

Molecular Identification of Sarcocystis rileyi and Sarcocystis sp. (Closely Related to Sarcocystis wenzeli) in Intestines of Mustelids from Lithuania

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Isolation of Oocysts/Sporocysts
2.2. Molecular Identification and Phylogenetic Analysis
2.3. Data Analysis
3. Results
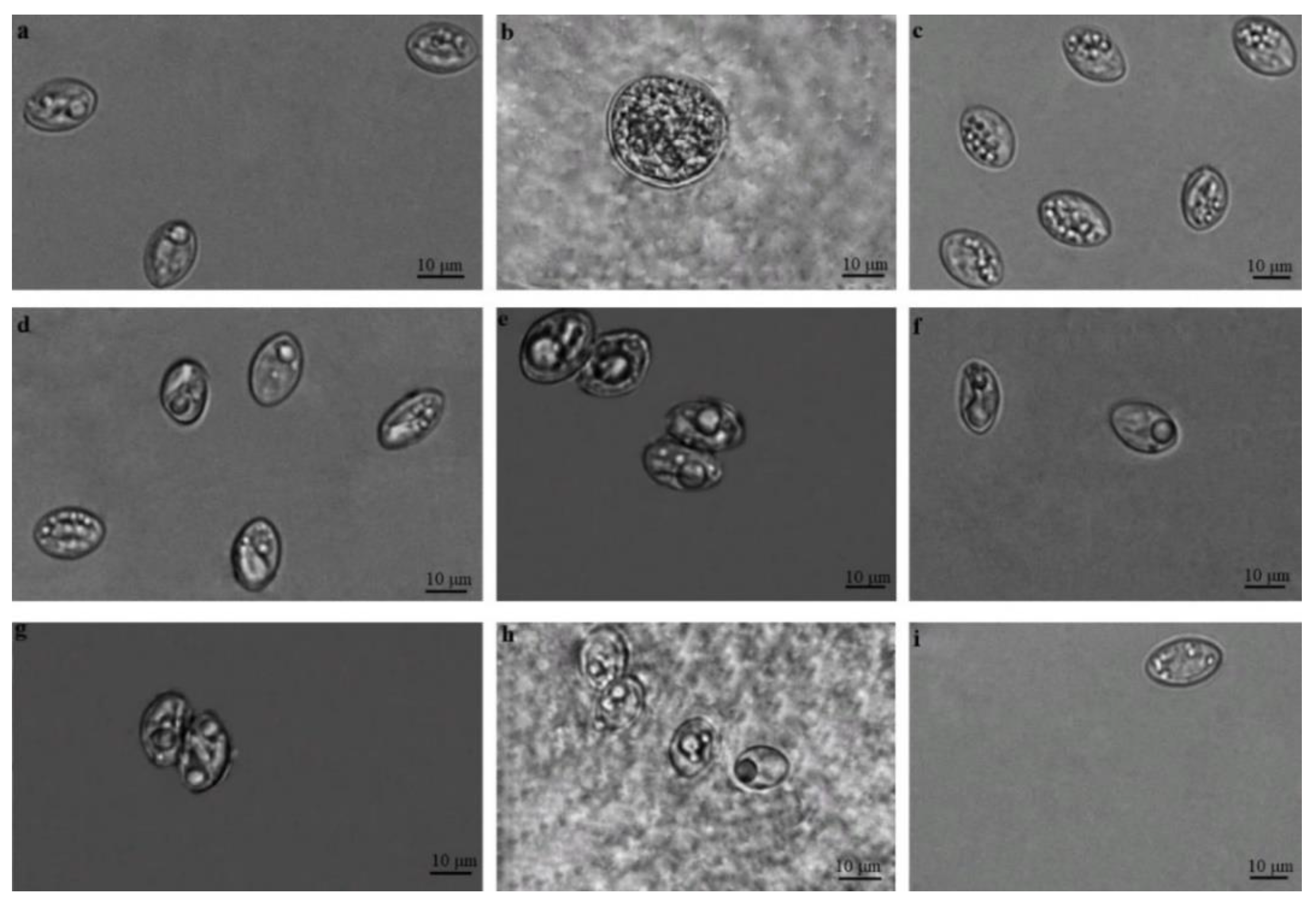
3.1. Microscopical Examination of Sarcocystis spp. Oocysts/Sporocysts
3.2. Sarcocystis Species Identification and Their Distribution in Intestine Samples of Mustelids
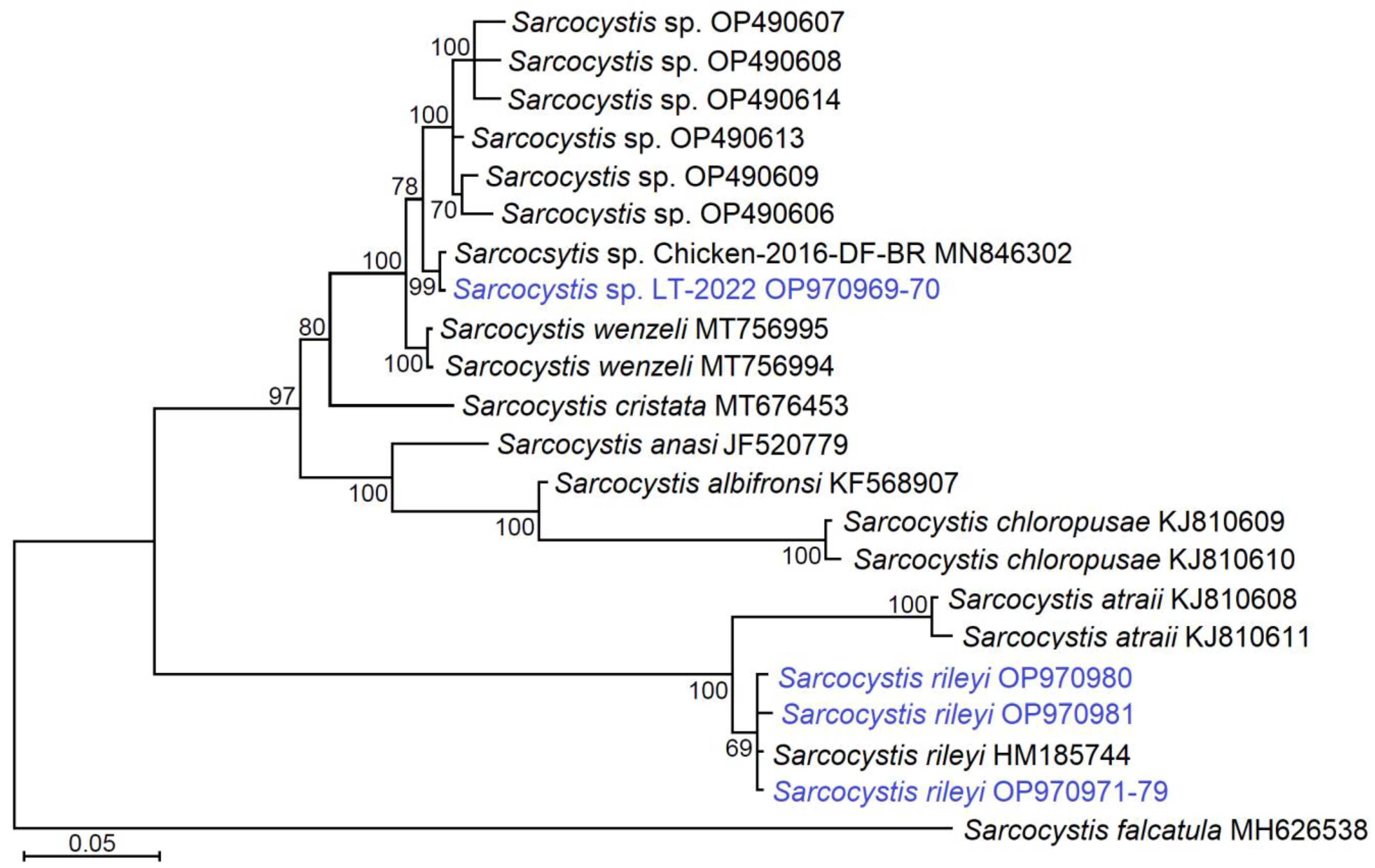
3.3. Phylogenetic Relationships of Identified Sarcocystis Species
4. Discussion
4.1. The Role of Mustelids in Distribution of Sarcocystis Species
4.2. Mustelids as Possible Definitive Hosts of S. rileyi
4.3. Detection of Sarcocystis sp. Closely Related to S. wenzeli in Small Intestine of Mustelids
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Sarcocystosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Mehlhorn, H.; Heydorn, A.O. The Sarcosporidia (Protozoa, Sporozoa): Life Cycle and Fine Structure. Adv. Parasitol. 1978, 16, 43–91. [Google Scholar] [CrossRef] [PubMed]
- Tenter, A.M. Current research on Sarcocystis species of domestic animals. Int. J. Parasitol. 1995, 25, 1311–1330. [Google Scholar] [CrossRef]
- Juozaitytė-Ngugu, E.; Švažas, S.; Šneideris, D.; Rudaitytė-Lukošienė, E.; Butkauskas, D.; Prakas, P. The Role of Birds of the Family Corvidae in Transmitting Sarcocystis Protozoan Parasites. Animals 2021, 11, 3258. [Google Scholar] [CrossRef] [PubMed]
- Máca, O.; González-Solís, D. Sarcocystis cristata sp. nov. (Apicomplexa, Sarcocystidae) in the imported great blue turaco Corythaeola cristata (Aves, Musophagidae). Parasites Vectors 2021, 14, 56. [Google Scholar] [CrossRef]
- Gallo, S.S.M.; Lindsay, D.S.; Ederli, N.B.; Matteoli, F.P.; Venancio, T.M.; de Oliveira, F.C.R. Identification of opossums Didelphis aurita (Wied-Neuweid, 1826) as a definitive host of Sarcocystis falcatula-like sporocysts. Parasitol. Res. 2018, 117, 213–223. [Google Scholar] [CrossRef]
- Kutkienė, L.; Prakas, P.; Sruoga, A.; Butkauskas, D. Description of Sarcocystis anasi sp. nov. and Sarcocystis albifronsi sp. nov. in birds of the order Anseriformes. Parasitol. Res. 2012, 110, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Prakas, P.; Oksanen, A.; Butkauskas, D.; Sruoga, A.; Kutkienė, L.; Švažas, S.; Isomursu, M.; Liaugaudaitė, S. Identification and Intraspecific Genetic Diversity of Sarcocystis rileyi from Ducks, Anas spp., in Lithuania and Finland. J. Parasitol. 2014, 100, 657–661. [Google Scholar] [CrossRef]
- El-Morsey, A.; El-Seify, M.; Desouky, A.-R.Y.; Abdel-Aziz, M.M.; El-Dakhly, K.M.; Kasem, S.; Abdo, W.; Haridy, M.; Sakai, H.; Yanai, T. Morphologic and molecular characteristics of Sarcocystis atraii n. sp. (Apicomplexa: Sarcocystidae) infecting the common coot (Fulica atra) from Egypt. Acta Parasitol. 2015, 60, 691–699. [Google Scholar] [CrossRef]
- El-Morsey, A.; El-Seify, M.; Desouky, A.Y.; Abdel-Aziz, M.M.; Sakai, H.; Yanai, T. Sarcocystis chloropusae (protozoa: Sarcocystidae) n. sp. from the common moorhen (Gallinula chloropus) from Egypt. Parasitology 2015, 142, 1063–1065. [Google Scholar] [CrossRef]
- Pan, J.; Ma, C.; Huang, Z.; Ye, Y.; Zeng, H.; Deng, S.; Hu, J.; Tao, J. Morphological and molecular characterization of Sarcocystis wenzeli in chickens (Gallus gallus) in China. Parasites Vectors 2020, 13, 512. [Google Scholar] [CrossRef]
- Kutkienė, L.; Prakas, P.; Sruoga, A.; Butkauskas, D. Identification of Sarcocystis rileyi from the mallard duck (Anas platyrhynchos) in Europe: Cyst morphology and results of DNA analysis. Parasitol. Res. 2011, 108, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.; Sørensen, S.R.; Kania, P.W.; Buchmann, K. Sarcocystis rileyi (Apicomplexa) in Anas platyrhynchos in Europe with a potential for spread. Int. J. Parasitol. Parasites Wildl. 2021, 15, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Prakas, P.; Kirillova, V.; Gavarāne, I.; Grāvele, E.; Butkauskas, D.; Rudaitytė-Lukošienė, E.; Kirjušina, M. Morphological and molecular description of Sarcocystis ratti n. sp. from the black rat (Rattus rattus) in Latvia. Parasitol. Res. 2019, 118, 2689–2694. [Google Scholar] [CrossRef] [PubMed]
- Gjerde, B. Phylogenetic relationships among Sarcocystis species in cervids, cattle and sheep inferred from the mitochondrial cytochrome c oxidase subunit I gene. Int. J. Parasitol. 2013, 43, 579–591. [Google Scholar] [CrossRef]
- Prakas, P.; Rudaitytė-Lukošienė, E.; Šneideris, D.; Butkauskas, D. Invasive American mink (Neovison vison) as potential definitive host of Sarcocystis elongata, S. entzerothi, S. japonica, S. truncata and S. silva using different cervid species as intermediate hosts. Parasitol. Res. 2021, 120, 2243–2250. [Google Scholar] [CrossRef]
- Prakas, P.; Butkauskas, D.; Švažas, S.; Stanevičius, V. Morphological and genetic characterisation of Sarcocystis halieti from the great cormorant (Phalacrocorax carbo). Parasitol. Res. 2018, 117, 3663–3667. [Google Scholar] [CrossRef]
- Kontrimavičius, V. Lietuvos Fauna: Žinduoliai; Mokslas: Vilnius, Lithuania, 1988. [Google Scholar]
- Balčiauskas, L.; Trakimas, G.; Juškaitis, R.; Ulevičius, A.; Balčiauskienė, L. Atlas of Lithuanian Mammals, Amphibians and Reptiles, 2nd ed.; Akstis: Vilnius, Lithuania, 1999. [Google Scholar]
- Baghli, A.; Engel, E.; Verhagen, R. Feeding habits and trophic niche overlap of two sympatric mustelidae, the polecat Mustela putorius and the beech marten Martes foina. Z. Für Jagdwiss. 2002, 48, 217–225. [Google Scholar] [CrossRef]
- Koepfli, K.-P.; Deere, K.A.; Slater, G.J.; Begg, C.; Begg, K.; Grassman, L.; Lucherini, M.; Veron, G.; Wayne, R.K. Multigene phylogeny of the Mustelidae: Resolving relationships, tempo and biogeographic history of a mammalian adaptive radiation. BMC Biol. 2008, 6, 10. [Google Scholar] [CrossRef]
- Newman, C.; Zhou, Y.-B.; Buesching, C.D.; Kaneko, Y.; Macdonald, D.W. Contrasting Sociality in Two Widespread, Generalist, Mustelid Genera, Meles and Martes. Mammal Study 2011, 36, 169–188. [Google Scholar] [CrossRef]
- Law, C.J.; Slater, G.J.; Mehta, R.S. Lineage Diversity and Size Disparity in Musteloidea: Testing Patterns of Adaptive Radiation Using Molecular and Fossil-Based Methods. Syst. Biol. 2018, 67, 127–144. [Google Scholar] [CrossRef]
- Prakas, P.; Balčiauskas, L.; Juozaitytė-Ngugu, E.; Butkauskas, D. The Role of Mustelids in the Transmission of Sarcocystis spp. Using Cattle as Intermediate Hosts. Animals 2021, 11, 822. [Google Scholar] [CrossRef]
- Bonesi, L.; Palazon, S. The American mink in Europe: Status, impacts, and control. Biol. Conserv. 2007, 134, 470–483. [Google Scholar] [CrossRef]
- Holopainen, S.; Väänänen, V.-M.; Vehkaoja, M.; Fox, A.D. Do alien predators pose a particular risk to duck nests in Northern Europe? Results from an artificial nest experiment. Biol. Invasions 2021, 23, 3795–3807. [Google Scholar] [CrossRef]
- Viksne, J.; Švažas, S.; Czajkowski, A. Atlas of Duck Populations in Eastern Europe; Akstis Press: Vilnius, Lithuania, 2010. [Google Scholar]
- Baltrūnaitė, L. Diet Composition of the Red Fox (Vulpes Vulpes L.), Pine Marten (Martes Martes L.) and Raccoon Dog (Nyctereutes Procyonoides Gray) in Clay Plain Landscape, Lithuania. Acta Zool. Litu. 2002, 12, 362–368. [Google Scholar] [CrossRef]
- Gjerde, B. Molecular characterisation of Sarcocystis rileyi from a common eider (Somateria mollissima) in Norway. Parasitol. Res. 2014, 113, 3501–3509. [Google Scholar] [CrossRef] [PubMed]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Bioinform. Methods Protoc. 2000, 132, 365–386. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Milne, I.; Wright, F.; Rowe, G.; Marshall, D.; Husmeier, D.; McGuire, G. TOPALi: Software for automatic identification of recombinant sequences within DNA multiple alignments. Bioinformatics 2004, 20, 1806–1807. [Google Scholar] [CrossRef]
- Reiczigel, J. Confidence intervals for the binomial parameter: Some new considerations. Stat. Med. 2003, 22, 611–621. [Google Scholar] [CrossRef]
- Reiczigel, J.; Abonyi-Tóth, Z.; Singer, J. An exact confidence set for two binomial proportions and exact unconditional confidence intervals for the difference and ratio of proportions. Comput. Stat. Data Anal. 2008, 52, 5046–5053. [Google Scholar] [CrossRef]
- Rózsa, L.; Reiczigel, J.; Majoros, G. Quantifying Parasites in Samples of Hosts. J. Parasitol. 2000, 86, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.M.; Sousa, S.K.; Paludo, G.R.; de Melo, C.B.; Llano, H.A.; Soares, R.M.; Castro, M.B. An undescribed species of Sarcocystis associated with necrotizing meningoencephalitis in naturally infected backyard chickens in the Midwest of Brazil. Parasitol. Int. 2020, 76, 102098. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Moré, G.; Maksimov, A.; Conraths, F.; Schares, G. Molecular identification of Sarcocystis spp. in foxes (Vulpes vulpes) and raccoon dogs (Nyctereutes procyonoides) from Germany. Veter Parasitol. 2016, 220, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Odening, K. The present state of species-systematics in Sarcocystis Lankester, 1882 (Protista, Sporozoa, Coccidia). Syst. Parasitol. 1998, 41, 209–233. [Google Scholar] [CrossRef]
- Stiles, C.W. On the presence of sarcosporidia in birds. USDA Bur. Anim. Ind. Bull. 1893, 3, 79–89. [Google Scholar]
- Dubey, J.P.; Cawthorn, R.J.; Speer, C.A.; Wobeser, G.A. Redescription of the sarcocysts of Sarcocystis rileyi (Apicomplexa: Sarcocystidae). J. Eukaryot. Microbiol. 2003, 50, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Erickson, A.B. Sarcocystis in Birds. Auk 1940, 57, 514–519. [Google Scholar] [CrossRef]
- Cornwell, G. New Waterfowl Host Records for Sarcocystis rileyi and a Review of Sarcosporidiosis in Birds. Avian Dis. 1963, 7, 212. [Google Scholar] [CrossRef]
- Chabreck, R.H. Sarcosporidiosis in Ducks in Louisiana. Trans. N. Am. Wildl. Conf. 1965, 30, 174–184. [Google Scholar]
- Drouin, T.E.; Mahrt, J.L. The Prevalence of Sarcocystis Lankester, 1882, in some Bird Species in Western Canada, with Notes on its Life Cycle. Can. J. Zool. 1979, 57, 1915–1921. [Google Scholar] [CrossRef] [PubMed]
- Fedynich, A.M.; Pence, D.B. Sarcocystis in Mallards on the Southern High Plains of Texas. Avian Dis. 1992, 36, 1067. [Google Scholar] [CrossRef]
- Dubey, J.P.; Rosenthal, B.M.; Felix, T.A. Morphologic and Molecular Characterization of the Sarcocysts of Sarcocystis rileyi (Apicomplexa: Sarcocystidae) from the Mallard Duck (Anas platyrhynchos). J. Parasitol. 2010, 96, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Padilla-Aguilar, P.; Romero-Callejas, E.; Osorio-Sarabia, D.; Ramírez-Lezama, J.; Cigarroa-Toledo, N.; Machain-Williams, C.; Manterola, C.; Zarza, H. Detection and Molecular Identification of Sarcocystis rileyi (Apicomplexa: Sarcocystidae) from a Northern Shoveler (Anas clypeata) in Mexico. J. Wildl. Dis. 2016, 52, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Kalisinska, E.; Betlejewska, K.M.; Schmidt, M.; Gozdzicka-Jozefiak, A.; Tomczyk, G. Protozoal Macrocysts in the Skeletal Muscle of a Mallard duck in Poland: The First Recorded Case. Acta Parasitol. 2003, 48, 1–5. [Google Scholar]
- Prakas, P.; Liaugaudaitė, S.; Kutkienė, L.; Sruoga, A.; Švažas, S. Molecular identification of Sarcocystis rileyi sporocysts in red foxes (Vulpes vulpes) and raccoon dogs (Nyctereutes procyonoides) in Lithuania. Parasitol. Res. 2015, 114, 1671–1676. [Google Scholar] [CrossRef]
- Cromie, R.; Ellis, M. Sarcocystis Survey. Sarcocystis Survey Feedback Report–The UK Wildfowl Sarcocystis Survey. 2019. Available online: www.sarcocystissurvey.org.uk/2015-2018-feedback-report/ (accessed on 15 December 2022).
- Szekeres, S.; Juhász, A.; Kondor, M.; Takács, N.; Sugár, L.; Hornok, S. Sarcocystis rileyi emerging in Hungary: Is rice breast disease underreported in the region? Acta Vet. Hung. 2019, 67, 401–406. [Google Scholar] [CrossRef]
- Muir, A.; Ellis, M.; Blake, D.P.; Chantrey, J.; Strong, E.A.; Reeves, J.P.; Cromie, R.L. Sarcocystis rileyi in UK free-living wildfowl (Anatidae): Surveillance, histopathology and first molecular characterisation. Vet. Rec. 2020, 186, 186. [Google Scholar] [CrossRef]
- Mooij, J.H. Protection and use of Waterbirds in the European Union. Beitr. Jagd Wildforschung 2005, 30, 49–76. [Google Scholar]
- Hirschfeld, A.; Attard, G.; Scott, L. Bird Hunting in Europe: An Analysis of Bag Figures and the Potential Impact on the Conservation of Threatened Species. Br. Birds 2019, 112, 153–166. [Google Scholar]
- Sibille, S.; Griffin, C.; Scallan, D. Europe’s Huntable Birds: A Review of Status and Conservation Priorities. European Federation for Hunting and Conservation (FACE). 2020. Available online: https://www.face.eu/ (accessed on 14 December 2022).
- Friend, M.; Franson, J.C. Field Manual of Wildlife Diseases—General Field Procedures and Diseases of Birds; US Geological Survey: Reston, Virginia, 1999. [Google Scholar]
- Cawthorn, R.J.; Rainnie, D.; Wobeser, G. Experimental transmission of Sarcocystis sp. (Protozoa: Sarcocystidae) between the shoveler (Anas clypeata) duck and the striped skunk (Mephitis mephitis). J. Wildl. Dis. 1981, 17, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Wicht, R.J. Transmission of Sarcocystis rileyi to the striped skunk (Mephitis mephitis). J. Wildl. Dis. 1981, 17, 387–388. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.E.; Reeder, D.M. Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; The Johns Hopkins University Press: Baltimore, MD, USA, 2005; p. 512. [Google Scholar]
- Flynn, J.J.; Finarelli, J.A.; Zehr, S.; Hsu, J.; Nedbal, M.A. Molecular Phylogeny of the Carnivora (Mammalia): Assessing the Impact of Increased Sampling on Resolving Enigmatic Relationships. Syst. Biol. 2005, 54, 317–337. [Google Scholar] [CrossRef] [PubMed]
- Šlapeta, J.R.; Modrý, D.; Votýpka, J.; Jirků, M.; Lukeš, J.; Koudela, B. Evolutionary relationships among cyst-forming coccidia Sarcocystis spp. (Alveolata: Apicomplexa: Coccidea) in endemic African tree vipers and perspective for evolution of heteroxenous life cycle. Mol. Phylogenet. Evol. 2003, 27, 464–475. [Google Scholar] [CrossRef]
- Morrison, D.A.; Bornstein, S.; Thebo, P.; Wernery, U.; Kinne, J.; Mattsson, J.G. The current status of the small subunit rRNA phylogeny of the coccidia (Sporozoa). Int. J. Parasitol. 2004, 34, 501–514. [Google Scholar] [CrossRef]
- Jasiulionis, M.; Stirkė, V.; Balčiauskas, L. Invasive Raccoon Dog (Nyctereutes procyonoides) and Raccoon (Procyon lotor) Monitoring in Lithuania Based on Camera Traps Data. Biol. Life Sci. Forum 2022, 15, 3. [Google Scholar] [CrossRef]
- Mao, J.B.; Zuo, Y.X. Studies on the Prevalence and Experimental Transmission of Sarcocystis sp. in Chickens. Acta Vet. Zootech Sin. 1994, 25, 555–559. [Google Scholar]
- Pak, S.M.; Perminova, V.V.; Yeshtokina, N.V. Sarcocystis citellivulpes sp. n. from the Yellow Suslik Citellus fulvus Lichtenstain, 1923. In Toksoplazmidy, Protozoologiya; Beyer, T.V., Bezukladnikova, N.A., Galuzo, I.G., Konovalova, S.I., Pak, S.M., Eds.; Akademii Nauk Sovetskoi Sotsialisticheskoi Respubliki: Moscow, Russia, 1979. [Google Scholar]
- Švažas, S.; Kozulin, A. Waterbirds of Large Fishponds of Belarus and Lithuania; Institute of Ecology Press: Vilnius, Lithuania, 2002. [Google Scholar]
- Lanszki, J.; Heltai, M. Feeding Habits of Sympatric Mustelids in an Agricultural Area of Hungary. Acta Zool. Acad. Sci. Hung. 2011, 57, 291–304. [Google Scholar]
- Zschille, J.; Stier, N.; Roth, M.; Mayer, R. Feeding habits of invasive American mink (Neovison vison) in northern Germany—Potential implications for fishery and waterfowl. Acta Theriol. 2013, 59, 25–34. [Google Scholar] [CrossRef]
- Tsunoda, H.; Peeva, S.; Raichev, E.; Kronawetter, T.; Kirilov, K.B.; Georgiev, D.; Kaneko, Y. Patterns of spatial distribution and diel activity in carnivore guilds (Carnivora). J. Vertebr. Biol. 2022, 71, 22018-1–22018-11. [Google Scholar] [CrossRef]
- Munday, B.L.; Humphrey, J.D.; Kila, V. Pathology Produced by, Prevalence, of, and Probable Life-cycle of a Species of Sarcocystis in the Domestic Fowl. Avian Dis. 1977, 21, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Mutalib, A.; Keirs, R.; Maslin, W.; Topper, M.; Dubey, J.P. Sarcocystis-Associated Encephalitis in Chickens. Avian Dis. 1995, 39, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Prakas, P.; Kutkienė, L.; Butkauskas, D.; Sruoga, A.; Žalakevičius, M. Molecular and morphological investigations of Sarcocystis corvusi sp. nov. from the jackdaw (Corvus monedula). Parasitol. Res. 2013, 112, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Olias, P.; Olias, L.; Lierz, M.; Mehlhorn, H.; Gruber, A.D. Sarcocystis calchasi is distinct to Sarcocystis columbae sp. nov. from the wood pigeon (Columba palumbus) and Sarcocystis sp. from the sparrowhawk (Accipiter nisus). Vet. Parasitol. 2010, 171, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Kutkienė, L.; Prakas, P.; Butkauskas, D.; Sruoga, A. Description of Sarcocystis turdusi sp. nov. from the common blackbird (Turdus merula). Parasitology 2012, 139, 1438–1443. [Google Scholar] [CrossRef]
- Prakas, P.; Butkauskas, D.; Švažas, S.; Juozaitytė-Ngugu, E.; Stanevičius, V. Morphologic and genetic identification of Sarcocystis fulicae n. sp.(Apicomplexa: Sarcocystidae) from the Eurasian coot (Fulica atra). J. Wildl. Dis. 2018, 54, 765–771. [Google Scholar] [CrossRef]
- Prakas, P.; Butkauskas, D.; Juozaitytė-Ngugu, E. Molecular identification of four Sarcocystis species in the herring gull, Larus argentatus, from Lithuania. Parasites Vectors 2020, 13, 2. [Google Scholar] [CrossRef]
- Juozaitytė-Ngugu, E.; Butkauskas, D.; Švažas, S.; Prakas, P. Investigations on Sarcocystis species in the leg muscles of the bird family Corvidae in Lithuania. Parasitol. Res. 2022, 121, 703–711. [Google Scholar] [CrossRef]
- Prakas, P.; Butkauskas, D.; Juozaitytė-Ngugu, E. Molecular and morphological description of Sarcocystis kutkienae sp. nov. from the common raven (Corvus corax). Parasitol. Res. 2020, 119, 4205–4210. [Google Scholar] [CrossRef]
- Ministry of Agriculture of the Republic of Lithuania. Lithuanian Agrifood Sector. 2020. Available online: https://zum.lrv.lt/uploads/zum/documents/files/LT_versija/Naujiena/Leidiniai/Lithuanian_agrifood_sector_2020.pdf (accessed on 16 December 2022).




{kind=link}
{kind=link}
{kind=link}
| Host Species | Microscopical Detection of Sarcocystis spp. | The Size of Sporocysts | The Size of Sporulating Oocysts | The Size of Free Oocysts | |
|---|---|---|---|---|---|
| Infected/Investigate (%) | 95% CI | ||||
| American mink | 31/61 (50.8%) | 38.5–63.2 | 10.2–14.1 × 7.1–9.4 (12.3 × 8.3; n = 170) | 14.5–21.1 × 10.9–17.5 (18.0 × 14.0; n = 30) | 14.8–23.5 × 13.5–22.4 (21.1 × 18.3; n = 14) |
| European pine marten | 15/26 (57.7%) | 38.3–75.4 | 10.1–14.1 × 6.9–10.1 (11.6 × 8.2; n = 115) | 12.4–19.2 × 10.1–18.3 (14.9 × 12.8; n = 20) | 17.9–23.1 × 15.5–21.5 (21.8 × 17.6; n = 17) |
| European polecat | 11/18 (61.1%) | 37.4–81.5 | 10.0–14.6 × 6.7–9.9 (12.4 × 8.3; n = 140) | 13.3–19.5 × 11.1–18.0 (17.5 × 13.3; n = 40) | 13.9–23.0 × 12.0–22.0 (19.2 × 18.5; n = 15) |
| European badger | 1/6 (16.7%) | 8.6–58.9 | 10.0–14.1 × 6.4–9.6 (12.7 × 8.1; n = 15) | 13.5–18.6 × 9.7–16.0 (16.2 × 13.1; n = 17) | - |
| Beech marten | 3/4 (75.0%) | 24.9–98.7 | 7.0–12.6 × 7.0–8.6 (10.2 × 7.8; n = 10) | 13.5–23.9 × 10.1–17.5 (16.2 × 13.1; n = 23) | - |
| Overall | 61/115 (53.0%) | 43.9–62.2 | 7.0–14.6 × 6.4–10.2 (12.1 × 8.2; n = 450) | 12.4–23.9 × 9.7–18.3 (16.5 × 13.2; n = 130) | 13.9–23.5 × 12.0–22.2 (20.6 × 17.4; n = 46) |
| Host Species | N | Sarcocystis rileyi (%, 95% CI) | Sarcocystis sp. (%, 95% CI) |
|---|---|---|---|
| American mink | 61 | 8 (13.1, 6.2–24.4) | 1 (1.6, 0.9–8.7) |
| European pine marten | 26 | 0 | 1 (3.8, 0.2–18.8) |
| European polecat | 18 | 2 (11.1, 20.0–33.0) | 0 |
| European badger | 6 | 1 (16.7, 8.6–58.9) | 0 |
| Beech marten | 4 | 0 | 0 |
| Overall | 115 | 11 (9.6, 5.1–16.4) | 2 (3.3, 0.3–6.3) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prakas, P.; Moskaliova, D.; Šneideris, D.; Juozaitytė-Ngugu, E.; Maziliauskaitė, E.; Švažas, S.; Butkauskas, D. Molecular Identification of Sarcocystis rileyi and Sarcocystis sp. (Closely Related to Sarcocystis wenzeli) in Intestines of Mustelids from Lithuania. Animals 2023, 13, 467. https://doi.org/10.3390/ani13030467
Prakas P, Moskaliova D, Šneideris D, Juozaitytė-Ngugu E, Maziliauskaitė E, Švažas S, Butkauskas D. Molecular Identification of Sarcocystis rileyi and Sarcocystis sp. (Closely Related to Sarcocystis wenzeli) in Intestines of Mustelids from Lithuania. Animals. 2023; 13(3):467. https://doi.org/10.3390/ani13030467
Chicago/Turabian StylePrakas, Petras, Darija Moskaliova, Donatas Šneideris, Evelina Juozaitytė-Ngugu, Evelina Maziliauskaitė, Saulius Švažas, and Dalius Butkauskas. 2023. "Molecular Identification of Sarcocystis rileyi and Sarcocystis sp. (Closely Related to Sarcocystis wenzeli) in Intestines of Mustelids from Lithuania" Animals 13, no. 3: 467. https://doi.org/10.3390/ani13030467
APA StylePrakas, P., Moskaliova, D., Šneideris, D., Juozaitytė-Ngugu, E., Maziliauskaitė, E., Švažas, S., & Butkauskas, D. (2023). Molecular Identification of Sarcocystis rileyi and Sarcocystis sp. (Closely Related to Sarcocystis wenzeli) in Intestines of Mustelids from Lithuania. Animals, 13(3), 467. https://doi.org/10.3390/ani13030467