
The Effect of Sexes and Seasons on the Morphological Structures of the Ruminant Digestive System of Blue Sheep (Pseudois nayaur)
 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
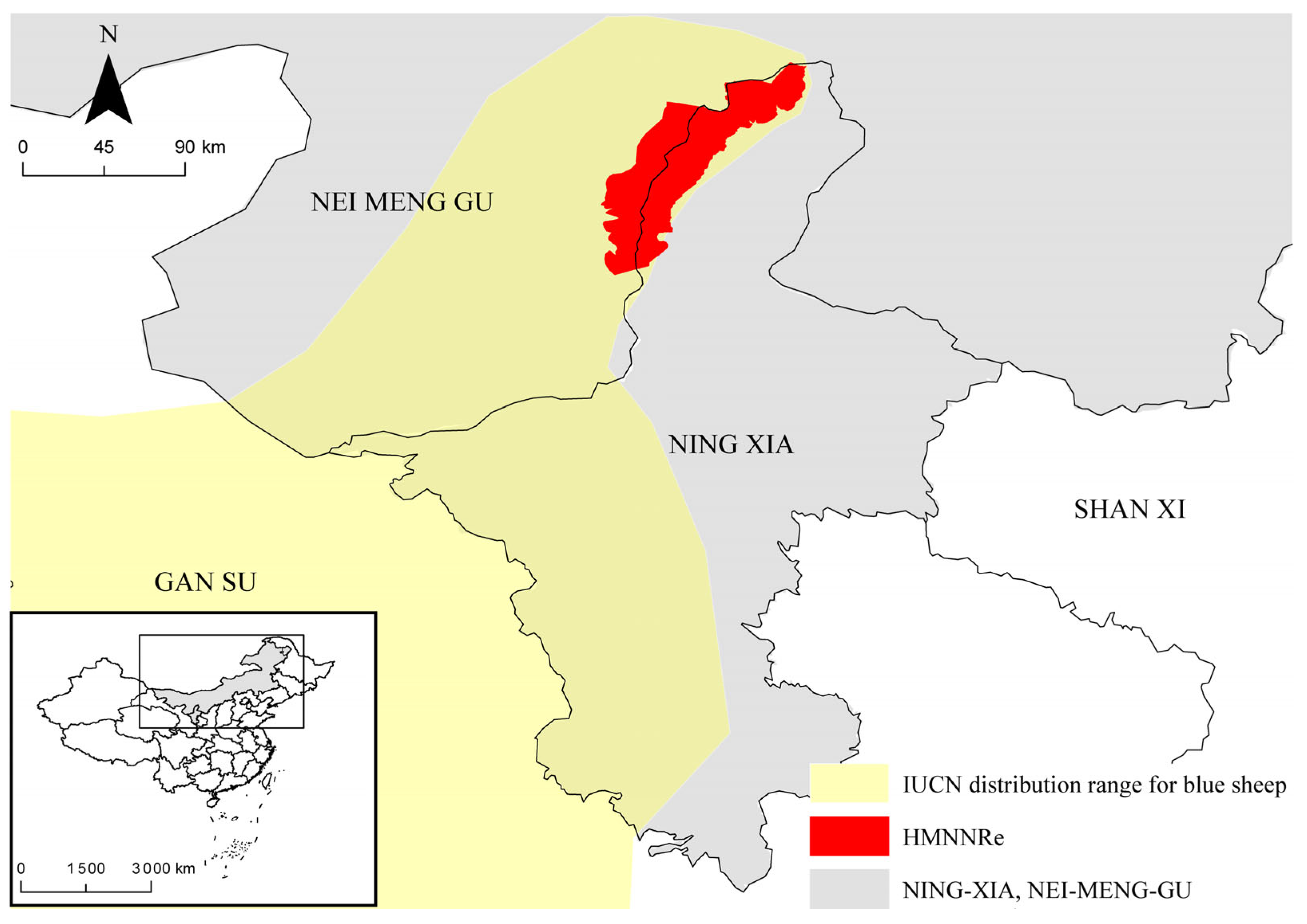
2.1. Study Area
2.2. Measurement of External Morphological Characteristics of Blue Sheep
2.3. Measurement of Rumen Surface Enlargement Factor
2.4. Distribution of Feed Particles in Different Parts of Digestive Tract of Blue Sheep
2.5. Statistical Analyses
3. Results
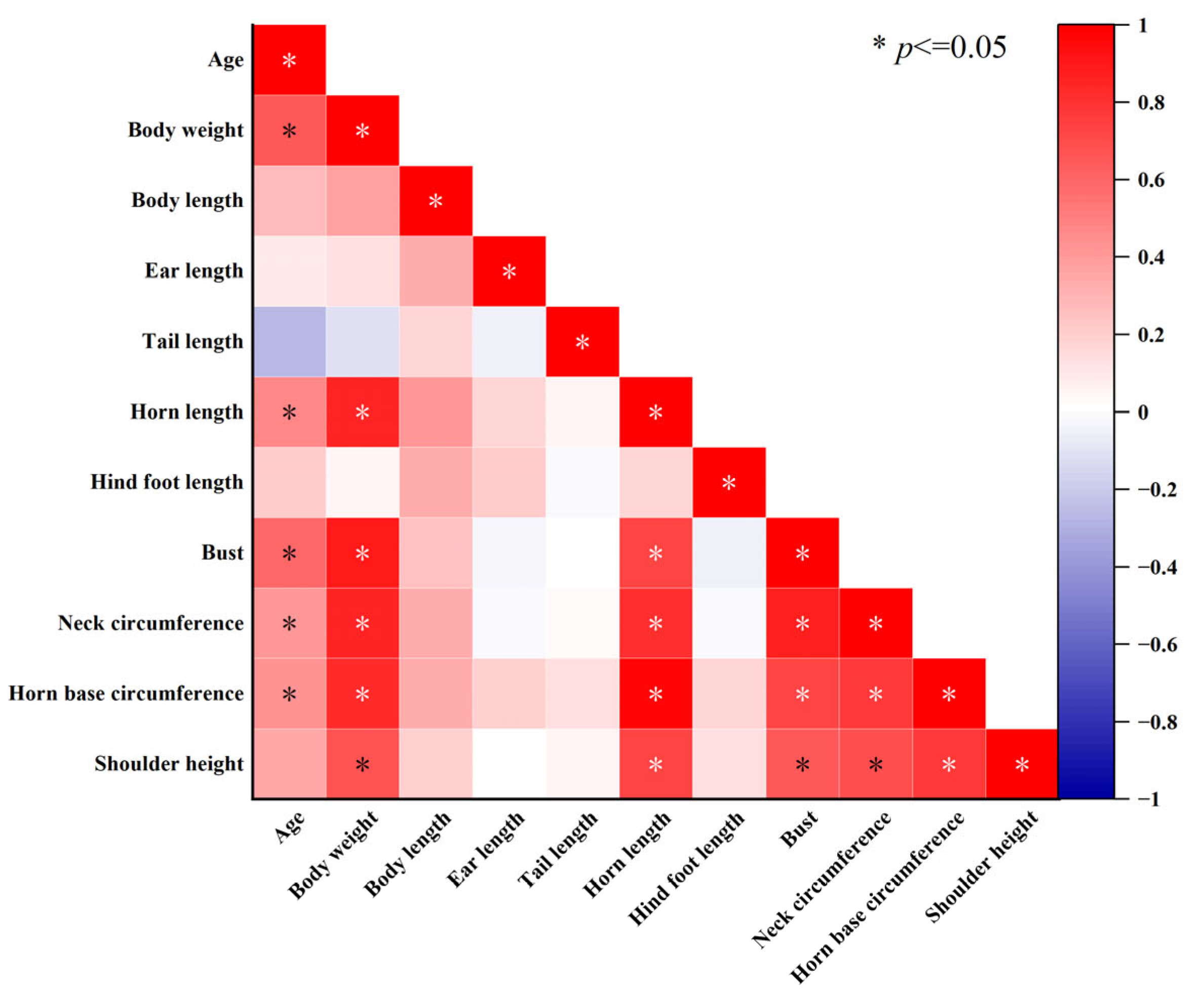
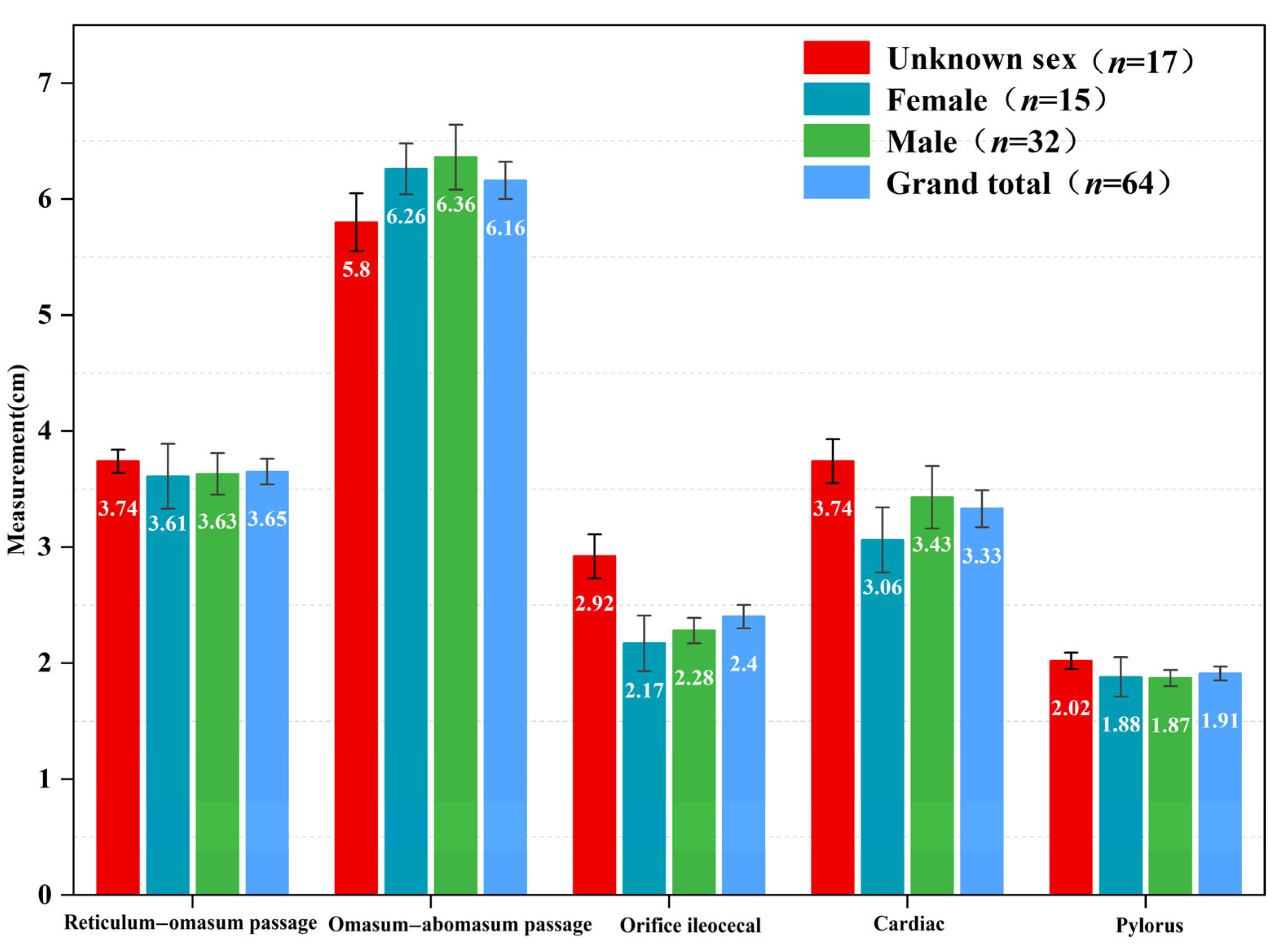
3.1. External Morphological Characteristics of Blue Sheep
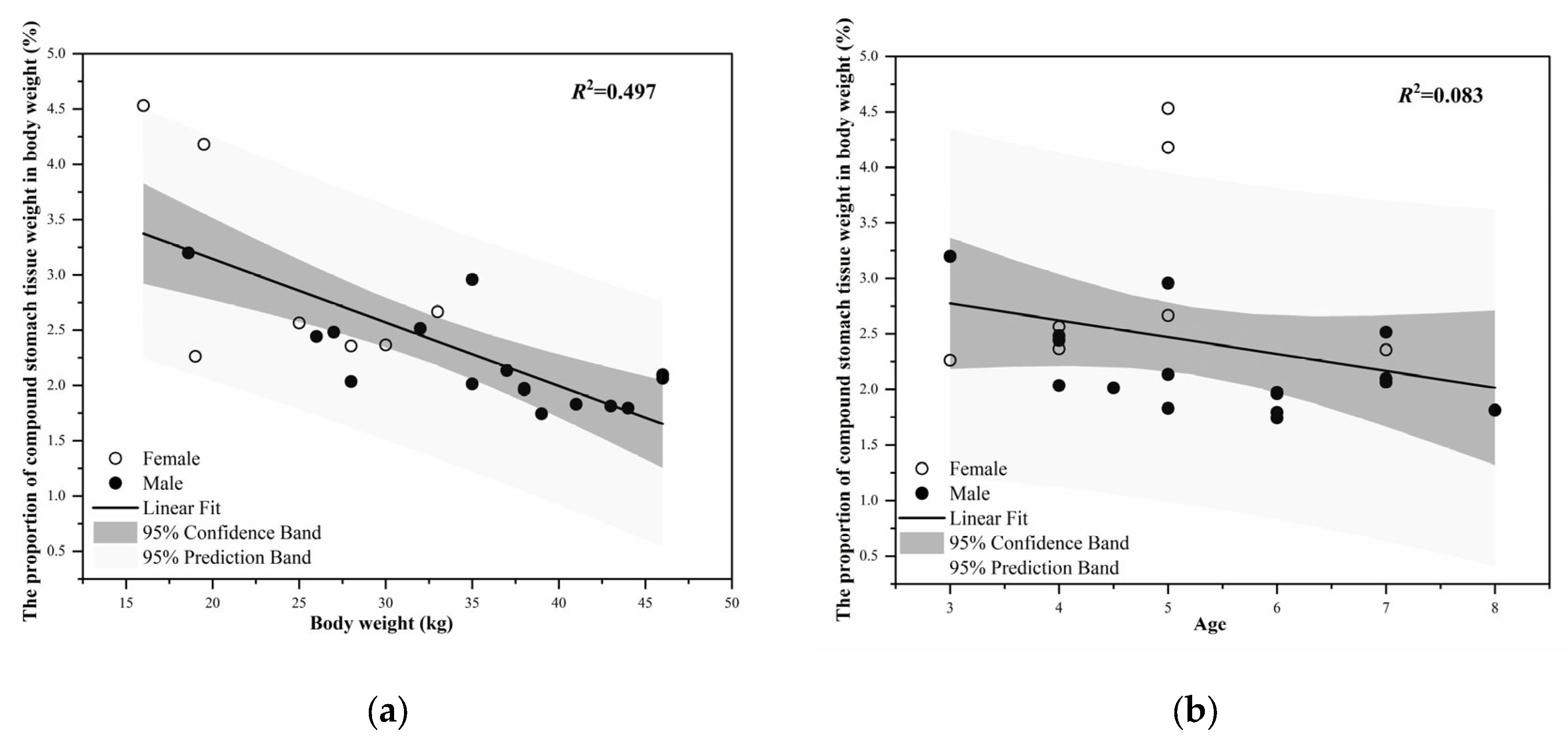
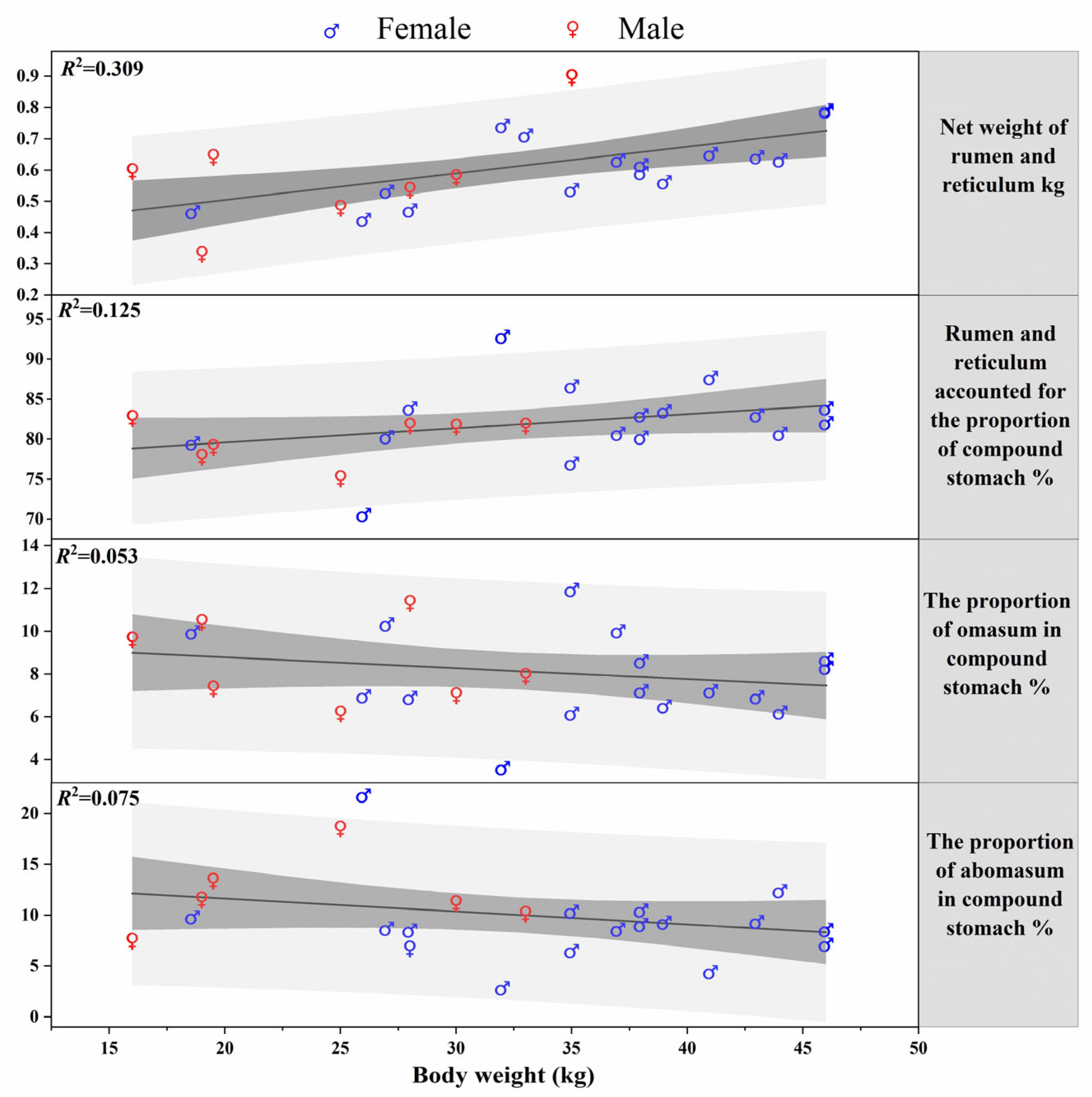
3.2. Morphology and Weight Change of the Stomach of Blue Sheep
3.3. Rumen Surface Enlargement Factor of Blue Sheep
3.4. Intestinal Length and Weight of Blue Sheep
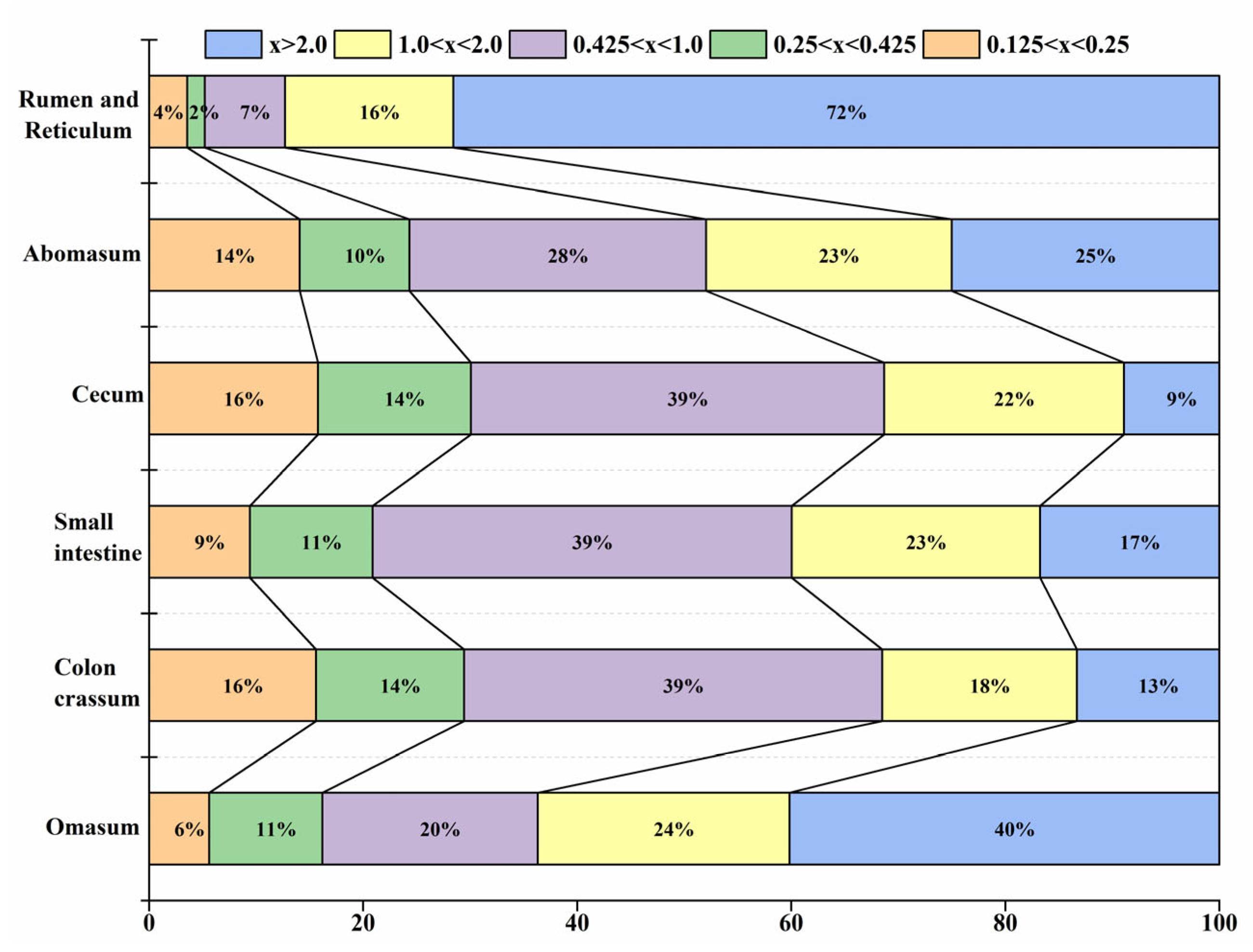
3.5. Distribution of Feed Particles in the Digestive Tract of Blue Sheep in Different Seasons
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chilliard, Y.; Doreau, M.; Veissier, I.; Bocquier, F. Guest editorial: Ruminant physiology; digestion, metabolism and effects of nutrition on reproduction and welfare. Animal 2010, 4, 977. [Google Scholar] [CrossRef] [PubMed]
- Tarso, S.; Daniela, O.; José, A.B.A. Ruminants as Part of the Global Food System: How Evolutionary Adaptations and Diversity of the Digestive System Brought them to the Future. Anim. Res. 2016, 3, 94–101. [Google Scholar] [CrossRef]
- Clauss, M.; Hume, I.D.; Hummel, J. Evolutionary adaptations of ruminants and their potential relevance for modern production systems. Animal 2010, 4, 979–992. [Google Scholar] [CrossRef]
- Hofmann, R.R. The Ruminant Stomach. Stomach Structure and Feeding Habits of East African Game Ruminants; East African Literature Bureau: Nairobi, Kenya, 1973. [Google Scholar]
- Przybyo, M.; Flaga, J.; Clauss, M.; Szczepanik, K.; Miltko, R.; Beecki, G.; Kowalski, Z.M.; Górka, P. Increased intake of mono-and disaccharides by Reeves’s muntjac (Muntiacus reevesi). Effect on gastrointestinal tract structure and function and blood parameters. J. Anim. Physiol. Anim. Nutr. 2022, 106, 922–938. [Google Scholar] [CrossRef] [PubMed]
- Paweł, G.; Bogdan, Ś.; Jadwiga, F.; Jarosław, O.; Marcin, W.; Klaudia, K.; Godlewski, M.M.; Romuald, Z.; Kowalski, Z.M. Effect of exogenous butyrate on the gastrointestinal tract of sheep. I. Structure and function of the rumen, omasum, and abomasum1. J. Anim. Sci. 2019, 98, 5311–5324. [Google Scholar] [CrossRef]
- Ehrlich, C.; Codron, D.; Hofmann, R.R.; Hummel, J.; Clauss, M. Comparative omasum anatomy in ruminants: Relationships with natural diet, digestive physiology, and general considerations on allometric investigations. J. Morphol. 2019, 280, 259–277. [Google Scholar] [CrossRef] [PubMed]
- Millen, D.D.; Arrigoni, M.D.B.; Pacheco, R.D.L. Rumenology; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- König, A.; Hudler, M.; Dahl, S.-A.; Bolduan, C.; Brugger, D.; Windisch, W. Response of roe deer (Capreolus capreolus) to seasonal and local changes in dietary energy content and quality. Anim. Prod. Sci. 2020, 60, 1315–1325. [Google Scholar] [CrossRef]
- Valerius, G. On the Relationship of Social Evolution and Ecology in Ungulates. Am. Zool. 1974, 14, 205–220. [Google Scholar] [CrossRef]
- Barboza, P.S.; Peltier, T.C.; Forster, R.J. Ruminal fermentation and fill change with season in an arctic grazer: Responses to hyperphagia and hypophagia in muskoxen (Ovibos moschatus). Physiol. Biochem. Zool. 2006, 79, 497–513. [Google Scholar] [CrossRef]
- Ramzinski, D.M.; Weckerly, F.W. Scaling Relationship between Body Weight and Fermentation Gut Capacity in Axis Deer. J. Mammal. 2007, 88, 415–420. [Google Scholar] [CrossRef]
- Weckerly, F.; Bhaskar, G.; Duarte, A.; Luna, R.S.; Starns, H.D. Heavier rumen–reticulum organs in white-tailed deer (Odocoileus virginianus) is consistent with dietary bulk not quality. Can. J. Zool. 2018, 96, 748–752. [Google Scholar] [CrossRef]
- Hofmann, R.R. Evolutionary Steps of Ecophysiological Adaptation and Diversification of Ruminants: A Comparative View of Their Digestive System. Oecologia 1989, 78, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Clauss, M.; Jürgen, H. Physiological adaptations of ruminants and their potential relevance for production systems. Rev. Bras. Zootec. 2017, 46, 606–613. [Google Scholar] [CrossRef]
- Smith, L.W.; Weinland, B.T.; Waldo, D.R.; Leffel, E.C. Rate of Plant Cell Wall Particle Size Reduction in the Rumen. J. Dairy Sci. 1983, 66, 2124–2136. [Google Scholar] [CrossRef]
- Soest, P.J.V. Nutritional Ecology of the Ruminant; Cornell University Press: Ithaca, NY, USA, 2018. [Google Scholar]
- Pei, Y.; Wang, D. Factors Influencing the Retention Time of Digesta in Mammals. Chin. J. Zool. 2000, 35, 50–53. [Google Scholar] [CrossRef]
- Pearce, G.R. Changes in particle size in the reticulorumen of sheep. Aust. J. Agric. Res. 1967, 18, 119–125. [Google Scholar] [CrossRef]
- Sun, G.; Zhang, H.; Wei, Q.; Zhao, C.; Li, Y. Comparative Analyses of Fecal Microbiota in European Mouflon (Ovis orientalis musimon) and Blue Sheep (Pseudois nayaur) Living at Low or High Altitudes. Front. Microbiol. 2019, 10, 1735. [Google Scholar] [CrossRef]
- Sheikh, K.M.; Molur, S. Status and Red List of Pakistan’s Mammals, Based on Conservation Assessment and Management Plan for Mammals; IUCN: Islamabad, Pakistan, 2005; Volume 344. [Google Scholar]
- Schaller, G.B. Mountain Monarchs: Wild Sheep and Goats of The Himalaya; University of Chicago Press: Chicago, IL, USA, 1977. [Google Scholar]
- Mishra, C.; Wieren, S.E.V.; Ketner, P.; Heitkönig, I.M.A.; Prins, H.H.T. Competition between domestic livestock and wild bharal Pseudois nayaur in the Indian Trans-Himalaya. J. Appl. Ecol. 2004, 41, 344–354. [Google Scholar] [CrossRef]
- Davies, N.B.; Krebs, J.R. An Introduction to Behavioural Ecology, 3rd ed.; Blackwell Scientific Publications: Cambridge, UK, 1993; pp. 48–76. [Google Scholar]
- Davies, N.B.; Krebs, J.R.; West, S.A. An Introduction to Behavioural Ecology, 4th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2012. [Google Scholar]
- Sheng, H. Fur Animal Handbook; Lexicographical Publishing House: Shanghai, China, 1994. [Google Scholar]
- Wang, X.; Liu, Z.; Xu, H.; Li, M.; Li, Y. The Blue Sheep population ecology and its conservation in Helan Mountain, China. Chin. Biodivers. 1998, 6, 1–5. [Google Scholar] [CrossRef]
- Namgail, T.; Fox, J.L.; Bhatnagar, Y.V. Habitat segregation between sympatric Tibetan argali Ovis ammon hodgsoni and blue sheep Pseudois nayaur in the Indian Trans-Himalaya. J. Zool. 2004, 262, 57–63. [Google Scholar] [CrossRef]
- Aryal, A.; Coogan, S.C.P.; Ji, W.; Rothman, J.M.; Raubenheimer, D. Foods, macronutrients and fibre in the diet of blue sheep (Psuedois nayaur) in the Annapurna Conservation Area of Nepal. Ecol. Evol. 2015, 5, 4006–4017. [Google Scholar] [CrossRef] [PubMed]
- Khattak, R.H.; Ali, H.; Rehman, E.U.; Nawaz, M.A. Population Structure of Blue Sheep (Pseudois nayaur) in Shimshal Valley Gilgit-Baltistan Pakistan. Pak. J. Zool. 2020, 52, 425–824. [Google Scholar] [CrossRef]
- Craig, H.T. 2000 IUCN Red List of Threatened Species; Species Survival Commission, IUCN: Gland, Switzerland, 2000. [Google Scholar]
- Wang, X.; Schaller, G.B. Status of large mammals in western Inner Mongolia, China. J. East China Norm. Univ. (Spec. Issue Mamm.) 1996, 6, 97–108. [Google Scholar]
- Wegge, P.; Shrestha, R.; Flagstad, Ø. Snow leopard Panthera uncia predation on livestock and wild prey in a mountain valley in northern Nepal: Implications for conservation management. Wildl. Biol. 2012, 18, 131–141. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, X.; Li, Z. The number and distribution of rock sheep in Helan Mountain. Chin. J. Zool. 2007, 43, 6–7. [Google Scholar] [CrossRef]
- Lu, H.; Li, Z.; Zhai, H.; Li, T.; Hu, T.; Zhao, C.; Wang, H.; Li, Y.; Shi, H.; Wang, Z.; et al. Monitoring and Survey of Blue sheep in Germany—Aided Project Area of state Natural Reserve of Helanshan Mountain in Ningxia. Ningxia J. Agric. For. 2000, S1, 15–18. [Google Scholar]
- Liu, Z.; Wang, X.; Teng, L.; Cui, D.; Li, X. Estimating seasonal density of blue sheep (Pseudois nayaur) in the Helan Mountain region using distance sampling methods. Ecol. Res. 2008, 23, 393–400. [Google Scholar] [CrossRef]
- Aryal, A.; Brunton, D.; Raubenheimer, D. Habitat assessment for the translocation of blue sheep to maintain a viable snow leopard population in the Mt Everest Region, Nepal. Zool. Ecol. 2013, 23, 66–82. [Google Scholar] [CrossRef]
- Aryal, A.; Brunton, D.; Ji, W.; Raubenheimer, D. Blue sheep in the Annapurna Conservation Area, Nepal: Habitat use, population biomass and their contribution to the carrying capacity of snow leopards. Integr. Zool. 2014, 9, 34–45. [Google Scholar] [CrossRef]
- Packer, J.G.; Takhtajan, A. Floristic Regions of the World. Bull. Torrey Bot. Club 1988, 115, 230. [Google Scholar] [CrossRef]
- Luo, Y. Compared Study on the Diet, Habitat Selection of Sympatric Blue Sheep (Pseudois nayaur) and Red Deer (Cervus elaphus alxaicus) in Helan Mountains, China. Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2011. [Google Scholar]
- Zhang, W.; Cui, Z.; Li, Y.; Wang, Z.; Yu, Y.; He, M. Characteristics of Quaternary glaciers in Helan Mountain and their coupling relationship with climate and structure. Chin. Sci. Bull. 2012, 57, 13. [Google Scholar] [CrossRef]
- Di, W. Plantae Vasculares Helanshanicae; Northwestern University Press: Xi’an, China, 1987. [Google Scholar]
- Di, Q.; Tian, L.; Li, Z. The characteristics of Helan Mountains flora of spermatophytes. Acta Bot. Boreali-Occident. Sin. 1988, 8, 242–254. [Google Scholar]
- Li, Z.; Wang, J.; Khattak, R.H.; Han, X.; Liu, P.; Liu, Z.; Teng, L. Coexistence mechanisms of sympatric ungulates: Behavioral and physiological adaptations of blue sheep (Pseudois nayaur) and red deer (Cervus elaphus alxaicus) in Helan Mountains, China. Front. Ecol. Evol. 2022, 10, 1–14. [Google Scholar] [CrossRef]
- Li, Z.; Pang, B.; Yao, Z.; Mi, S.; Liu, Z.; Teng, L. Microhabitat selection of blue eared pheasant (Crossoptilon auritum) during breeding period in Helan Mountains, China. Avian Biol. Res. 2022, 15, 173–179. [Google Scholar] [CrossRef]
- Ashby, K.R.; Henry, B.A.M. Age criteria and life expectancy of Roe deer (Capreolus capreolus) in coniferous forest in North-eastern England. Proc. Zool. Soc. Lond. 1979, 189, 207–220. [Google Scholar] [CrossRef]
- Xiong, B. Atlas of Sheep Solid Anatomy; China Agriculture Press: Beijing, China, 2012. [Google Scholar]
- Luna, R.; Duarte, A.; Weckerly, F. Rumen-reticulum characteristics, scaling relationships, and ontogeny in white-tailed deer (Odocoileus virginianus). Can. J. Zool. 2012, 90, 1351–1358. [Google Scholar] [CrossRef]
- McGrosky, A.; Codron, D.; Müller, D.W.H.; Navarrete, A.; Isler, K.; Hofmann, R.R.; Clauss, M. Gross intestinal morphometry and allometry in ruminants. J. Morphol. 2019, 280, 1254–1266. [Google Scholar] [CrossRef] [PubMed]
- Hemmoda, A.S.; Berg, R. Gross-anatomical studies on the ruminal pillar system of the Egyptian water buffalo (Bos bubalis L.). Anat. Histol. Embryol. 1980, 9, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, R.R. Morphophysiological Evolutionary Adaptations of the Ruminant Digestive System; Aspects of Digestive Physiology in Ruminants; Cornell University Press: Ithaca, NY, USA, 2019. [Google Scholar]
- Prins, R.A.; Geelen, M.J.J. Rumen characteristics of red deer, fallow deer, and roe deer. J. Wildl. Manag. 1971, 35, 673–680. [Google Scholar] [CrossRef]
- Ravimurugan, T.; Devendran, P. Body measurements and body weight of Ramnad White sheep. Indian J. Small Rumin. 2009, 15, 266–267. [Google Scholar]
- Ma, J.; Li, J.; Jiang, Z.; Wang, W. Morphological and structural characteristics of digestive tract of yellow sheep. Acta Theriol. Sin. 2001, 21, 14–22. [Google Scholar] [CrossRef]
- Hakonson, T.E.; Ward, W.F. The Contribution of Various Tissues and Organs to Total Body Mass in the Mule Deer. J. Mammal. 1971, 52, 628–630. [Google Scholar] [CrossRef]
- Short, H. Postnatal stomach development of white-tailed deer. J. Wildl. Manag. 1964, 28, 445–458. [Google Scholar] [CrossRef]
- Takatsuki, S. The weight contribution of stomach compartment of sika deer. J. Wildl. Manag. 1988, 52, 313–316. [Google Scholar] [CrossRef]
- Chivers, D.J.; Hladik, C.M. Morphology of the gastrointestinal tract in primates: Comparisons with other mammals in relation to diet. J. Morphol. 1980, 166, 337–386. [Google Scholar] [CrossRef]
- Ortigues, I.; Doreau, M. Responses of the splanchnic tissues of ruminants to changes in intake: Absorption of digestion end products, tissue mass, metabolic activity and implications to whole animal energy metabolism. Ann. Zootech. 1995, 44, 321–346. [Google Scholar] [CrossRef]
- McLeod, K.; Baldwin, R. Effects of diet forage: Concentrate ratio and metabolizable energy intake on visceral organ growth and in vitro oxidative capacity of gut tissues in sheep. J. Anim. Sci. 2000, 78, 760–770. [Google Scholar] [CrossRef]
- McBride, B.; Kelly, J. Energy cost of absorption and metabolism in the ruminant gastrointestinal tract and liver: A review. J. Anim. Sci. 1990, 68, 2997–3010. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Rodríguez, J.; Monleón, E.; Sanz, A.; Badiola, J.; Joy, M. Rumen fermentation and histology in light lambs as affected by forage supply and lactation length. Res. Vet. Sci. 2012, 92, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, R.R.; Schnorr, B. Die Funktionelle Morphologie des Wiederkäuer-Magens: Schleimhaut und Versorgungsbahnen; Enke: Stuttgart, Germany, 1982. [Google Scholar]
- Clauss, M.; Hofmann, R.R.; Fickel, J.; Streich, W.J.; Hummel, J. The intraruminal papillation gradient in wild ruminants of different feeding types: Implications for rumen physiology. J. Morphol. 2009, 270, 929–942. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Jiang, Z. Feeding strategies of Przewalski’s antelope. Curr. Zool. 2002, 48, 309–316. [Google Scholar] [CrossRef]
- Beauchemin, K.A. Ingestion and Mastication of Feed by Dairy Cattle. Vet. Clin. N. Am. Food Anim. Pract. 1991, 7, 439–463. [Google Scholar] [CrossRef]
- Domingue, B.F.O.; Dellow, D.W.; Barry, T.N.O. The efficiency of chewing during eating and ruminating in goats and sheep. Br. J. Nutr. 1991, 65, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Van, S.; Peter, J. Nutrition Ecology of the Ruminant; Cornell University Press: Ithaca, NY, USA, 1994. [Google Scholar]
- Liu, Z.; Wang, X.; Li, Z.; Cui, D. Summer feeding and bedding habitat selection by blue sheep (Pseudois nayaur) in Helan Mountains. Acta Ecol. Sin. 2008, 28, 4277–4285. [Google Scholar] [CrossRef]
- Allen, M.S. Physical constraints on voluntary intake of forages by ruminants. J. Anim. Sci. 1996, 74, 3063–3075. [Google Scholar] [CrossRef]
- Li, J.; Jiang, Z. Distribution of different size food particles in alimentary tract of yellow sheep. Acta Zool. Sin. 2001, 47, 488–594. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement Item | Male (n = 30) | Female (n = 11) | Grand Total (n = 41) | ANOVA/ANCOVA |
|---|---|---|---|---|
| Body weight (kg) | 35.3 ± 1.4 | 23.8 ± 2.0 | 32.2 ± 1.4 | F1,38 = 16.745, p < 0.001 |
| Body length (cm) | 91.2 ± 1.4 | 87.6 ± 1.6 | 90.2 ± 1.1 | F1,39 = 1.953, p = 0.170 |
| Ear length (cm) | 10.7 ± 0.1 | 10.6 ± 0.4 | 10.7 ± 0.2 | F1,39 = 0.039, p = 0.845 |
| Tail length (cm) | 10.2 ± 0.3 | 9.9 ± 0.3 | 10.2 ± 0.2 | F1,39 = 0.365, p = 0.549 |
| Horn length (cm) | 41.5 ± 2.1 | 7.2 ± 1.0 | 32.3 ± 2.8 | F1,38 = 88.862, p < 0.001 |
| Hind foot length (cm) | 27.4 ± 0.8 | 26.1 ± 0.7 | 27.1 ± 0.6 | F1,38 = 0.048, p = 0.828 |
| Bust (cm) | 83.5 ± 1.4 | 73.0 ± 2.3 | 80.7 ± 1.4 | F1,38 = 11.629, p = 0.002 |
| Neck circumference (cm) | 37.7 ± 1.4 | 26.6 ± 1.0 | 34.8 ± 1.3 | F1,38 = 18.777, p < 0.001 |
| Horn base circumference (cm) | 22.8 ± 0.9 | 7.5 ± 0.8 | 18.7 ± 1.3 | F1,38 = 97.624, p < 0.001 |
| Shoulder height (cm) | 70.5 ± 1.0 | 64.3 ± 1.0 | 68.8 ± 0.9 | F1,39 = 12.278, p = 0.001 |
| Measurement Item | Unknown Sex | Male | Female | Total | |
|---|---|---|---|---|---|
| Body weight | 32.0 (1) | 36.5 ± 7.4 (20) | 24.3 ± 6.4 (7) | 33.3 ± 8.7 (28) | |
| Stomach | Tissue weight | 0.72 ± 0.17 (17) | 0.74 ± 0.28 (31) | 0.79 ± 0.22 (15) | 0.78 ± 0.03 (61) |
| Including contents | 1.92 ± 0.39 (17) | 4.20 ± 0.39 (31) | 4.31 ± 0.59 (15) | 3.77 ± 0.28 (63) | |
| Rumen and reticulum | Tissue weight | 0.58 ± 0.14 (17) | 0.66 ± 0.18 (29) | 0.64 ± 0.19 (15) | 0.63 ± 0.17 (61) |
| Including contents | 1.53 ± 0.38 (17) | 3.79 ± 0.39 (31) | 3.87 ± 0.58 (15) | 3.19 ± 0.28 (63) | |
| Omasum | Tissue weight | 0.07 ± 0.04 (17) | 0.07 ± 0.03 (32) | 0.08 ± 0.03 (15) | 0.07 ± 0.03 (64) |
| Including contents | 0.24 ± 0.01 (17) | 0.26 ± 0.02 (31) | 0.26 ± 0.03 (15) | 0.26 ± 0.01 (63) | |
| Abomasum | Tissue weight | 0.06 ± 0.02 (17) | 0.07 ± 0.02 (32) | 0.07 ± 0.02 (15) | 0.07 ± 0.02 (64) |
| Including contents | 0.14 ± 0.01 (17) | 0.16 ± 0.01 (31) | 0.18 ± 0.03 (15) | 0.16 ± 0.01 (63) | |
| Stomach | Rumen and Reticulum | Omasum | Abomasum | |||||
|---|---|---|---|---|---|---|---|---|
| Weight of Tissue | Weight Including Contents | Weight of Tissue | Weight Including Contents | Weight of Tissue | Weight Including Contents | Weight of Tissue | Weight Including Contents | |
| Spring | 0.77 ± 0.04 (4) | 2.61 ± 1.18 (4) | 0.68 ± 0.08 (4) | 2.40 ± 1.05 (4) | 0.05 ± 0.02 (4) | 0.13 ± 0.08 (4) | 0.04 ± 0.02 (4) | 0.08 ± 0.04 (4) |
| Summer | 0.51 ± 0.08 (4) | 3.68 ± 1.22 (4) | 0.41 ± 0.07 (4) | 1.78 ± 0.51 (4) | 0.05 ± 0.01 (4) | 0.09 ± 0.01 (4) | 0.06 ± 0.01 (4) | 0.08 ± 0.01 (4) |
| Autumn | 0.73 ± 0.09 (4) | 5.90 ± 2.51 (4) | 0.56 ± 0.08 (4) | 5.49 ± 2.32 (4) | 0.05 ± 0.01 (4) | 0.08 ± 0.01 (4) | 0.11 ± 0.01 (4) | 0.33 ± 0.20 (4) |
| Winter | 0.78 ± 0.03 (17) | 4.98 ± 0.37 (17) | 0.63 ± 0.03 (17) | 4.58 ± 0.32 (17) | 0.06 ± 0.01 (17) | 0.24 ± 0.01 (17) | 0.07 ± 0.01 (17) | 0.15 ± 0.01 (17) |
| ANOVA (F) | F4,29 = 2.66, p = 0.08 | F4,29 = 1.72, p = 0.20 | F4,29 = 2.57, p = 0.08 | F4,29 = 3.06 p = 0.07 | F4,29 = 0.91, p = 0.46 | F4,29 = 9.57, p < 0.001 | F4,29 = 4.30, p = 0.02 | F4,29 = 4.63, p = 0.01 |
| Organ | Papillae Number (/2 cm2) | Papillae Height (cm) | Papillae Width (cm) | Surface Enlargement Factor |
|---|---|---|---|---|
| Atrium ruminis | 55 ± 19 a | 0.263 ± 0.093 b | 0.105 ± 0.035 b | 3.90 ± 1.58 b |
| Dorsal rumen | 78 ± 26 b | 0.162 ± 0.070 a | 0.075 ± 0.028 a | 2.35 ± 0.96 a |
| Ventral rumen | 76 ± 25 b | 0.166 ± 0.063 a | 0.085 ± 0.027 a | 2.53 ± 0.94 a |
| Dorsal rumen blindsacs | 66 ± 25 ab | 0.179 ± 0.073 a | 0.081 ± 0.037 a | 2.62 ± 1.33 a |
| Length | Weight | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Small Intestine | Cecum | Colon Crassum | Microcolon | Rectum | Small Intestine | Cecum | Colon Crassum | Microcolon | Rectum | ||
| Sex | Unknown sex | 1671.4 ± 42.4 | 34.5 ± 1.9 | 79.2 ± 6.6 | 366.5 ± 19.7 | 129.4 ± 8.6 | 1714.9 ± 22.8 | 1684.7 ± 42.5 | 1669.9 ± 19.9 | 1714.9 ± 22.8 | 1684.7 ± 42.5 |
| Male | 1714.9 ± 22.8 | 33.3 ± 1.2 | 61.2 ± 2.1 | 397.6 ± 12.6 | 137.1 ± 4.8 | 33.3 ± 1.2 | 33.2 ± 1.2 | 33.6± 0.7 | 33.3 ± 1.2 | 33.2 ± 1.2 | |
| Female | 1684.7 ± 42.5 | 33.2 ± 1.2 | 70.9 ± 9.2 | 380.1 ± 14.1 | 124.1 ± 8.7 | 61.2 ± 2.1 | 70.9 ± 9.2 | 67.9 ± 3.0 | 61.2 ± 2.1 | 70.9 ± 9.2 | |
| Total | 1669.9 ± 19.9 | 33.6 ± 0.7 | 67.9 ± 3.0 | 384.9 ± 8.7 | 132.1 ± 3.8 | 397.6 ± 12.6 | 380.1 ± 14.1 | 384.9 ± 8.7 | 397.6 ± 12.6 | 380.1 ± 14.1 | |
| Season | Winter | 1671.4 ± 21.1 | 33.9 ± 0.7 | 68.0 ± 3.2 | 386.4 ± 9.7 | 135.1 ± 3.6 | 0.51 ± 0.02 | 0.18 ± 0.02 | 0.21 ± 0.02 | 0.28 ± 0.01 | 0.24 ± 0.01 |
| Summer | 1642.4 ± 56.3 | 28.5 ± 2.8 | 66.0 ± 8.4 | 368.3 ± 32.7 | 97.2 ± 17.3 | 0.31 ± 0.06 | 0.12 ± 0.04 | 0.16 ± 0.04 | 0.15 ± 0.02 | 0.14 ± 0.03 | |
| ANOVA | F1,61 = 0.153, p = 0.697 | F1,61 = 0.153, p = 0.697 | F1,61 = 0.153, p = 0.697 | F1,61 = 0.153, p = 0.697 | F1,61 = 0.153, p = 0.697 | F1,61 = 0.153, p = 0.697 | F1,61 = 0.153, p = 0.697 | F1,61 = 0.153, p = 0.697 | F1,61 = 0.153, p = 0.697 | F1,61 = 0.153, p = 0.697 | |
| Organ | Season | Particle Size Distribution of Different Grades (%) | ||||
|---|---|---|---|---|---|---|
| 0.125 < x < 0.25 | 0.25 < x < 0.425 | 0.425 < x < 1.0 | 1.0 < x < 2.0 | x > 2.0 | ||
| Rumen and Reticulum | Summer | 5.75 ± 1.00 | 3.42 ± 0.80 | 8.93 ± 0.83 | 4.20 ± 1.31 | 77.69 ± 2.02 |
| Winter | 4.17 ± 1.07 | 2.71 ± 0.96 | 9.80 ± 1.80 | 17.97 ± 6.47 | 65.35 ± 7.17 | |
| ANOVA | F = 0.33, p = 0.57 | F = 0.08, p = 0.78 | F = 0.04, p = 0.85 | F = 6.69, p = 0.02 | F = 4.44, p = 0.04 | |
| Omasum | Summer | 16.05 ± 2.52 | 12.98 ± 3.89 | 25.46 ± 6.14 | 16.71 ± 7.32 | 28.80 ± 16.82 |
| Winter | 6.44 ± 1.45 | 10.06 ± 2.44 | 26.99 ± 4.42 | 25.17 ± 3.78 | 31.34 ± 6.35 | |
| ANOVA | F = 6.34, p = 0.02 | F = 0.21, p = 0.65 | F = 0.02, p = 0.90 | F = 7.02, p = 0.01 | F = 0.02, p = 0.89 | |
| Abomasum | Summer | 12.55 ± 6.60 | 14.94 ± 4.68 | 18.76 ± 1.27 | 26.71 ± 5.64 | 27.04 ± 6.68 |
| Winter | 16.76 ± 2.65 | 11.60 ± 1.96 | 29.57 ± 3.43 | 19.76 ± 3.75 | 22.32 ± 3.89 | |
| ANOVA | F = 0.34, p = 0.56 | F = 0.40, p = 0.53 | F = 7.51, p = 0.02 | F = 0.05, p = 0.49 | F = 0.21, p = 0.65 | |
| Colon crassum | Summer | 17.33 ± 1.34 | 14.88 ± 5.37 | 31.66 ± 8.17 | 13.25 ± 5.68 | 22.88 ± 8.12 |
| Winter | 15.85 ± 2.49 | 12.50 ± 1.60 | 37.28 ± 3.40 | 18.29 ± 3.21 | 16.09 ± 3.77 | |
| ANOVA | F = 0.05, p = 0.82 | F = 0.28, p = 0.60 | F = 0.38, p = 0.55 | F = 0.35, p = 0.56 | F = 0.46, p = 0.51 | |
| Small intestine | Summer | 18.56 ± 4.39 | 9.71 ± 2.26 | 19.89 ± 6.71 | 22.07 ± 4.02 | 29.77 ± 8.24 |
| Winter | 11.63 ± 2.52 | 11.25 ± 1.41 | 38.74 ± 3.23 | 24.49 ± 2.07 | 13.89 ± 3.03 | |
| ANOVA | F = 1.09, p = 0.31 | F = 0.17, p = 0.68 | F = 4.80, p = 0.04 | F = 0.20, p = 0.66 | F = 3.95, p = 0.04 | |
| Cecum | Summer | 16.06 ± 2.85 | 12.96 ± 2.75 | 26.22 ± 5.35 | 23.15 ± 4.89 | 21.61 ± 6.28 |
| Winter | 18.12 ± 2.21 | 14.14 ± 1.53 | 35.93 ± 2.54 | 20.85 ± 3.12 | 9.96 ± 1.98 | |
| ANOVA | F = 0.13, p = 0.72 | F = 0.29, p = 0.60 | F = 9.05, p = 0.01 | F = 0.08, p = 0.78 | F = 4.47, p = 0.03 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, D.; Si, Y.; Wang, J.; Li, Z.; Khattak, R.H.; Liu, Z.; Teng, L. The Effect of Sexes and Seasons on the Morphological Structures of the Ruminant Digestive System of Blue Sheep (Pseudois nayaur). Animals 2023, 13, 1084. https://doi.org/10.3390/ani13061084
Meng D, Si Y, Wang J, Li Z, Khattak RH, Liu Z, Teng L. The Effect of Sexes and Seasons on the Morphological Structures of the Ruminant Digestive System of Blue Sheep (Pseudois nayaur). Animals. 2023; 13(6):1084. https://doi.org/10.3390/ani13061084
Chicago/Turabian StyleMeng, Dehuai, Yuhui Si, Jifei Wang, Zongzhi Li, Romaan Hayat Khattak, Zhensheng Liu, and Liwei Teng. 2023. "The Effect of Sexes and Seasons on the Morphological Structures of the Ruminant Digestive System of Blue Sheep (Pseudois nayaur)" Animals 13, no. 6: 1084. https://doi.org/10.3390/ani13061084
APA StyleMeng, D., Si, Y., Wang, J., Li, Z., Khattak, R. H., Liu, Z., & Teng, L. (2023). The Effect of Sexes and Seasons on the Morphological Structures of the Ruminant Digestive System of Blue Sheep (Pseudois nayaur). Animals, 13(6), 1084. https://doi.org/10.3390/ani13061084