8 | 7.4 |
| Crude protein | 53.4 | 52.7 | 52.2 |
| Lipids | 10.8 | 10.7 | 11.2 |
| Ash | 12.2 | 13.4 | 13.3 |
| Astaxanthin | 0 | 0.0193 | 0.0186 |
| Treatment | WGR (%) | SGR (%/Day) | |
|---|---|---|---|
| 7 weeks | C | 33.91 ± 3.94 a | 0.59 ± 0.06 a |
| AS | 33.71 ± 15.70 a | 0.55 ± 0.24 a | |
| HP | 39.82 ± 8.00 a | 0.67 ± 0.11 a | |
| 14 weeks | C | 66.07 ± 3.24 a | 0.52 ± 0.02 a |
| AS | 76.04 ± 9.92 a | 0.57 ± 0.06 a | |
| HP | 98.94 ± 8.05 b | 0.70 ± 0.04 b |
| Comparison | Gene ID | Gene Name | Gene Annotation | Log2FC | Regulated |
|---|---|---|---|---|---|
| HP vs. C | gene-LOC121962187 | fabp2 | fatty-acid-binding protein, intestinal-like | −2.50 | down |
| gene-fabp2 | fabp2 | fatty-acid-binding protein, intestinal | −1.85 | down | |
| gene-creb3l1 | creb3l1 | cyclic AMP-responsive element-binding protein 3-like protein 1 | 1.09 | up | |
| gene-abcc6a | abcc6a | multidrug-resistance-associated protein 1 | 1.36 | up | |
| gene-rdh12 | rdh12 | retinol dehydrogenase 12-like | −1.65 | down | |
| AS vs. C | gene-tyrp1b | tyrp1b | 5,6-dihydroxyindole-2-carboxylic acid oxidase-like | 1.91 | up |
| gene-pmela | pmela | melanocyte protein PMEL-like | 1.35 | up | |
| HP vs. AS | gene-LOC121950639 | pnpla2 | patatin-like phospholipase-domain-containing protein 2 | −1.60 | down |
| gene-pmela | pmela | melanocyte protein PMEL-like | −1.21 | down | |
| gene-LOC121963361 | pmelb | premelanosome protein b | −1.06 | down | |
| gene-LOC121962312 | tyrp1 | 5,6-dihydroxyindole-2-carboxylic acid oxidase | −1.36 | down | |
| gene-tyrp1b | tyrp1b | 5,6-dihydroxyindole-2-carboxylic acid oxidase-like | −1.58 | down |
| Group | Ion Mode | Up | Down | All |
|---|---|---|---|---|
| HP vs. C | POS | 188 | 11 | 199 |
| AS vs. C | POS | 142 | 15 | 157 |
| HP vs. AS | POS | 75 | 30 | 105 |
| HP vs. C | NEG | 170 | 8 | 178 |
| AS vs. C | NEG | 84 | 8 | 92 |
| HP vs. AS | NEG | 59 | 15 | 74 |
| Metabolite ID | Name | Mass Error (ppm) | m/z | RT (min) |
|---|---|---|---|---|
| neg_5462 | 16-Hydroxyhexadecanoic acid | 3.66 | 271.23 | 7.93 |
| neg_1491 | 2-Hydroxybutyric acid | 15.15 | 103.04 | 0.85 |
| neg_3002 | Sepiapterin | −0.61 | 511.14 | 1.43 |
| neg_2991 | Uridine | 6.13 | 243.06 | 1.42 |
| Comparison | ko_ID | KEGG Pathway | Ion Mode | p-Value |
|---|---|---|---|---|
| HP vs. C | ko00592 | α-Linolenic acid metabolism | POS | 4.24 × 10−3 |
| ko04977 | Vitamin digestion and absorption | POS | 3.24 × 10−2 | |
| AS vs. C | ko00040 | Pentose and glucuronate interconversions | POS | 3.60 × 10−2 |
| ko00480 | Glutathione metabolism | NEG | 2.04 × 10−2 | |
| ko04216 | Ferroptosis | NEG | 2.04 × 10−2 | |
| ko04918 | Thyroid hormone synthesis | NEG | 2.04 × 10−2 | |
| HP vs. AS | ko01054 | Nonribosomal peptide structures | POS | 3.90 × 10−2 |
| ko01210 | 2-Oxocarboxylic acid metabolism | POS | 4.97 × 10−2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Tian, C.; Zhu, K.; Liu, Y.; Zhao, C.; Jiang, M.; Zhu, C.; Li, G. Effects of Natural and Synthetic Astaxanthin on Growth, Body Color, and Transcriptome and Metabolome Profiles in the Leopard Coralgrouper (Plectropomus leopardus). Animals 2023, 13, 1252. https://doi.org/10.3390/ani13071252
Zhang J, Tian C, Zhu K, Liu Y, Zhao C, Jiang M, Zhu C, Li G. Effects of Natural and Synthetic Astaxanthin on Growth, Body Color, and Transcriptome and Metabolome Profiles in the Leopard Coralgrouper (Plectropomus leopardus). Animals. 2023; 13(7):1252. https://doi.org/10.3390/ani13071252
Chicago/Turabian StyleZhang, Junpeng, Changxu Tian, Kecheng Zhu, Yong Liu, Can Zhao, Mouyan Jiang, Chunhua Zhu, and Guangli Li. 2023. "Effects of Natural and Synthetic Astaxanthin on Growth, Body Color, and Transcriptome and Metabolome Profiles in the Leopard Coralgrouper (Plectropomus leopardus)" Animals 13, no. 7: 1252. https://doi.org/10.3390/ani13071252
APA StyleZhang, J., Tian, C., Zhu, K., Liu, Y., Zhao, C., Jiang, M., Zhu, C., & Li, G. (2023). Effects of Natural and Synthetic Astaxanthin on Growth, Body Color, and Transcriptome and Metabolome Profiles in the Leopard Coralgrouper (Plectropomus leopardus). Animals, 13(7), 1252. https://doi.org/10.3390/ani13071252