
Molecular Cloning, Screening of Single Nucleotide Polymorphisms, and Analysis of Growth-Associated Traits of igf2 in Spotted Sea Bass (Lateolabrax maculatus)

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Extraction of Total RNA and cDNA Synthesis
2.3. Cloning igf2 from cDNA
2.4. Cloning of igf2 from Genomic DNA
2.5. Sequence Analysis
2.6. Identification of SNPs and Statistical Analyses
3. Results
3.1. cDNA Cloning and Characterization of igf2
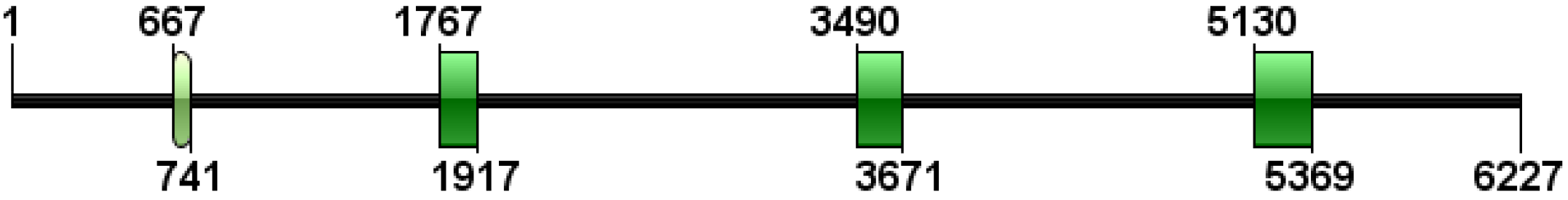
3.2. Characterization of the Genome Sequence of igf2
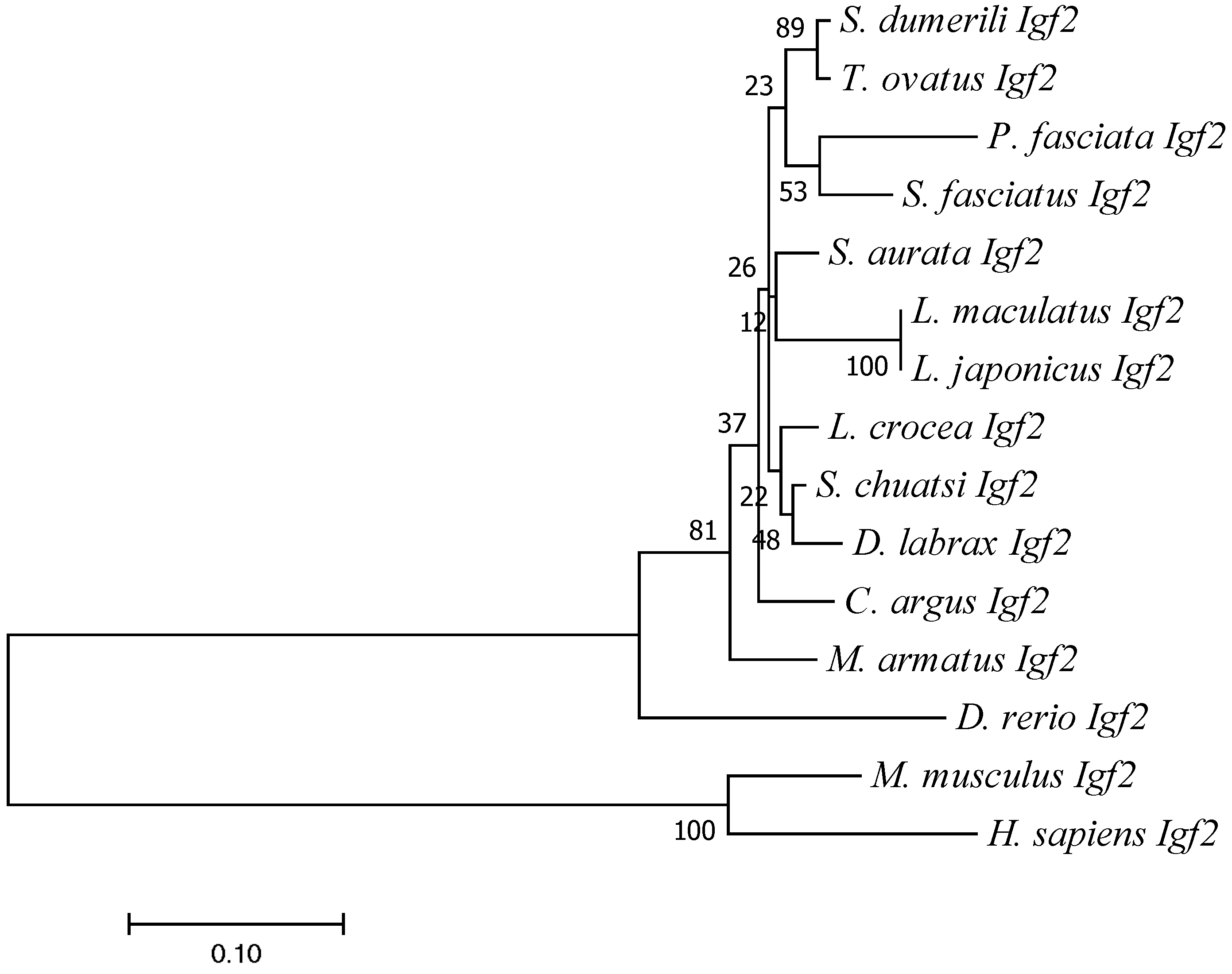
3.3. Sequence Identity and Phylogenetic Analysis of Igf2
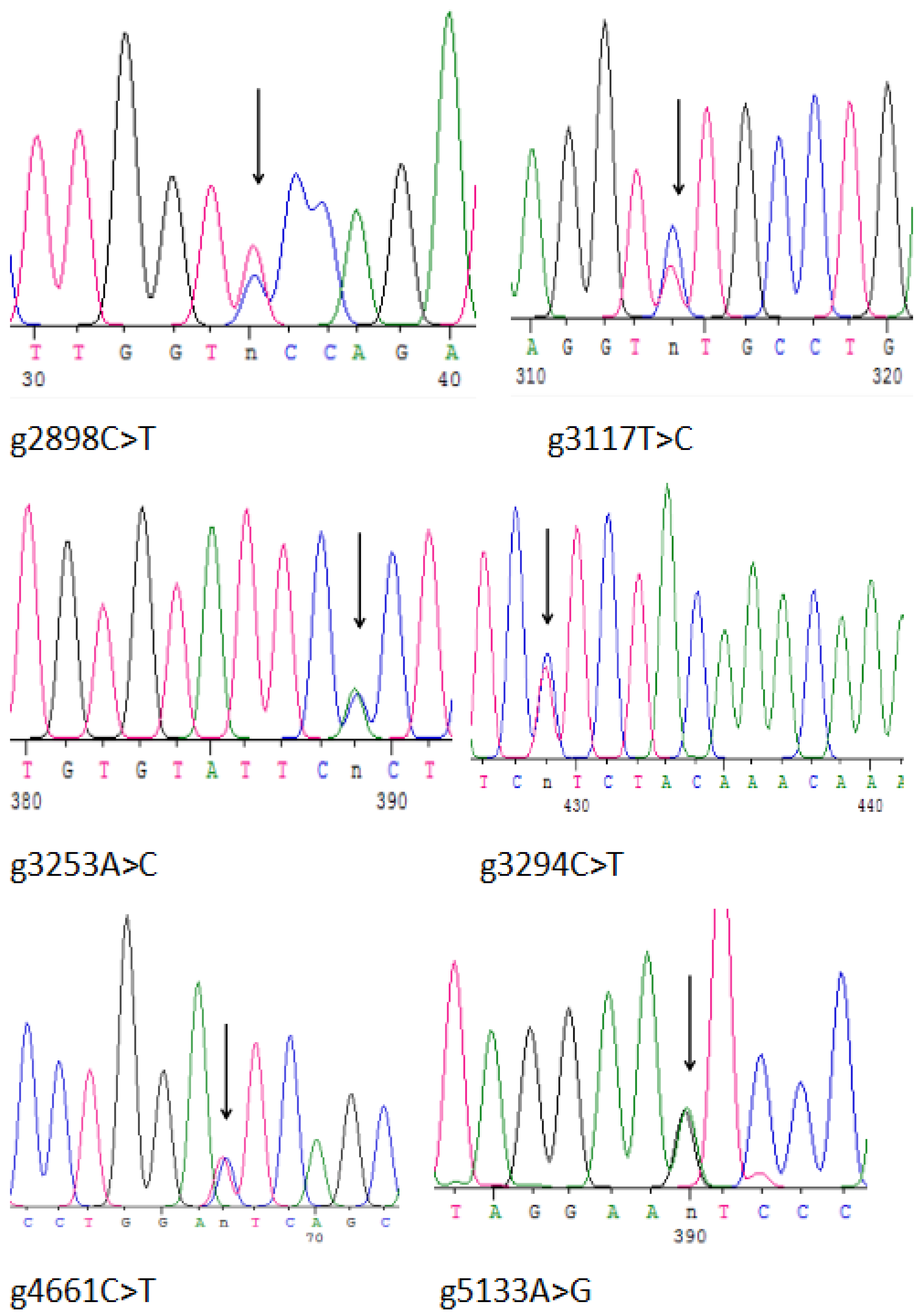
3.4. Detection of SNPs
3.5. Association of SNPs with Growth Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chandhini, S.; Trumboo, B.; Jose, S.; Varghese, T.; Rajesh, M.; Kumar, V. Insulin-like growth factor signalling and its significance as a biomarker in fish and shellfish research. Fish Physiol. Biochem. 2021, 47, 1011–1031. [Google Scholar] [CrossRef]
- Chu, M.X.; Jia, Y.; Wu, Z.; Huan, H.; Guo, X.; Yin, S.; Zhang, K. Genome-wide characterization of three IGFs in hybrid yellow catfish (Pseudobagrus fulvidraco ♀× Pseudobagrus vachellii ♂) and the association of IGF2 allelic variants with growth traits. Aquac. Rep. 2022, 26, 101315. [Google Scholar] [CrossRef]
- Zeng, N.; Bao, J.; Shu, T.; Shi, C.; Zhai, G.; Jin, X.; He, J.; Lou, Q.; Yin, Z. Sexual dimorphic effects of igf1 deficiency on metabolism in zebrafish. Front. Endocrinol. 2022, 13, 879962. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Z.; Kang, T.; Li, M.; Wang, D.; Cheng, C.H.K. Igf3: A novel player in fish reproductiont. Biol. Reprod. 2021, 104, 1194–1204. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Peng, L.B.; Wang, D.; Zhu, Q.L.; Zheng, J.L. Combined effects of polystyrene microplastics and cadmium on oxidative stress, apoptosis, and GH/IGF axis in zebrafish early life stages. Sci. Total Environ. 2022, 813, 152514. [Google Scholar] [CrossRef]
- Jones, J.I.; Clemmons, D.R. Insulin-like growth factors and their binding proteins: Biological actions. Endocr. Rev. 1995, 16, 3–34. [Google Scholar]
- Dixit, M.; Poudel, S.B.; Yakar, S. Effects of GH/IGF axis on bone and cartilage. Mol. Cell. Endocrinol. 2021, 519, 111052. [Google Scholar] [CrossRef]
- Gökçek, E.Ö.; Işık, R. Associations between genetic variants of the insulin-like growth factor I (IGF-I) gene and growth traits in European Sea bass (Dicentrarchus labrax, L.). Fish Physiol. Biochem. 2020, 46, 1131–1138. [Google Scholar] [CrossRef]
- Shamblott, M.J.; Chen, T.T. Identification of a second insulin-like growth factor in a fish species. Proc. Natl. Acad. Sci. USA 1992, 89, 8913–8917. [Google Scholar] [CrossRef]
- Chen, J.Y.; Chang, C.Y.; Chen, J.C.; Shen, S.C.; Wu, J.L. Production of biologically active recombinant tilapia insulin-like growth factor-II polypeptides in Escherichia coli cells and characterization of the genomic structure of the coding region. DNA Cell Biol. 1997, 16, 883–892. [Google Scholar] [CrossRef]
- Tse, M.C.; Vong, Q.P.; Cheng, C.H.; Chan, K.M. PCR-cloning and gene expression studies in common carp (Cyprinus carpio) insulin-like growth factor-II. Biochim. Biophys. Acta (BBA)-Gene Struct. Expr. 2002, 1575, 63–74. [Google Scholar] [CrossRef]
- Hartnett, L.; Glynn, C.; Nolan, C.M.; Grealy, M.; Byrnes, L. Insulin-like growth factor-2 regulates early neural and cardiovascular system development in zebrafish embryos. Int. J. Dev. Biol. 2010, 54, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Hong, Y. Medaka insulin-like growth factor-2 supports self-renewal of the embryonic stem cell line and blastomeres in vitro. Sci. Rep. 2017, 7, 78. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.N.; Jiang, X.Y.; Pu, J.W.; Li, Z.R.; Zou, S.M. Functional conservation and divergence of duplicated insulin-like growth factor 2 genes in grass carp (Ctenopharyngodon idellus). Gene 2011, 470, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Zou, S.; Kamei, H.; Modi, Z.; Duan, C. Zebrafish IGF genes:gene duplication, conservation and divergence, and novel roles in midline and notochord development. PLoS ONE 2009, 4, e7026. [Google Scholar] [CrossRef]
- Hoegg, S.; Brinkmann, H.; Taylor, J.S.; Meyer, A. Phylogenetic timing of the fish-specific genome duplication correlates with the diversification of teleost fish. J. Mol. Evol. 2004, 59, 190–203. [Google Scholar] [CrossRef]
- Macqueen, D.J.; Johnston, I.A. A well-constrained estimate for the timing of the salmonid whole genome duplication reveals major decoupling from species diversification. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132881. [Google Scholar] [CrossRef]
- Hollenbeck, C.M.; Johnston, I.A. Genomic Tools and Selective Breeding in Molluscs. Front. Genet. 2018, 9, 253. [Google Scholar] [CrossRef]
- Fan, J.; Ma, D.; Zhu, H.; Jiang, P.; Su, H. Gene structure, SNP screening and growth correlation analysis of the preproinsulin gene in grass carp (Ctenopharyngodon idellus). J. Genet. 2021, 100, 48. [Google Scholar] [CrossRef]
- Tsai, H.Y.; Hamilton, A.; Guy, D.R.; Houston, R.D. Single nucleotide polymorphisms in the insulin-like growth factor 1(IGF1) gene are associated with growth related traits in farmed Atlantic salmon. Anim. Genet. 2014, 45, 709–715. [Google Scholar] [CrossRef]
- Shao, C.; Li, C.; Wang, N.; Qin, Y.; Xu, W.; Liu, Q.; Zhou, Q.; Zhao, Y.; Li, X.; Liu, S.; et al. Chromosome-level genome assembly of the spotted sea bass, Lateolabrax maculatus. GigaScience 2018, 7, giy114. [Google Scholar] [CrossRef]
- Yan, L.; Wang, P.; Zhao, C.; Fan, S.; Lin, H.; Guo, Y.; Ma, Z.; Qiu, L. Toxic responses of liver in Lateolabrax maculatus during hypoxia and re-oxygenation. Aquat. Toxicol. 2021, 236, 105841. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Shi, X.; Liu, Z.; Ren, W.; Wang, X.; Huang, B.; Kou, X.; Liang, H.; Wang, C.; Chai, W. A Novel A > G Polymorphism in the Intron 1 of LCORL Gene Is Significantly Associated with Hide Weight and Body Size in Dezhou Donkey. Animals 2022, 12, 2581. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Bai, J.; Hu, Y.; Ye, X.; Li, S.; Yu, L. Genotypes, haplotypes and diplotypes of IGF-II SNPs and their association with growth traits in largemouth bass (Micropterus salmoides). Mol. Biol. Rep. 2012, 39, 4359–4365. [Google Scholar] [CrossRef] [PubMed]
- Tse, M.C.; Chan, K.M.; Cheng, C.H. Cloning, characterization and promoter analysis of the common carp IGF-II gene. Gene 2008, 412, 26–38. [Google Scholar] [CrossRef]
- Su, S.Y.; Dong, Z.J.; Qu, J.Q.; Liang, Z.Y.; Zhang, J.Q.; Ma, L.X.; Liu, W.; Xu, P.; Yuan, X.H. Molecular cloning and single nucleotide polymorphism analysis of IGF2a genes in the common carp (Cyprinus carpio). Genet. Mol. Res. 2012, 11, 1327–1340. [Google Scholar] [CrossRef]
- Palamarchuk, A.; Gritsenko, O.; Holthuizen, E.; Sussenbach, J.; Caelers, A.; Reinecke, M.; Kavsan, V. Complete nucleotide sequence of the chum salmon insulin-like growth factor II gene. Gene 2002, 295, 223–230. [Google Scholar] [CrossRef]
- Yu, J.; Chen, X.; Li, J.; Tang, Y.; Li, H.; Xu, P.; Dong, Z. Isolation of IGF2 and association of IGF2 polymorphism with growth trait in genetically improved farmed tilapias, Oreochromis niloticus L. Aquac. Res. 2010, 41, e743–e750. [Google Scholar]
- Collet, C.; Candy, J.; Richardson, N.; Sara, V. Organization, sequence, and expression of the gene encoding IGFII from barramundi (Teleosteii; Lates calcarifer). Biochem. Genet. 1997, 35, 211–224. [Google Scholar] [CrossRef]
- Teng, T.; Zhao, X.; Li, C.; Guo, J.; Wang, Y.; Pan, C.; Liu, E.; Ling, Q. Cloning and expression of IGF-I, IGF-II, and GHR genes and the role of their single-nucleotide polymorphisms in the growth of pikeperch (Sander lucioperca). Aquacult. Int. 2020, 28, 1547–1561. [Google Scholar] [CrossRef]
- Abo-Al-Ela, H.G.; El-Magd, M.A.; El-Nahas, A.F.; Mansour, A.A. Association of a novel SNP in exon 10 of the IGF2 gene with growth traits in Egyptian water buffalo (Bubalus bubalis). Trop. Anim. Health Prod. 2014, 46, 947–952. [Google Scholar] [CrossRef] [PubMed]
- Sélénou, C.; Brioude, F.; Giabicani, E.; Sobrier, M.L.; Netchine, I. IGF2: Development, Genetic and Epigenetic Abnormalities. Cells 2022, 11, 1886. [Google Scholar] [CrossRef] [PubMed]
- Devran, Z.; Kahveci, E.; Hong, Y.; Studholme, D.J.; Tör, M. Identifying molecular markers suitable for Frl selection in tomato breeding. TAG Theor. Appl. Genet. 2018, 131, 2099–2105. [Google Scholar] [CrossRef] [PubMed]
- Mao, C.; Zhang, T.; Ren, A.; Jia, X.; Lai, S.J.; Chen, S.Y. Genome-wide SNP discovery and genetic diversity evaluation of Liangshan cattle in China. Anim. Biotechnol. 2021, 32, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Z.; Zhan, Z.Y.; Li, X.Y.; Wu, S.R.; Sun, Y.J.; Xue, J.; Lan, X.Y.; Lei, C.Z.; Zhang, C.L.; Jia, Y.T.; et al. SNP and haplotype analysis reveal IGF2 variants associated with growth traits in Chinese Qinchuan cattle. Mol. Biol. Rep. 2014, 41, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.C.; Li, X.Y.; Su, S.Y.; Liang, J.D.; Xu, P.; He, X.J. Bouzoualegh R.Construction of SNPs from common carp IGF2b and its application in Huanghe carp new strain (Cyprinus carpio hacmalopterus Temminck et Schlegel). J. Fish. Sci. China 2019, 26, 686–694, (Abastract in English only). [Google Scholar]
- Mi, T.; Liu, K.; Guo, T.; Li, L.; Wang, Y.; Li, C.; Cui, Y.; Dai, J.; Zhang, Y.; Hu, S. Analysis of the eighth intron polymorphism of NR6A1 gene in sheep and its correlation with lumbar spine number. Anim. Biotechnol. 2021, 7, 1–7, advance online publication. [Google Scholar] [CrossRef]
- Komar, A.A. Silent SNPs: Impact on gene function and phenotype. Pharmacogenomics 2007, 8, 1075–1080. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yu, X.; Tong, J. Polymorphisms in Myostatin Gene and associations with growth traits in the common carp (Cyprinus carpio L.). Int. J. Mol. Sci. 2012, 13, 14956–14961. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5’→3’) | Product Length (bp) | Purpose |
|---|---|---|---|
| cs | CCGCACTAAAACAGGAAATC | 129 | Verification of cDNA sequence |
| ca | TCCTTTTAGCGATCAGGATG | ||
| gs | ATAAGAGCCTACTTGTCCCT | 3901 | Verification of DNA sequence |
| ga | GGCAATTCAGACAGAAGGTA | ||
| E1s | TGAGAATGCACTTGAGTCAG | 714 | SNP identification at 108-821 bp |
| E1a | CGTTTTAACCTCACAACTGG | ||
| E2s | CCTGTTATGAGAGGCAACAT | 953 | SNP identification at 1558-2510 bp |
| E2a | GTTGATGTTAATGATCCGCC | ||
| E3s | GTTCCTCTACCTGCAATCAA | 1036 | SNP identification at 2826-3861 bp |
| E3a | GGAAGGTAACTGCTTACCAA | ||
| E4s | GGGGATAAACACTCCTCAAG | 1677 | SNP identification ay 4542-6218 bp |
| E4a | CAACCTCTTCTTTGAGTCCA |
| Latin Name | Accession Number | Identity (%) |
|---|---|---|
| Lateolabrax maculatus | ON462263 | – |
| Lateolabrax japonicus | AEX60713.2 | 100 |
| Siniperca chuatsi | ADO14143.1 | 93 |
| Seriola dumerili | XP_022609942.1 | 93 |
| Trachinotus ovatus | ANJ77685.1 | 93 |
| Larimichthys crocea | XP_019124122.1 | 92 |
| Salarias fasciatus | XP_029951810.1 | 92 |
| Sparus aurata | ABQ52655.1 | 92 |
| Dicentrarchus labrax | AAW29441.1 | 90 |
| Channa argus | KAF3689021.1 | 88 |
| Mastacembelus armatus | XP_026167195.1 | 87 |
| Poeciliopsis fasciata | ABC60243.1 | 87 |
| Danio rerio | NP_001001815.1 | 76 |
| Mus musculus | AAH53489.1 | 47 |
| Homo sapiens | AAB34155.1 | 44 |
| Locus | Position | Genotype and Number | Genotype Frequency |
|---|---|---|---|
| g2898C>T | Intron 2 | CC(153)/TT(12)/TC(25) | 0.81/0.06/0.13 |
| g2907C>T | Intron 2 | CC(153)/TC(37) | 0.81/0.19 |
| g3016A>C | Intron 2 | AA(165)/CC(22)/AC(3) | 0.92/0.11/0.02 |
| g3117T>C | Intron 2 | TT(178)/CC(2)/TC(10) | 0.94/0.01/0.05 |
| g3230A>C | Intron 2 | AA(136)/CC(9)/AC(45) | 0.72/0.05/0.23 |
| g3253A>C | Intron 2 | AA(111)/CC(26)/AC(53) | 0.58/0.14/0.28 |
| g3294C>T | Intron 2 | TT(81)/CC(97)/TC(12) | 0.43/0.51/0.06 |
| g3329G>A | Intron 2 | AA(19)/GG(171) | 0.10/0.90 |
| g3435C>A | Intron 2 | AA(11)/CC(179) | 0.06/0.94 |
| g3474T>G | Intron 2 | TT(175)/GG(15) | 0.92/0.08 |
| g4661C>T | Intron 3 | CC(149)/TT(10)/TC(28) | 0.80/0.05/0.15 |
| g4927A>G | Intron 3 | GG(23)/AA(167) | 0.12/0.88 |
| g5064C>T | Intron 3 | CC(131)/TT(59) | 0.69/0.31 |
| g5133A>G | Exon 4 | AA(140)/GG(26)/GA(24) | 0.74/0.14/0.12 |
| Locus | Genotype | BM (g) | SL (cm) | BD (mm) | BW (mm) | HL (mm) | TL (cm) |
|---|---|---|---|---|---|---|---|
| g2907C>T | CC | 586.34 ± 133.16 | 33.29 ± 2.31 | 82.52 ± 11.63 a | 43.08 ± 8.08 | 92.30 ± 9.75 a | 36.33 ± 2.56 |
| TC | 594.11 ± 104.86 | 33.29 ± 2.01 | 86.90 ± 8.69 b | 44.56 ± 6.71 | 95.8181 ± 7.66 b | 36.72 ± 2.18 | |
| g3230A>C | AA | 595.23 ± 127.35 | 33.50 ± 2.21 | 83.47 ± 12.11 | 43.35 ± 8.28 | 93.28 ± 9.77 | 36.67 ± 2.39 a |
| CC | 641.84 ± 129.13 | 33.43 ± 1.48 | 86.11 ± 6.74 | 46.29 ± 5.35 | 95.12 ± 6.91 | 37.72 ± 2.30 a | |
| AC | 564.70 ± 131.72 | 32.79 ± 2.50 | 82.58 ± 8.89 | 42.87 ± 6.60 | 91.51 ± 8.82 | 35.56 ± 2.76 b | |
| g3294C>T | CC | 580.76 ± 137.30 | 33.32 ± 2.23 | 81.81 ± 11.72 a | 41.53 ± 9.24 a | 91.31 ± 9.94 a | 36.07 ± 2.53 |
| TC | 578.12 ± 96.09 | 33.26 ± 1.47 | 81.13 ± 13.01 ab | 44.02 ± 5.51 ab | 90.17 ± 10.10 ab | 36.62 ± 1.61 | |
| TT | 603.29 ± 122.90 | 33.36 ± 2.42 | 85.60 ± 10.02 b | 45.48 ± 5.34 b | 95.33 ± 8.24 b | 36.89 ± 2.58 | |
| g5064C>T | CC | 603.03 ± 134.40 | 33.61 ± 2.43 a | 83.73 ± 10.56 | 43.53 ± 7.91 | 92.94 ± 8.80 | 36.61 ± 2.65 |
| TT | 566.32 ± 117.03 | 32.86 ± 1.80 b | 82.22 ± 12.46 | 42.86 ± 7.95 | 92.49 ± 11.12 | 36.19 ± 2.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, S.; Wang, P.; Zhao, C.; Yan, L.; Zhang, B.; Qiu, L. Molecular Cloning, Screening of Single Nucleotide Polymorphisms, and Analysis of Growth-Associated Traits of igf2 in Spotted Sea Bass (Lateolabrax maculatus). Animals 2023, 13, 982. https://doi.org/10.3390/ani13060982
Fan S, Wang P, Zhao C, Yan L, Zhang B, Qiu L. Molecular Cloning, Screening of Single Nucleotide Polymorphisms, and Analysis of Growth-Associated Traits of igf2 in Spotted Sea Bass (Lateolabrax maculatus). Animals. 2023; 13(6):982. https://doi.org/10.3390/ani13060982
Chicago/Turabian StyleFan, Sigang, Pengfei Wang, Chao Zhao, Lulu Yan, Bo Zhang, and Lihua Qiu. 2023. "Molecular Cloning, Screening of Single Nucleotide Polymorphisms, and Analysis of Growth-Associated Traits of igf2 in Spotted Sea Bass (Lateolabrax maculatus)" Animals 13, no. 6: 982. https://doi.org/10.3390/ani13060982
APA StyleFan, S., Wang, P., Zhao, C., Yan, L., Zhang, B., & Qiu, L. (2023). Molecular Cloning, Screening of Single Nucleotide Polymorphisms, and Analysis of Growth-Associated Traits of igf2 in Spotted Sea Bass (Lateolabrax maculatus). Animals, 13(6), 982. https://doi.org/10.3390/ani13060982