
Serotonin and Dopamine Blood Levels in ADHD-Like Dogs


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Surveyed Animals and Data Collection
2.2. Dog Assessment
2.3. Serotonin and Dopamine Measurement
2.4. Statistical Analyses
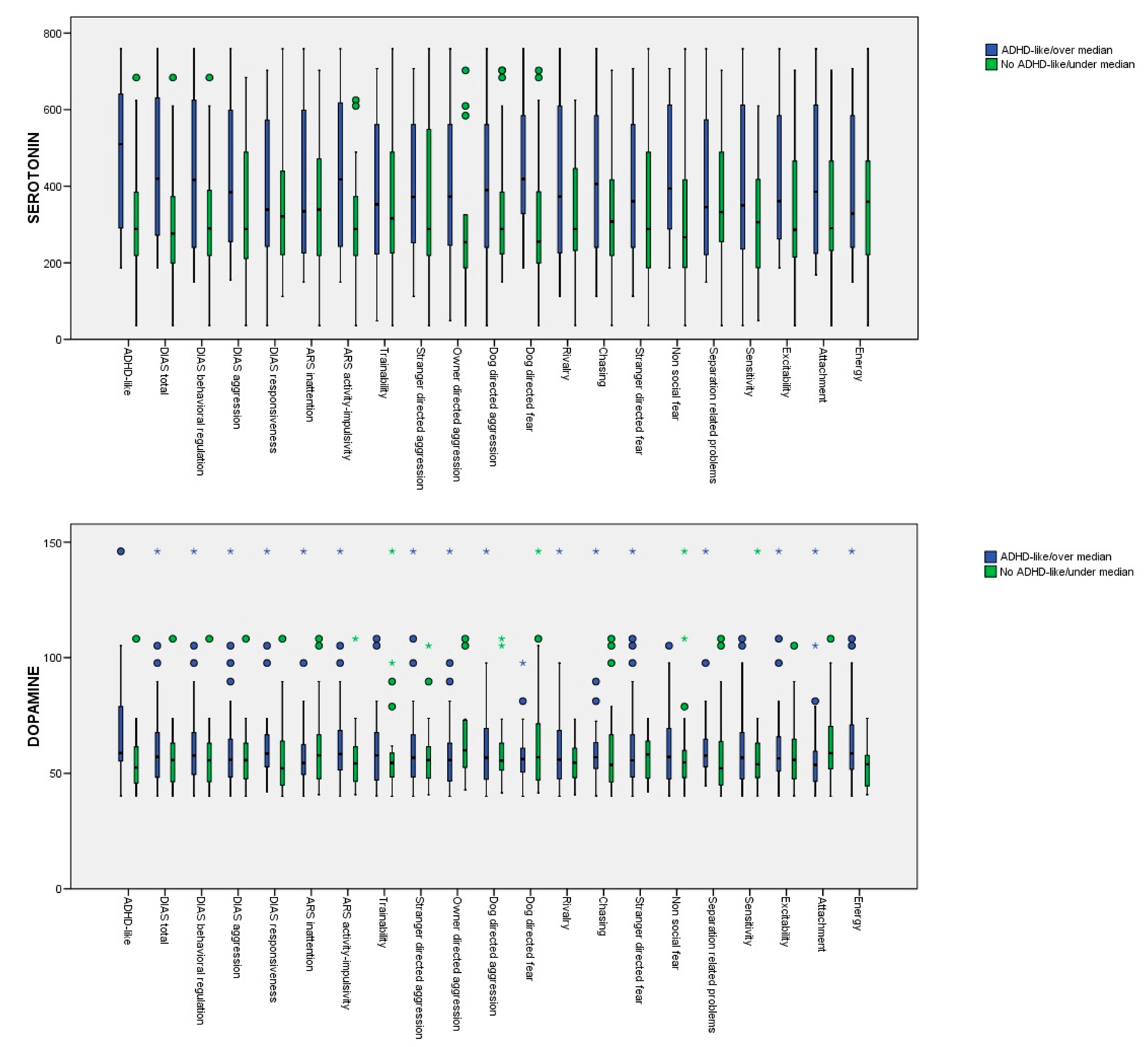
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
| Serum Serotonin Concentration (95% CI) | Serum Dopamine Concentration (95% CI) | |
|---|---|---|
| ADHD-like | 0.03 | 0.016 |
| DIAS total score | 0.005 | 0423 |
| DIAS behavioral regulation score | 0.01 | 0.233 |
| DIAS responsiveness score | 0.523 | 0.119 |
| DIAS aggression score | 0.002 | 0.970 |
| Dog ARS activity-impulsivity score | 0.012 | 0.011 |
| Dog ARS inattention score | 0.435 | 0.563 |
| C-BARQ trainability | 0.869 | 0.443 |
| C-BARQ stranger-directed aggression | 0.316 | 0.538 |
| C-BARQ owner-directed aggression | 0.043 | 0.154 |
| C-BARQ dog-directed aggression | 0.272 | 0.836 |
| C-BARQ dog-directed fear | 0.004 | 0.593 |
| C-BARQ rivalry | 0.176 | 0.676 |
| C-BARQ chasing | 0.212 | 0.469 |
| C-BARQ stranger-directed fear | 0.257 | 0.927 |
| C-BARQ nonsocial fear | 0.023 | 0.428 |
| C-BARG separation-related problems | 0.961 | 0.157 |
| C-BARQ sensitivity | 0.132 | 0.686 |
| C-BARQ excitability | 0.104 | 0.599 |
| C-BARQ attachment | 0.140 | 0.142 |
| C-BARQ energy | 0.560 | 0.040 |
References
- Paclt, I.; Koudelová, J.; Křepelová, A.; Uhlíková, P.; Gazdíková, M.; Bauer, P. Biochemical Markers and Genetic Research of ADHD. Neuroendocrinol. Lett. 2005, 26, 423–430. [Google Scholar] [PubMed]
- Morrison, J. DSM-5® Guía para el Diagnóstico Clínico; Editorial El Manual Moderno: Mexico City, Mexico, 2015. [Google Scholar]
- Faraone, S.V.; Biederman, J.; Spencer, T.J.; Aleardi, M. Comparing the Efficacy of Medications for ADHD Using Meta-Analysis. Medscape Gen. Med. 2006, 8, 4. [Google Scholar]
- Hoogman, M.; Bralten, J.; Hibar, D.P.; Mennes, M.; Zwiers, M.P.; Schweren, L.S.J.; van Hulzen, K.J.E.; Medland, S.E.; Shumskaya, E.; Jahanshad, N.; et al. Subcortical Brain Volume Differences in Participants with Attention Deficit Hyperactivity Disorder in Children and Adults: A Cross-Sectional Mega-Analysis. Lancet Psychiatry 2017, 4, 310–319. [Google Scholar] [CrossRef]
- Xavier Castellanos, F.; Lee, P.P.; Sharp, W.; Neal Jeffries, M.O.; Greenstein, D.K.; Clasen, L.S.; Blumenthal, J.D.; Regina James, M.S.; Ebens, C.L.; James Walter, B.M.; et al. Developmental Trajectories of Brain Volume Abnormalities in Children and Adolescents with Attention-Deficit/Hyperactivity Disorder. JAMA 2002, 288, 1740–1748. [Google Scholar] [CrossRef]
- Hoogman, M.; Rijpkema, M.; Janss, L.; Brunner, H.; Fernandez, G.; Buitelaar, J.; Franke, B.; Arias-Vásquez, A. Current Self-Reported Symptoms of Attention Deficit/Hyperactivity Disorder Are Associated with Total Brain Volume in Healthy Adults. PLoS ONE 2012, 7, e31273. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.; Gilliam, M.; Liverpool, M.; Weddle, C.; Malek, M.; Sharp, W.; Greenstein, D.; Evans, A.; Rapoport, J.; Giedd, J. Cortical Development in Typically Developing Children with Symptoms of Hyperactivity and Impulsivity: Support for a Dimensional View of Attention Deficit Hyperactivity Disorder. Am. J. Psychiatry 2011, 168, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Vas, J.; Topál, J.; Péch, É.; Miklósi, Á. Measuring Attention Deficit and Activity in Dogs: A New Application and Validation of a Human ADHD Questionnaire. Appl. Anim. Behav. Sci. 2007, 103, 105–117. [Google Scholar] [CrossRef]
- Lit, L.; Schweitzer, J.B.; Iosif, A.M.; Oberbauer, A.M. Owner Reports of Attention, Activity, and Impulsivity in Dogs: A Replication Study. Behav. Brain Funct. 2010, 6, 1. [Google Scholar] [CrossRef]
- Piotti, P.; Satchell, L.P.; Lockhart, T.S. Impulsivity and Behaviour Problems in Dogs: A Reinforcement Sensitivity Theory Perspective. Behav. Process. 2018, 151, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Sonuga-Barke, E.J.S. Causal Models of Attention-Deficit/Hyperactivity Disorder: From Common Simple Deficits to Multiple Developmental Pathways. Biol. Psychiatry 2005, 57, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, N.; Chamberlain, S.R.; Sahakian, B.J.; Robbins, T.W. The Roles of Dopamine and Noradrenaline in the Pathophysiology and Treatment of Attention-Deficit/Hyperactivity Disorder. Biol. Psychiatry 2011, 69, e145–e157. [Google Scholar] [CrossRef]
- Cho, H.S.; Baek, D.J.; Baek, S.S. Effect of Exercise on Hyperactivity, Impulsivity and Dopamine D2 Receptor Expression in the Substantia Nigra and Striatum of Spontaneous Hypertensive Rats. J. Exerc. Nutr. Biochem. 2014, 18, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Mogavero, F.; Jager, A.; Glennon, J.C. Clock Genes, ADHD and Aggression. Neurosci. Biobehav. Rev. 2018, 91, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Mü ller Smith, K.; Daly, M.; Fischer, M.; Yiannoutsos, C.T.; Bauer, L.; Barkley, R.; Navia, B.A. Association of the Dopamine Beta Hydroxylase Gene with Attention Deficit Hyperactivity Disorder: Genetic Analysis of the Milwaukee Longitudinal Study. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2003, 119, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Luescher, U.A. Hyperkinesis in Dogs: Six Case Reports. Can. Vet. J. 1993, 34, 368–370. [Google Scholar] [PubMed]
- Piturru, P. Methylphenidate Use in Dogs with Attention Deficit Hyperactivity Disorder (ADHD). A Case Report of a Weimaraner Bitch. Tierarztl. Prax. Ausgabe K Kleintiere-Heimtiere 2014, 42, 111–116. [Google Scholar] [CrossRef]
- Lowe, N.; Kirley, A.; Hawi, Z.; Sham, P.; Wickham, H.; Kratochvil, C.J.; Smith, S.D.; Lee, S.Y.; Levy, F.; Kent, L.; et al. Joint Analysis of the DRD5 Marker Concludes Association with Attention-Deficit/Hyperactivity Disorder Confined to the Predominantly Inattentive and Combined Subtypes. Am. J. Hum. Genet. 2004, 74, 348–356. [Google Scholar] [CrossRef]
- Kubinyi, E.; Vas, J.; Hejjas, K.; Ronai, Z.; Brúder, I.; Turcsán, B.; Sasvari-Szekely, M.; Miklósi, Á. Polymorphism in the Tyrosine Hydroxylase (TH) Gene Is Associated with Activity-Impulsivity in German Shepherd Dogs. PLoS ONE 2012, 7, e30271. [Google Scholar] [CrossRef]
- Hejjas, K.; Vas, J.; Topal, J.; Szantai, E.; Ronai, Z.; Szekely, A.; Kubinyi, E.; Horvath, Z.; Sasvari-Szekely, M.; Miklosi, A. Association of Polymorphisms in the Dopamine D4 Receptor Gene and the Activity-Impulsivity Endophenotype in Dogs. Anim. Genet. 2007, 38, 629–633. [Google Scholar] [CrossRef]
- Bolaños, C.A.; Willey, M.D.; Maffeo, M.L.; Powers, K.D.; Kinka, D.W.; Grausam, K.B.; Henderson, R.P. Antidepressant Treatment Can Normalize Adult Behavioral Deficits Induced by Early-Life Exposure to Methylphenidate. Biol. Psychiatry 2008, 63, 309–316. [Google Scholar] [CrossRef]
- Neil, R.; Carlson, B. Physiology of Behavior, 13th ed.; Pearson Education: Essex, UK, 2022. [Google Scholar]
- Quist, J.F.; Barr, C.L.; Schachar, R.; Roberts, W.; Malone, M.; Tannock, R.; Basile, V.S.; Beitchman, J.; Kennedy, J.L. The Serotonin 5-HT1B Receptor Gene and Attention Deficit Hyperactivity Disorder. Mol. Psychiatry 2003, 8, 98–102. [Google Scholar] [CrossRef]
- Banerjee, E.; Nandagopal, K. Does Serotonin Deficit Mediate Susceptibility to ADHD? Neurochem. Int. 2015, 82, 52–68. [Google Scholar] [CrossRef]
- Bleuer-Elsner, S.; Muller, G.; Beata, C.; Zamansky, A.; Marlois, N. Effect of Fluoxetine at a Dosage of 2-4 Mg/Kg Daily in Dogs Exhibiting Hypersensitivity-Hyperactivity Syndrome, a Retrospective Study. J. Vet. Behav. 2021, 44, 25–31. [Google Scholar] [CrossRef]
- Masson, S.; Gaultier, E. Retrospective Study on Hypersensitivity-Hyperactivity Syndrome in Dogs: Long-Term Outcome of High Dose Fluoxetine Treatment and Proposal of a Clinical Score. Dog Behav. 2018, 4, 15–32. [Google Scholar] [CrossRef]
- Lunõ, I.; Rosado, B.; Palacio, J.; Villegas, A.; González-Martínez, Á.; Garciá-Belenguer, S. Hyperactivity in a Weimaraner Dog. Dog Behav. 2015, 1, 32–40. [Google Scholar] [CrossRef]
- Barkley, R.A. Behavioral Inhibition, Sustained Attention, and Executive Functions: Constructing a Unifying Theory of ADHD. Psychol. Bull. 1997, 121, 65–94. [Google Scholar] [CrossRef] [PubMed]
- Coleman, M. Serotonin Concentrations in Whole Blood of Hyperactive Children. J. Pediatr. 1971, 78, 985–990. [Google Scholar] [CrossRef]
- Spivak, B.; Vered, Y.; Yoran-Hegesh, R.; Averbuch, E.; Mester, R.; Graf, E.; Weizman, A. Circulatory Levels of Catecholamines, Serotonin and Lipids in Attention Deficit Hyperactivity Disorder. Acta Psychiatr. Scand. 1999, 99, 300–304. [Google Scholar] [CrossRef]
- Lombroso, P.J.; Quist, J.F.; Kennedy, J.L. Genetics of Childhood Disorders: XXIII. ADHD, Part 7: The Serotonin System. J. Am. Acad. Child Adolesc. Psychiatry 2001, 40, 253–256. [Google Scholar]
- Wright, H.F.; Mills, D.S.; Pollux, P.M.J. Behavioural and Physiological Correlates of Impulsivity in the Domestic Dog (Canis familiaris). Physiol. Behav. 2012, 105, 676–682. [Google Scholar] [CrossRef]
- Rosado, B.; García-Belenguer, S.; León, M.; Chacón, G.; Villegas, A.; Palacio, J. Blood Concentrations of Serotonin, Cortisol and Dehydroepiandrosterone in Aggressive Dogs. Appl. Anim. Behav. Sci. 2010, 123, 124–130. [Google Scholar] [CrossRef]
- León, M.; Rosado, B.; García-Belenguer, S.; Chacón, G.; Villegas, A.; Palacio, J. Assessment of Serotonin in Serum, Plasma, and Platelets of Aggressive Dogs. J. Vet. Behav. Clin. Appl. Res. 2012, 7, 348–352. [Google Scholar] [CrossRef]
- Rosado, B.; García-Belenguer, S.; Palacio, J.; Chacón, G.; Villegas, A.; Alcalde, A.I. Serotonin Transporter Activity in Platelets and Canine Aggression. Vet. J. 2010, 186, 104–105. [Google Scholar] [CrossRef]
- Amat, M.; Le Brech, S.; Camps, T.; Torrente, C.; Mariotti, V.M.; Ruiz, J.L.; Manteca, X. Differences in Serotonin Serum Concentration between Aggressive English Cocker Spaniels and Aggressive Dogs of Other Breeds. J. Vet. Behav. Clin. Appl. Res. 2013, 8, 19–25. [Google Scholar] [CrossRef]
- Hoppe, N.; Bininda-Emonds, O.R.P.; Gansloßer, U. Correlates of Attention Deficit Hyperactivity Disorder (ADHD)-Like Behavior in Domestic Dogs: First Results from a Questionnaire-Based Study. Vet. Med.–Open J. 2017, 2, 95–131. [Google Scholar] [CrossRef]
- Serpell, J.A.; Hsu, Y. Development and Validation of a Novel Method for Evaluating Behavior and Temperament in Guide Dogs. Appl. Anim. Behav. Sci. 2001, 72, 347–364. [Google Scholar] [CrossRef]
- Duffy, D.L.; Hsu, Y.; Serpell, J.A. Breed Differences in Canine Aggression. Appl. Anim. Behav. Sci. 2008, 114, 441–460. [Google Scholar] [CrossRef]
- Serpell, J.A.; Hsu, Y. Effects of Breed, Sex, and Neuter Status on Trainability in Dogs. Anthrozoös 2005, 18, 196–207. [Google Scholar] [CrossRef]
- Wright, H.F.; Mills, D.S.; Pollux, P.M.J. Development and Validation of a Psychometric Tool for Assessing Impulsivity in the Domestic Dog (Canis familiaris). Int. J. Comp. Psychol. 2011, 24, 210–225. [Google Scholar] [CrossRef]
- Riemer, S.; Mills, D.S.; Wright, H. Impulsive for Life? The Nature of Long-Term Impulsivity in Domestic Dogs. Anim. Cogn. 2014, 17, 815–819. [Google Scholar] [CrossRef]
- Crockett, M.J.; Clark, L.; Robbins, T.W. Reconciling the Role of Serotonin in Behavioral Inhibition and Aversion: Acute Tryptophan Depletion Abolishes Punishment-Induced Inhibition in Humans. J. Neurosci. 2009, 29, 11993–11999. [Google Scholar] [CrossRef]
- Baumann, M.H.; Williams, Z.; Zolkowska, D.; Rothman, R.B. Serotonin (5-HT) Precursor Loading with 5-Hydroxy-l-Tryptophan (5-HTP) Reduces Locomotor Activation Produced by (+)-Amphetamine in the Rat. Drug Alcohol Depend. 2011, 114, 147–152. [Google Scholar] [CrossRef]
- Kabuki, Y.; Yamane, H.; Hamasu, K.; Furuse, M. Different Locomotor Activities and Monoamine Levels in the Brains of Djungarian Hamsters (D. sungorus) and Roborovskii Hamsters (D. roborovskii). Exp. Anim. 2008, 57, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Takada, Y.; Urano, T.; Takada, A. 5-HT4 Receptors in the Hippocampus Modulate Rat Locomotor Activity. Hippocampus 2002, 12, 304–310. [Google Scholar] [CrossRef]
- Hechtman, L. Developmental, Neurobiological, and Psychosocial Aspects of Hyperactivity, Impulsivity, and Inattention. Child Adolesc. Psychiatry 1991, 318–330. [Google Scholar]
- Peremans, K.; Audenaert, K.; Coopman, F.; Blanckaert, P.; Jacobs, F.; Otte, A.; Verschooten, F.; Van Bree, H.; Van Heeringen, K.; Mertens, J.; et al. Estimates of Regional Cerebral Blood Flow and 5-HT2A Receptor Density in Impulsive, Aggressive Dogs with 99mTc-ECD and 123I-5-I-R91150. Eur. J. Nucl. Med. Mol. Imaging 2003, 30, 1538–1546. [Google Scholar] [CrossRef] [PubMed]
- Biederman, J.; Faraone, S.V.; Spencer, T.; Wilens, T.; Norman, D.; Lapey, K.A.; Mick, E.; Lehman, B.K.; Doyle, A. Patterns of Psychiatric Comorbidity, Cognition, and Psychosocial Functioning in Adults with Attention Deficit Hyperactivity Disorder. Am. J. Psychiatry 1993, 150, 1792–1798. [Google Scholar] [CrossRef] [PubMed]
- Sobanski, E. Psychiatric Comorbidity in Adults with Attention-Deficit/Hyperactivity Disorder (ADHD). Eur. Arch. Psychiatry Clin. Neurosci. 2006, 256, i26–i31. [Google Scholar] [CrossRef]
- Rucklidge, J.J.; Downs-Woolley, M.; Taylor, M.; Brown, J.A.; Harrow, S.E. Psychiatric Comorbidities in a New Zealand Sample of Adults with ADHD. J. Atten. Disord. 2016, 20, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Mikkola, S.; Salonen, M.; Puurunen, J.; Hakanen, E.; Sulkama, S.; Araujo, C.; Lohi, H. Aggressive Behaviour Is Affected by Demographic, Environmental and Behavioural Factors in Purebred Dogs. Sci. Rep. 2021, 11, 9433. [Google Scholar] [CrossRef]
- Deakin, J.F.W.; Graeff, F.G. 5-HT and Mechanisms of Defence. Author’s Response. J. Psychopharmacol. 1991, 5, 339–341. [Google Scholar] [CrossRef] [PubMed]
- Deakin, J.F.W. The Origins of “5-HT and Mechanisms of Defence” by Deakin and Graeff: A Personal Perspective. J. Psychopharmacol. 2013, 27, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Corchs, F.; Nutt, D.J.; Hince, D.A.; Davies, S.J.C.; Bernik, M.; Hood, S.D. Evidence for Serotonin Function as a Neurochemical Difference between Fear and Anxiety Disorders in Humans? J. Psychopharmacol. 2015, 29, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Van Ameringen, M.; Mancini, C.; Oakman, J.M.; Farvolden, P. Selective Serotonin Reuptake Inhibitors in the Treatment of Social Phobia: The Emerging Gold Standard. CNS Drugs 1999, 11, 307–315. [Google Scholar] [CrossRef]
- Den Boer, J.A.; Bosker, F.J.; Slaap, B.R. Serotonergic Drugs in the Treatment of Depressive and Anxiety Disorders. Hum. Psychopharmacol. 2000, 15, 315–336. [Google Scholar] [CrossRef]
- Seksel, K.; Lindeman, M.J. Use of Clomipramine in Treatment of Obsessive-Compulsive Disorder, Separation Anxiety and Noise Phobia in Dogs: A Preliminary, Clinical Study. Aust. Vet. J. 2001, 79, 252–256. [Google Scholar] [CrossRef]
- Ibáñez, M.; Anzola, B. Use of Fluoxetine, Diazepam, and Behavior Modification as Therapy for Treatment of Anxiety-Related Disorders in Dogs. J. Vet. Behav. Clin. Appl. Res. 2009, 4, 223–229. [Google Scholar] [CrossRef]
- Shannon, C.; Schwandt, M.L.; Champoux, M.; Shoaf, S.E.; Suomi, S.J.; Linnoila, M.; Higley, J.D. Maternal Absence and Stability of Individual Differences in CSF 5-HIAA Concentrations in Rhesus Monkey Infants. Am. J. Psychiatry 2005, 162, 1658–1664. [Google Scholar] [CrossRef]
- Kiser, D.; SteemerS, B.; Branchi, I.; Homberg, J.R. The Reciprocal Interaction between Serotonin and Social Behaviour. Neurosci. Biobehav. Rev. 2012, 36, 786–798. [Google Scholar] [CrossRef]
- Kinnally, E.L.; Capitanio, J.P.; Leibel, R.; Deng, L.; Leduc, C.; Haghighi, F.; Mann, J.J. Epigenetic Regulation of Serotonin Transporter Expression and Behavior in Infant Rhesus Macaques. Genes Brain Behav. 2010, 9, 575–582. [Google Scholar] [CrossRef]
- Hejjas, K.; Vas, J.; Kubinyi, E.; Sasvari-Szekely, M.; Miklosi, A.; Ronai, Z. Novel Repeat Polymorphisms of the Dopaminergic Neurotransmitter Genes among Dogs and Wolves. Mamm. Genome 2007, 18, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Hejjas, K.; Kubinyi, E.; Ronai, Z.; Szekely, A.; Vas, J.; Miklósi, Á.; Sasvari-Szekely, M.; Kereszturi, E. Molecular and Behavioral Analysis of the Intron 2 Repeat Polymorphism in the Canine Dopamine D4 Receptor Gene. Genes Brain Behav. 2009, 8, 330–336. [Google Scholar] [CrossRef]
- Sulkama, S.; Puurunen, J.; Salonen, M.; Mikkola, S.; Hakanen, E.; Araujo, C.; Lohi, H. Canine Hyperactivity, Impulsivity, and Inattention Share Similar Demographic Risk Factors and Behavioural Comorbidities with Human ADHD. Transl. Psychiatry 2021, 11, 501. [Google Scholar] [CrossRef]
- Zink, M.C.; Farhoody, P.; Elser, S.E.; Ruffini, L.D.; Gibbons, T.A.; Rieger, R.H. Evaluation of the Risk and Age of Onset of Cancer and Behavioral Disorders in Gonadectomized Vizslas. J. Am. Vet. Med. Assoc. 2014, 244, 309–319. [Google Scholar] [CrossRef]
- Fadel, F.R.; Driscoll, P.; Pilot, M.; Wright, H.; Zulch, H.; Mills, D. Differences in Trait Impulsivity Indicate Diversification of Dog Breeds into Working and Show Lines. Sci. Rep. 2016, 6, 22162. [Google Scholar] [CrossRef]
- Jagoe, A.; Serpell, J. Owner Characteristics and Interactions and the Prevalence of Canine Behaviour Problems. Appl. Anim. Behav. Sci. 1996, 47, 31–42. [Google Scholar] [CrossRef]
- Serpell, J.A.; Duffy, D.L. Aspects of Juvenile and Adolescent Environment Predict Aggression and Fear in 12-Month-Old Guide Dogs. Front. Vet. Sci. 2016, 3, 49. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Langford, F.; Kiddie, J. Risk Factors for Aggressive Behaviour in Domestic Dogs (Canis familiaris), as Reported by Owners in Mainland China. Appl. Anim. Behav. Sci. 2021, 234, 105211. [Google Scholar] [CrossRef]
- Wipfli, B.M. Serotonin and Psychological Variables in the Relationship between Exercise and Mental Health; Arizona State University: Tempe, Arizona, 2008; Volume 69. [Google Scholar]
- Tiira, K.; Lohi, H. Early Life Experiences and Exercise Associate with Canine Anxieties. PLoS ONE 2015, 10, e0141907. [Google Scholar] [CrossRef] [PubMed]

| Attention Deficit Symptoms |
|---|
| Other things attract attention easily |
| Loses interest easily |
| Difficulty concentrating |
| Does not pay attention to someone speaking to him/her |
| Difficulty performing practiced tasks |
| Easily distracted |
| Hyperactivity-impulsivity |
| Difficulty maintaining stay |
| Barks endlessly |
| Fidgets or in constant motion |
| Excessive active play/running around |
| Reacts hastily/anticipates |
| Cannot wait |
| Noninhibited bite |
| Lack of safety |
| Lack of sleep during the day and frequent waking up and movement during the night |
| Aggressive displays |
| Variable | Descriptive Features | ||
|---|---|---|---|
| Total (n = 58) | ADHD-Like (n = 36) | Not ADHD-Like (n = 22) | |
| Gender | Data | Data | |
| Female | 29 (50%) | 16 (44.4%) | 13 (59.1%) |
| Male | 29 (50%) | 20 (55.6%) | 9 (40.9%) |
| Neutered | |||
| Yes | 42 (72.4%) | 28 (77.8%) | 14 (36.4%) |
| No | 16 (17.6%) | 8 (22.2%) | 8 (63.6%) |
| Age (mean, median, (S.D.), and range) | 48.2, 45, (33.9), and 7–156 | 40.2, 34, (31.1), and 11–121 | 61.2, 61, (35.0), and 7–156 |
| Age at acquisition (mean, median, and (S.D.)) | 6.1, 2, and (11.1) | 6.2, 2, and (13.3) | 5.9, 2, and (6.3) |
| Weight (mean, median, and (S.D.)) | 21.4, 22, and (9.4) | 21.3, 21, and (8.4) | 21.7, 23, and (11.1) |
| Dog activity patterns | |||
| No activity | 54 (93.1%) | 34 (94.4%) | 20 (90.9%) |
| Agility/biking/marching/mushing/other | 4 (6.9%) | 2 (5.6%) | 2 (9.1%) |
| Dogs previously owned | |||
| None | 19 (32.8%) | 12 (33.3%) | 7 (31.8%) |
| One or more | 39 (67.2%) | 24 (66.7%) | 15 (68.2%) |
| Habitat | |||
| Rural | 19 (32.8%) | 11 (30.6%) | 9 (40.9%) |
| Urban | 39 (67.2%) | 25 (69.4%) | 13 (59.1%) |
| Serum Serotonin Concentration (95% CI) | Serum Dopamine Concentration (95% CI) | Mean Score (SD) | |
|---|---|---|---|
| ADHD-like | |||
| Yes | 311.91 (257.36–366.45) a | 55.49 (50.51–60.47) a | |
| No | 477.06 (391.20–562.92) b | 68.68 (57,54–70,81) b | |
| DIAS total score | |||
| Under median | 449.28 (376.98–521.59) a | 63.90 (54.66–73.14) | 0.46 (0.07) |
| Over median | 304.29 (240.75–367.83) b | 57.41 (51.49–63.39) | 0.73 (0.08) |
| DIAS behavioral regulation score | |||
| Under median | 428.56 (353.75–503.37) a | 63.99 (55.14–72.85) | 0.37 (0.10) |
| Over median | 321.18 (255.63–386.73) b | 57.04 (50.87–63.22) | 0.75 (0.11) |
| DIAS responsiveness score | |||
| Under median | 389.56 (315.02–464.10) | 64.32 (55.46–73.18) | 0.32 (0.08) |
| Over median | 364.01 (291.23–436.80) | 57.25 (50.57–63.92) | 0.65 (0.20) |
| DIAS aggression score | |||
| Under median | 414.08 (342.37–485.78) a | 63.16 (53.60–72.71) | 0.58 (0.11) |
| Over median | 334.82 (259.17–410.47) b | 57.57 (51.66–63.49) | 0.81 (0.08) |
| Dog ARS activity-impulsivity score | |||
| Under median | 432.10 (360.58–503.62) a | 64.14 (55.52–72.77) a | 2.90 (1.70) |
| Over median | 303.04 (240.17–365.91) b | 56.57 (50.38–62.77) b | 10.1 (3.61) |
| Dog ARS inattention score | |||
| Under median | 402.22 (328.84–475.59) | 60.28 (52.01–68.56) | 4.16 (3.14) |
| Over median | 347.45 (275.34–419.55) | 61.20 (53.76–68.64) | 14.77 (4.15) |
| C-BARQ trainability | |||
| Under median | 381.53 (310.88–452.18) | 60.48 (54.46–66.51) | 2.15 (0.36) |
| Over median | 371.32 (293.90–448.74) | 61.04 (50.44–71.64) | 3.13 (0.30) |
| C-BARQ stranger-directed aggression | |||
| Under median | 394.32 (331.29–457.34) | 62.75 (54.16–71.35 | 0.04 (0.07) |
| Over median | 356.56 (271.37–441.75) | 58.25 (51.71–64.79) | 0.98 (0.81) |
| C-BARQ owner-directed aggression | |||
| Under median | 401.08 (344.04–458.13) a | 58.98 (53.04–64.91) | 0.02 (0.07) |
| Over median | 303.90 (192.38–415.42) b | 67.35 (52.94–81.76) | 0.62 (0.55) |
| C-BARQ dog-directed aggression | |||
| Under median | 394.89 (327.73–462.05) | 61.66 (53.78–69.53) | 0.15 (0.03) |
| Over median | 348.81 (267.45–430.17) | 59.39 (51.76–67.02) | 1.56 (0.91) |
| C-BARQ dog-directed fear | |||
| Under median | 449.32 (378.88–519.75) a | 57.36 (51.96–62.75) | 0.21 (0.24) |
| Over median | 318.03 (250.60–385.99) b | 63.71 (54.44–72.99) | 1.38 (0.78) |
| C-BARQ rivalry | |||
| Under median | 421.10 (338.25–503.94) | 61.97 (52.81–71.13) | 0.02 (0.08) |
| Over median | 331.96 (258.93–404.99) | 55.61 (50.91–60.31) | 0.94 (0.54) |
| C-BARQ chasing | |||
| Under median | 414.29 (52.96–70.87) | 61.92 (52.96–70.87) | 0.66 (0.55) |
| Over median | 344.29 (52.64–66.80) | 59.72 (52.64–66.80) | 2.78 (0.78) |
| C-BARQ stranger-directed fear | |||
| Under median | 397.87 (336.00–459.75) | 62.76 (54.40–71.12) | 0.05 (0.10) |
| Over median | 344.20 (253.34–435.06) | 57.34 (52.59–62.08) | 1.58 (1.01) |
| C-BARQ nonsocial fear | |||
| Under median | 438.03 (366.61–509.45) a | 61.49 (54.85–68.13) | 0.27 (0.19) |
| Over median | 319.77 (251.00–388.54) b | 59.91 (50.79–69.03) | 1.54 (0.77) |
| C-BARG separation-related problems | |||
| Under median | 387.91 (308.59–467.23) | 63.22 (54.84–71.60) | 0.16 (0.17) |
| Over median | 366.43 (297.99–434.87) | 58.30 (50.94–65.67) | 1.18 (0.48) |
| C-BARQ sensitivity | |||
| Under median | 407.42 (340.31–474.52) | 60.95 (54.84–67.05) | 0.23 (0.21) |
| Over median | 321.65 (246.73–396.58) | 60.26 (48.60–71.92) | 1.34 (0.70) |
| C-BARQ excitability | |||
| Under median | 414.16 (346.02–482.31) | 62.92 (54.08–71.76) | 1.09 (0.36) |
| Over median | 336.64 (260.02–413.27) | 58.24 (51.84–64.64) | 2.39 (0.50) |
| C-BARQ attachment | |||
| Under median | 419.28 (344.33–494.23) | 59.31 (50.48–68.14) | 1.36 (0.45) |
| Over median | 334.29 (265.56–403.02) | 62.17 (55.41–68.94) | 3.08 (0.62) |
| C-BARQ energy | |||
| Under median | 392.08 (326.48–457.69) | 65.62 (57.16–74.08) a | 2.08 (1.12) |
| Over median | 354.84 (270.15–439.54) | 53.81 (49.13–58.49) b | 3.81 (0.72) |
| Coeffcients | ADHD-Like R2 = 0.4698 | DIAS Total R2 = 0.2119 | DIAS BR R2 = 0.2134 | DIAS A R2 = 0.1716 | Dog ARS AI R2 = 0.08921 |
|---|---|---|---|---|---|
| Serotonine | −0.0010 | −0.0007 | −0.0005 | −0.0005 | −0.0004 |
| Dopamine | −0.0057 | −0.0013 | |||
| Gender Male * | 0.0609 | 0.0411 | 0.1427 | 0.0207 | - |
| Neutered Entire ** | 0.0630 | 0.0230 | 0.0317 | - | - |
| Age | −0.0025 | - | −0.0002 | - | - |
| Age at acquisition | - | - | |||
| Weight | - | - | - | - | - |
| Dog activity pattern *** | - | - | - | −0.3389 | - |
| First dog | - | - | - | −0.0180 | - |
| Coeffcients | Dog-Directed Fear R2 = 0.4380 | Rivalry R2 = 0.25117 | Nonsocial Fear R2 = 0.1399 | Sensitivity R2 = 0.2143 | Attachment R2 = 0.2284 | Energy R2 = 0.2638 |
|---|---|---|---|---|---|---|
| Serotonine | −0.0006 | −0.0006 | −0.0004 | −0.0001 | −0.0005 | |
| Dopamine | −0.0044 | |||||
| Gender Male * | 0.3686 | −0.0129 | 0.1800 | |||
| Neutered Entire ** | 0.1095 | 0.1564 | ||||
| Age | −0.0046 | |||||
| Age at acquisition | −0.0908 | −0.1593 | ||||
| Weight | −0.0107 | 0.0028 | ||||
| Dog activity pattern *** | −0.0406 | |||||
| First dog | 0.2281 | −0.2629 | −0.2646 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Martínez, Á.; Muñiz de Miguel, S.; Graña, N.; Costas, X.; Diéguez, F.J. Serotonin and Dopamine Blood Levels in ADHD-Like Dogs. Animals 2023, 13, 1037. https://doi.org/10.3390/ani13061037
González-Martínez Á, Muñiz de Miguel S, Graña N, Costas X, Diéguez FJ. Serotonin and Dopamine Blood Levels in ADHD-Like Dogs. Animals. 2023; 13(6):1037. https://doi.org/10.3390/ani13061037
Chicago/Turabian StyleGonzález-Martínez, Ángela, Susana Muñiz de Miguel, Noemi Graña, Xiana Costas, and Francisco Javier Diéguez. 2023. "Serotonin and Dopamine Blood Levels in ADHD-Like Dogs" Animals 13, no. 6: 1037. https://doi.org/10.3390/ani13061037
APA StyleGonzález-Martínez, Á., Muñiz de Miguel, S., Graña, N., Costas, X., & Diéguez, F. J. (2023). Serotonin and Dopamine Blood Levels in ADHD-Like Dogs. Animals, 13(6), 1037. https://doi.org/10.3390/ani13061037