
Restoration of Developmental Competence in Low-Quality Porcine Cumulus–Oocyte Complexes through the Supplementation of Sonic Hedgehog Protein during In Vitro Maturation
 , and
, and 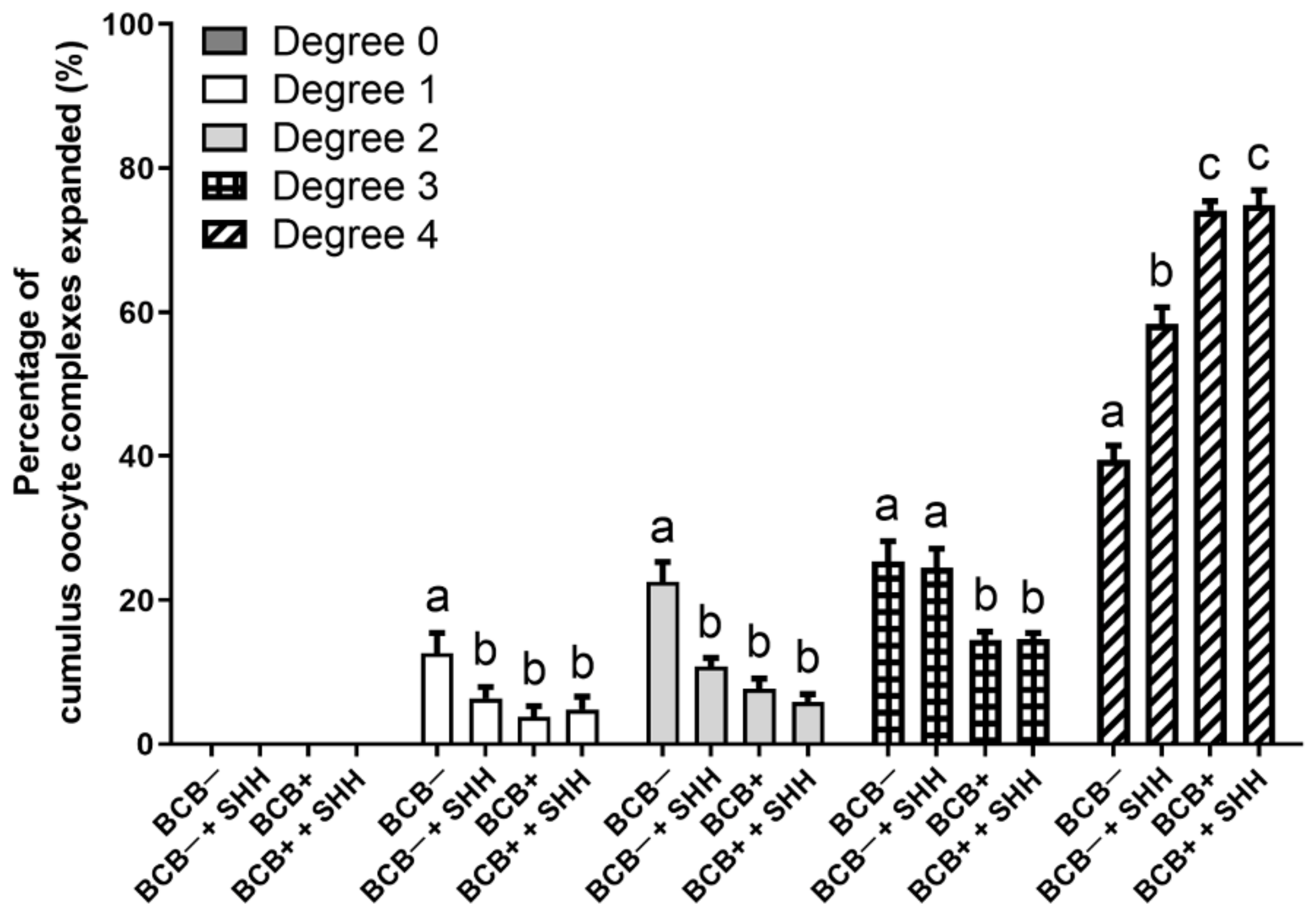{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Isolation and IVM of Porcine COCs
2.3. Evaluation of Cumulus Cell Expansion
2.4. Evaluation of Nuclear Maturation of Oocytes
2.5. Parthenogenetic Activation (PA) of Oocytes
2.6. CDX2 Staining by Immunocytochemistry
2.7. TUNEL Assay
2.8. Statistical Analysis
3. Results
3.1. Effect of SHH Protein Supplementation during Porcine IVM on Cumulus Cell Expansion of BCB− and BCB+ COCs
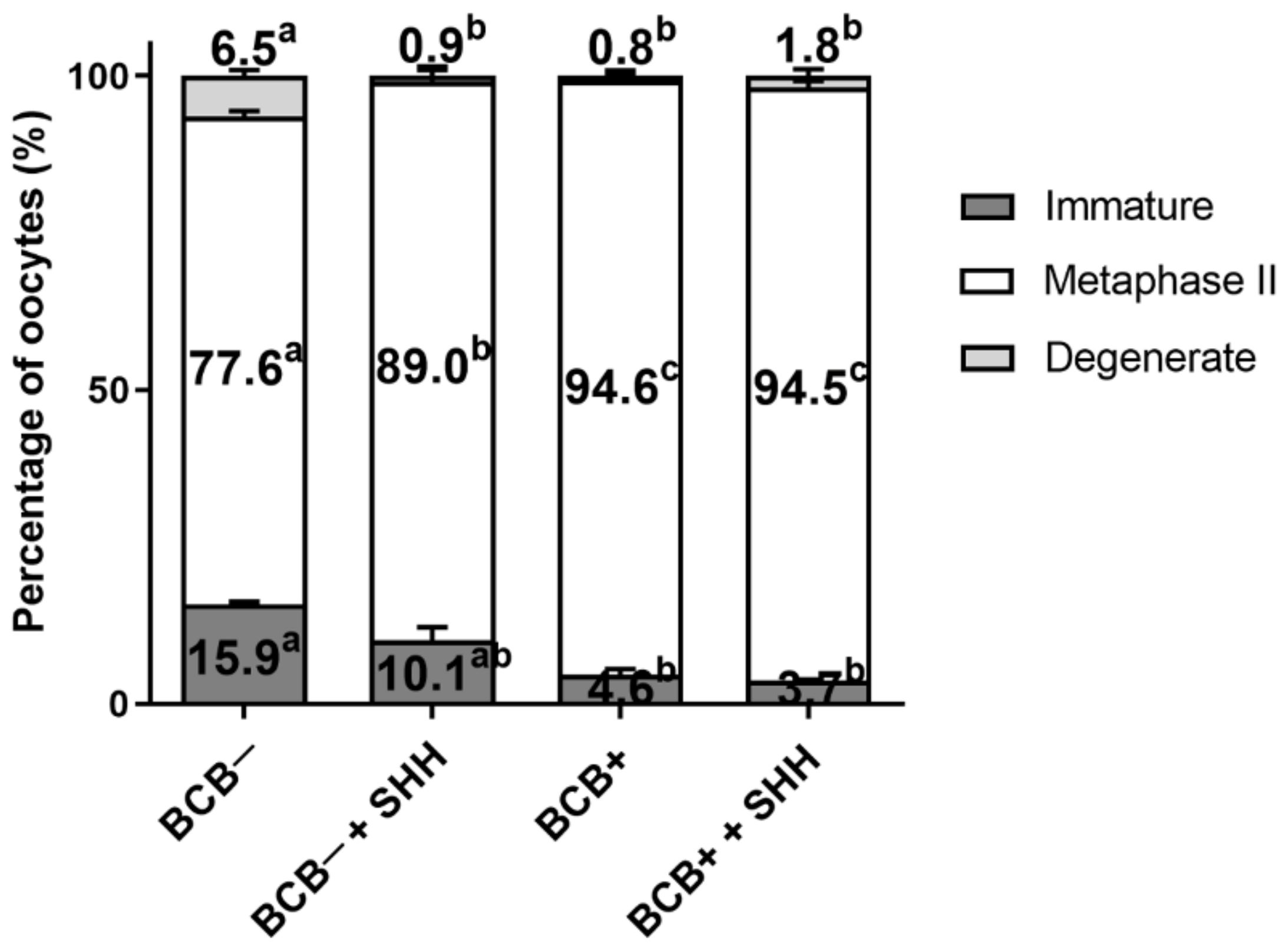
3.2. Effect of SHH Protein Supplementation during Porcine IVM on Oocyte Nuclear Maturation of BCB− and BCB+ COCs
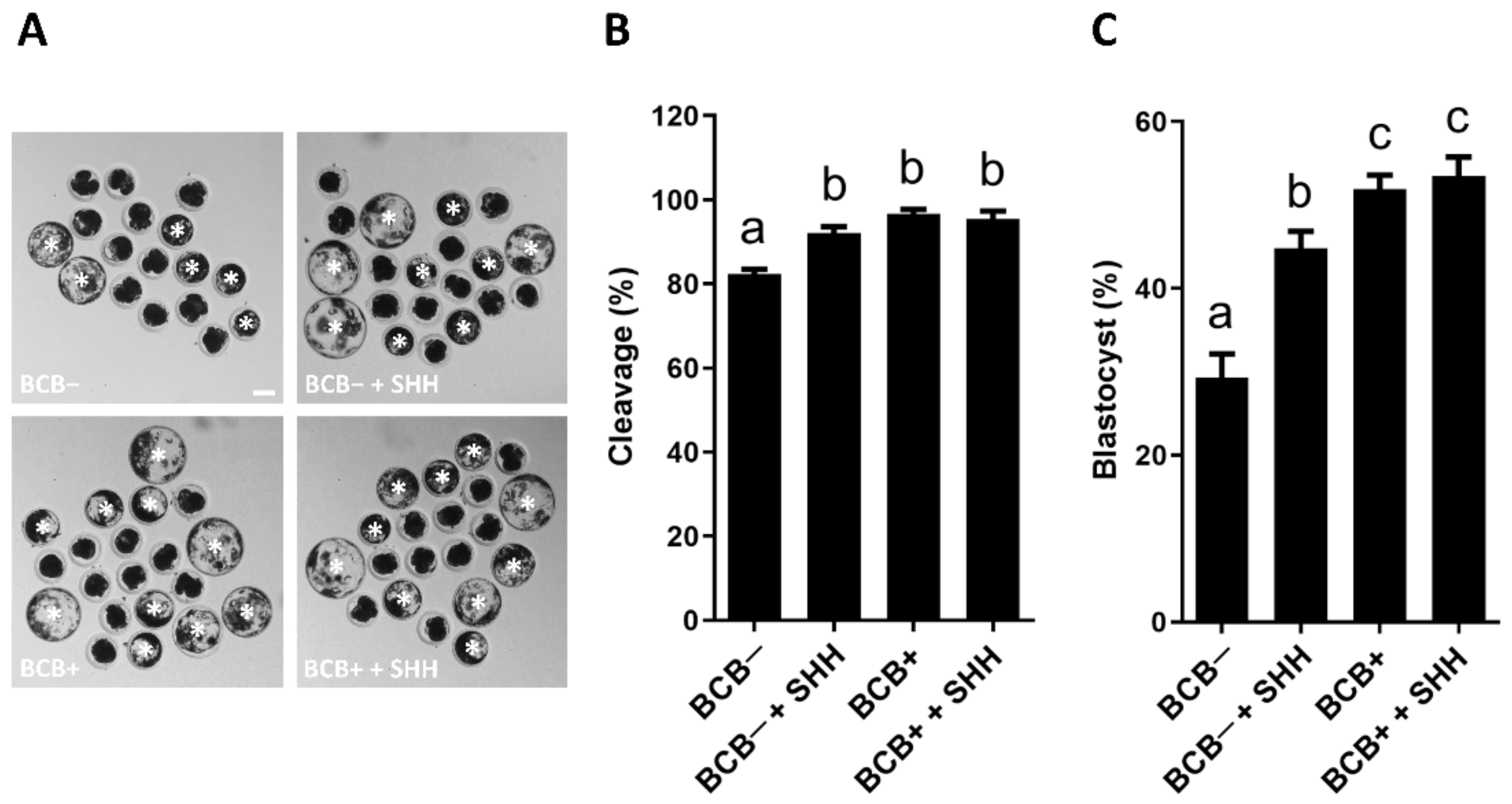
3.3. Effect of SHH Protein Supplementation during Porcine IVM on the Subsequent Development of PA Embryos Derived from BCB− and BCB+ COCs
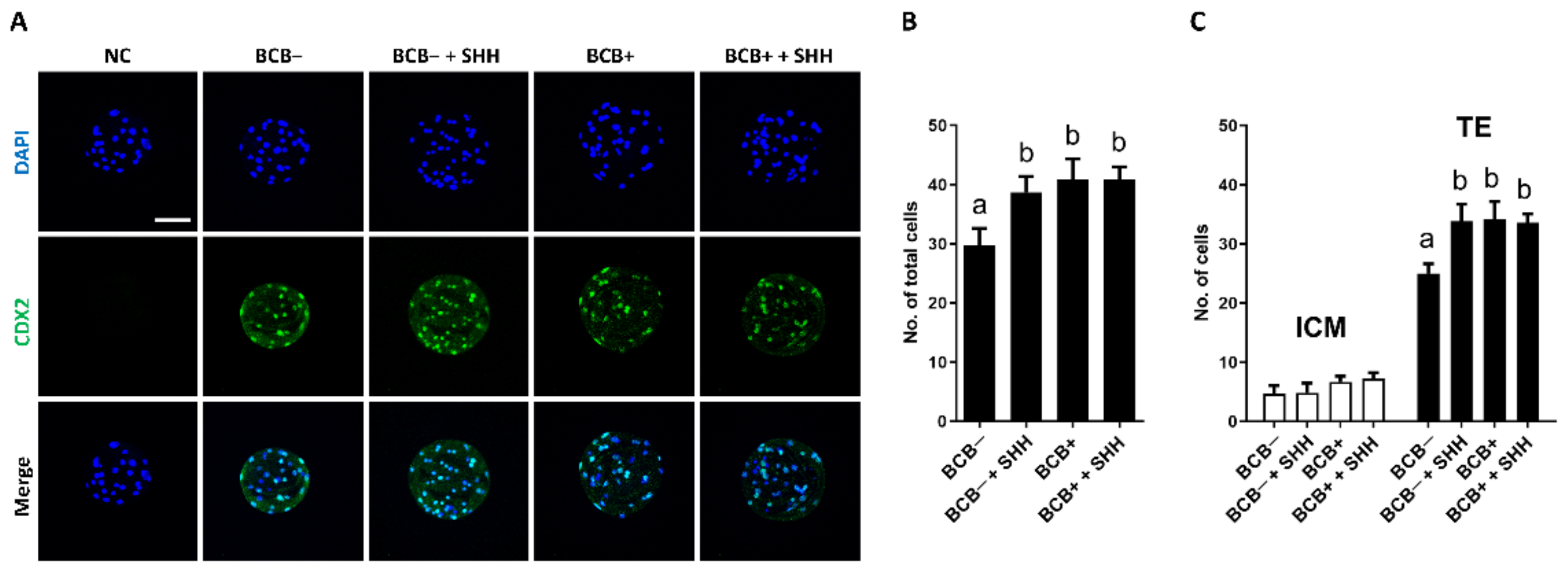
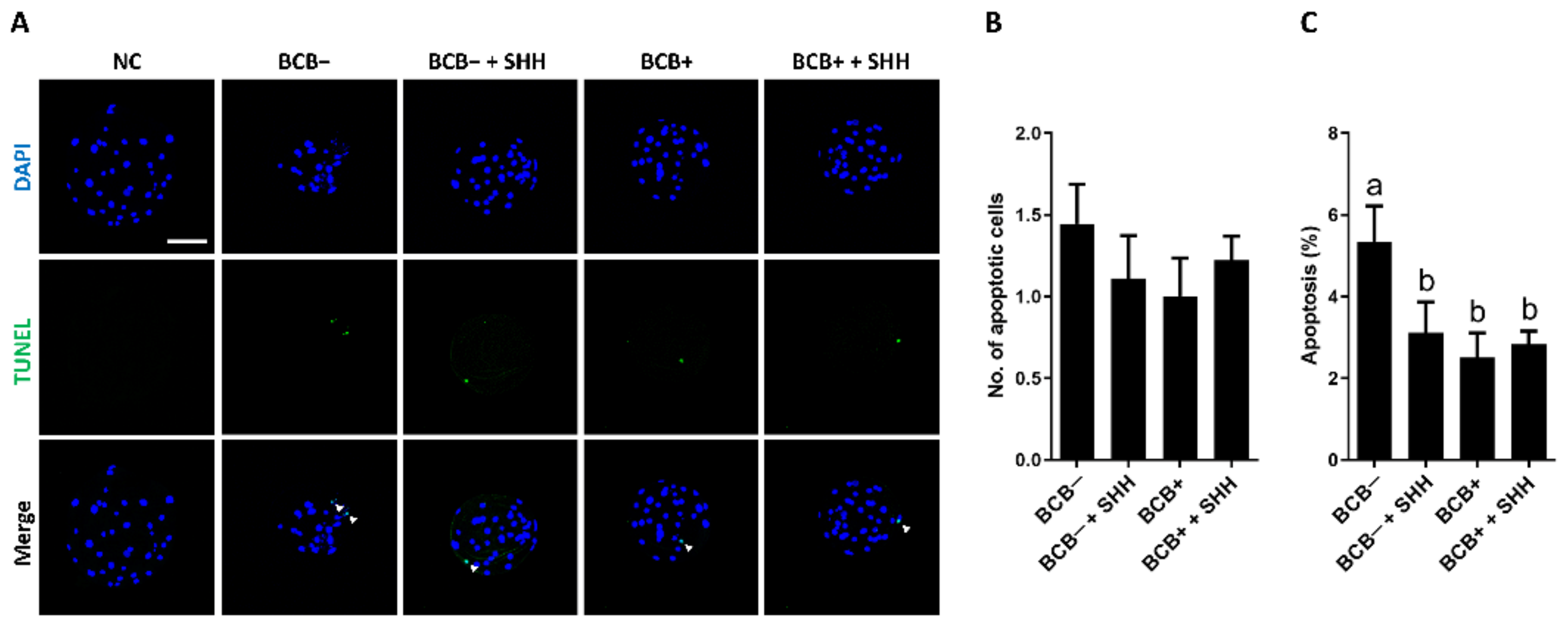
3.4. Effect of SHH Protein Supplementation during Porcine IVM on Quality and Apoptosis Levels in PA Blastocysts Derived from BCB− and BCB+ COCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karja, N.W.K. Nuclear maturation of porcine oocytes in vitro: Effect of the cumulus-oocyte complexes quality. Indones. J. Biotechnol. 2008, 13, 1078–1084. [Google Scholar]
- Warzych, E.; Cieslak, A.; Pawlak, P.; Renska, N.; Pers-Kamczyc, E.; Lechniak, D. Maternal nutrition affects the composition of follicular fluid and transcript content in gilt oocytes. Vet. Med. 2011, 56, 156–167. [Google Scholar] [CrossRef]
- Wongsrikeao, P.; Otoi, T.; Yamasaki, H.; Agung, B.; Taniguchi, M.; Naoi, H.; Shimizu, R.; Nagai, T. Effects of single and double exposure to brilliant cresyl blue on the selection of porcine oocytes for in vitro production of embryos. Theriogenology 2006, 66, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Mangia, F.; Epstein, C.J. Biochemical studies of growing mouse oocytes: Preparation of oocytes and analysis of glucose-6-phosphate dehydrogenase and lactate dehydrogenase activities. Dev. Biol. 1975, 45, 211–220. [Google Scholar] [CrossRef]
- Opiela, J.; Katska-Ksiazkiewicz, L. The utility of Brilliant Cresyl Blue (BCB) staining of mammalian oocytes used for in vitro embryo production (IVP). Reprod. Biol. 2013, 13, 177–183. [Google Scholar] [CrossRef]
- Walker, B.N.; Nix, J.; Wilson, C.; Marrella, M.A.; Speckhart, S.L.; Wooldridge, L.; Yen, C.N.; Bodmer, J.S.; Kirkpatrick, L.T.; Moorey, S.E.; et al. Tight gene co-expression in BCB positive cattle oocytes and their surrounding cumulus cells. Reprod. Biol. Endocrinol. RBE 2022, 20, 119. [Google Scholar] [CrossRef]
- Catala, M.G.; Izquierdo, D.; Rodriguez-Prado, M.; Hammami, S.; Paramio, M.T. Effect of oocyte quality on blastocyst development after in vitro fertilization (IVF) and intracytoplasmic sperm injection (ICSI) in a sheep model. Fertil. Steril. 2012, 97, 1004–1008. [Google Scholar] [CrossRef]
- Lee, S.; Kang, H.G.; Jeong, P.S.; Nanjidsuren, T.; Song, B.S.; Jin, Y.B.; Lee, S.R.; Kim, S.U.; Sim, B.W. Effect of Oocyte Quality Assessed by Brilliant Cresyl Blue (BCB) Staining on Cumulus Cell Expansion and Sonic Hedgehog Signaling in Porcine during In Vitro Maturation. Int. J. Mol. Sci. 2020, 21, 4423. [Google Scholar] [CrossRef]
- Ingham, P.W.; McMahon, A.P. Hedgehog signaling in animal development: Paradigms and principles. Genes Dev. 2001, 15, 3059–3087. [Google Scholar] [CrossRef]
- Varjosalo, M.; Li, S.P.; Taipale, J. Divergence of hedgehog signal transduction mechanism between Drosophila and mammals. Dev. Cell 2006, 10, 177–186. [Google Scholar] [CrossRef]
- Doheny, D.; Manore, S.G.; Wong, G.L.; Lo, H.W. Hedgehog Signaling and Truncated GLI1 in Cancer. Cells 2020, 9, 2114. [Google Scholar] [CrossRef]
- Russell, M.C.; Cowan, R.G.; Harman, R.M.; Walker, A.L.; Quirk, S.M. The hedgehog signaling pathway in the mouse ovary. Biol. Reprod. 2007, 77, 226–236. [Google Scholar] [CrossRef]
- Wang, D.C.; Huang, J.C.; Lo, N.W.; Chen, L.R.; Mermillod, P.; Ma, W.L.; Chiang, H.I.; Ju, J.C. Sonic Hedgehog promotes in vitro oocyte maturation and term development of embryos in Taiwan native goats. Theriogenology 2017, 103, 52–58. [Google Scholar] [CrossRef]
- Spicer, L.J.; Sudo, S.; Aad, P.Y.; Wang, L.S.; Chun, S.Y.; Ben-Shlomo, I.; Klein, C.; Hsueh, A.J. The hedgehog-patched signaling pathway and function in the mammalian ovary: A novel role for hedgehog proteins in stimulating proliferation and steroidogenesis of theca cells. Reproduction 2009, 138, 329–339. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Lin, D.P.; Yen, S.Y.; Tseng, J.K.; Chuang, J.F.; Chen, B.Y.; Lin, T.A.; Chang, H.H.; Ju, J.C. Sonic hedgehog promotes porcine oocyte maturation and early embryo development. Reprod. Fertil. Dev. 2009, 21, 805–815. [Google Scholar] [CrossRef]
- Vanderhyden, B.C.; Caron, P.J.; Buccione, R.; Eppig, J.J. Developmental pattern of the secretion of cumulus expansion-enabling factor by mouse oocytes and the role of oocytes in promoting granulosa cell differentiation. Dev. Biol. 1990, 140, 307–317. [Google Scholar] [CrossRef]
- Lee, S.; Kang, H.G.; Jeong, P.S.; Kim, M.J.; Park, S.H.; Song, B.S.; Sim, B.W.; Kim, S.U. Heat stress impairs oocyte maturation through ceramide-mediated apoptosis in pigs. Sci. Total Environ. 2021, 755, 144144. [Google Scholar] [CrossRef]
- Kang, H.G.; Jeong, P.S.; Kim, M.J.; Joo, Y.E.; Gwon, M.A.; Jeon, S.B.; Song, B.S.; Kim, S.U.; Lee, S.; Sim, B.W. Arsenic exposure during porcine oocyte maturation negatively affects embryonic development by triggering oxidative stress-induced mitochondrial dysfunction and apoptosis. Toxicology 2022, 480, 153314. [Google Scholar] [CrossRef]
- Yang, H.J.; Lee, S.; Sim, B.W.; Jeong, P.S.; Choi, S.A.; Park, Y.H.; Song, B.S.; Yoon, S.B.; Kang, P.; Jeong, K.J.; et al. Transient meiotic arrest maintained by DON (6-diazo-5-oxo-l-norleucine) enhances nuclear/cytoplasmic maturation of porcine oocytes. Reproduction 2019, 158, 543–554. [Google Scholar] [CrossRef]
- Joo, Y.E.; Jeong, P.S.; Lee, S.; Jeon, S.B.; Gwon, M.A.; Kim, M.J.; Kang, H.G.; Song, B.S.; Kim, S.U.; Cho, S.K.; et al. Anethole improves the developmental competence of porcine embryos by reducing oxidative stress via the sonic hedgehog signaling pathway. J. Anim. Sci. Biotechnol. 2023, 14, 32. [Google Scholar] [CrossRef]
- Jeong, P.S.; Lee, S.; Park, S.H.; Kim, M.J.; Kang, H.G.; Nanjidsuren, T.; Son, H.C.; Song, B.S.; Koo, D.B.; Sim, B.W.; et al. Butylparaben Is Toxic to Porcine Oocyte Maturation and Subsequent Embryonic Development Following In Vitro Fertilization. Int. J. Mol. Sci. 2020, 21, 3692. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Jeong, B.S.; Yang, X. Dynamic changes of cumulus-oocyte cell communication during in vitro maturation of porcine oocytes. Biol. Reprod. 2000, 63, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Turathum, B.; Gao, E.M.; Chian, R.C. The Function of Cumulus Cells in Oocyte Growth and Maturation and in Subsequent Ovulation and Fertilization. Cells 2021, 10, 2292. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.M.; Dunning, K.R.; Sutton-McDowall, M.; Gilchrist, R.B.; Thompson, J.G.; Russell, D.L. Failure to launch: Aberrant cumulus gene expression during oocyte in vitro maturation. Reproduction 2017, 153, R109–R120. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Cho, J. The role of sonic hedgehog signaling pathway in in vitro oocyte maturation. J. Anim. Reprod. Biotechnol. 2021, 36, 183–188. [Google Scholar] [CrossRef]
- Lee, S.; Jin, J.X.; Taweechaipaisankul, A.; Kim, G.A.; Ahn, C.; Lee, B.C. Melatonin influences the sonic hedgehog signaling pathway in porcine cumulus oocyte complexes. J. Pineal Res. 2017, 63, e12424. [Google Scholar] [CrossRef] [PubMed]
- Eppig, J.J.; Schroeder, A.C.; O’Brien, M.J. Developmental capacity of mouse oocytes matured in vitro: Effects of gonadotrophic stimulation, follicular origin and oocyte size. J. Reprod. Fertil. 1992, 95, 119–127. [Google Scholar] [CrossRef]
- Lonergan, P.; Monaghan, P.; Rizos, D.; Boland, M.P.; Gordon, I. Effect of follicle size on bovine oocyte quality and developmental competence following maturation, fertilization, and culture in vitro. Mol. Reprod. Dev. 1994, 37, 48–53. [Google Scholar] [CrossRef]
- Trounson, A.; Anderiesz, C.; Jones, G. Maturation of human oocytes in vitro and their developmental competence. Reproduction 2001, 121, 51–75. [Google Scholar] [CrossRef]
- Lee, S.; Jin, J.X.; Taweechaipaisankul, A.; Kim, G.A.; Ahn, C.; Lee, B.C. Sonic hedgehog signaling mediates resveratrol to improve maturation of pig oocytes in vitro and subsequent preimplantation embryo development. J. Cell. Physiol. 2018, 233, 5023–5033. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, P.-S.; Kang, H.-G.; Song, B.-S.; Kim, S.-U.; Sim, B.-W.; Lee, S. Restoration of Developmental Competence in Low-Quality Porcine Cumulus–Oocyte Complexes through the Supplementation of Sonic Hedgehog Protein during In Vitro Maturation. Animals 2023, 13, 1001. https://doi.org/10.3390/ani13061001
Jeong P-S, Kang H-G, Song B-S, Kim S-U, Sim B-W, Lee S. Restoration of Developmental Competence in Low-Quality Porcine Cumulus–Oocyte Complexes through the Supplementation of Sonic Hedgehog Protein during In Vitro Maturation. Animals. 2023; 13(6):1001. https://doi.org/10.3390/ani13061001
Chicago/Turabian StyleJeong, Pil-Soo, Hyo-Gu Kang, Bong-Seok Song, Sun-Uk Kim, Bo-Woong Sim, and Sanghoon Lee. 2023. "Restoration of Developmental Competence in Low-Quality Porcine Cumulus–Oocyte Complexes through the Supplementation of Sonic Hedgehog Protein during In Vitro Maturation" Animals 13, no. 6: 1001. https://doi.org/10.3390/ani13061001
APA StyleJeong, P.-S., Kang, H.-G., Song, B.-S., Kim, S.-U., Sim, B.-W., & Lee, S. (2023). Restoration of Developmental Competence in Low-Quality Porcine Cumulus–Oocyte Complexes through the Supplementation of Sonic Hedgehog Protein during In Vitro Maturation. Animals, 13(6), 1001. https://doi.org/10.3390/ani13061001