

A Novel Approach in the Development of Larval Largemouth Bass Micropterus salmoides Diets Using Largemouth Bass Muscle Hydrolysates as the Protein Source
 , ,
, ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Conditions
2.2. Muscle Hydrolysis
- -
- 1.5 h: 1 h at pH 3–4, followed by 30 min at pH 7–9;
- -
- 3 h: 1 h at pH 3–4, followed by 2 h at pH 7–9;
- -
- 6 h: 1 h at pH 3–4, followed by 5 h at pH 7–9.
2.3. Diets
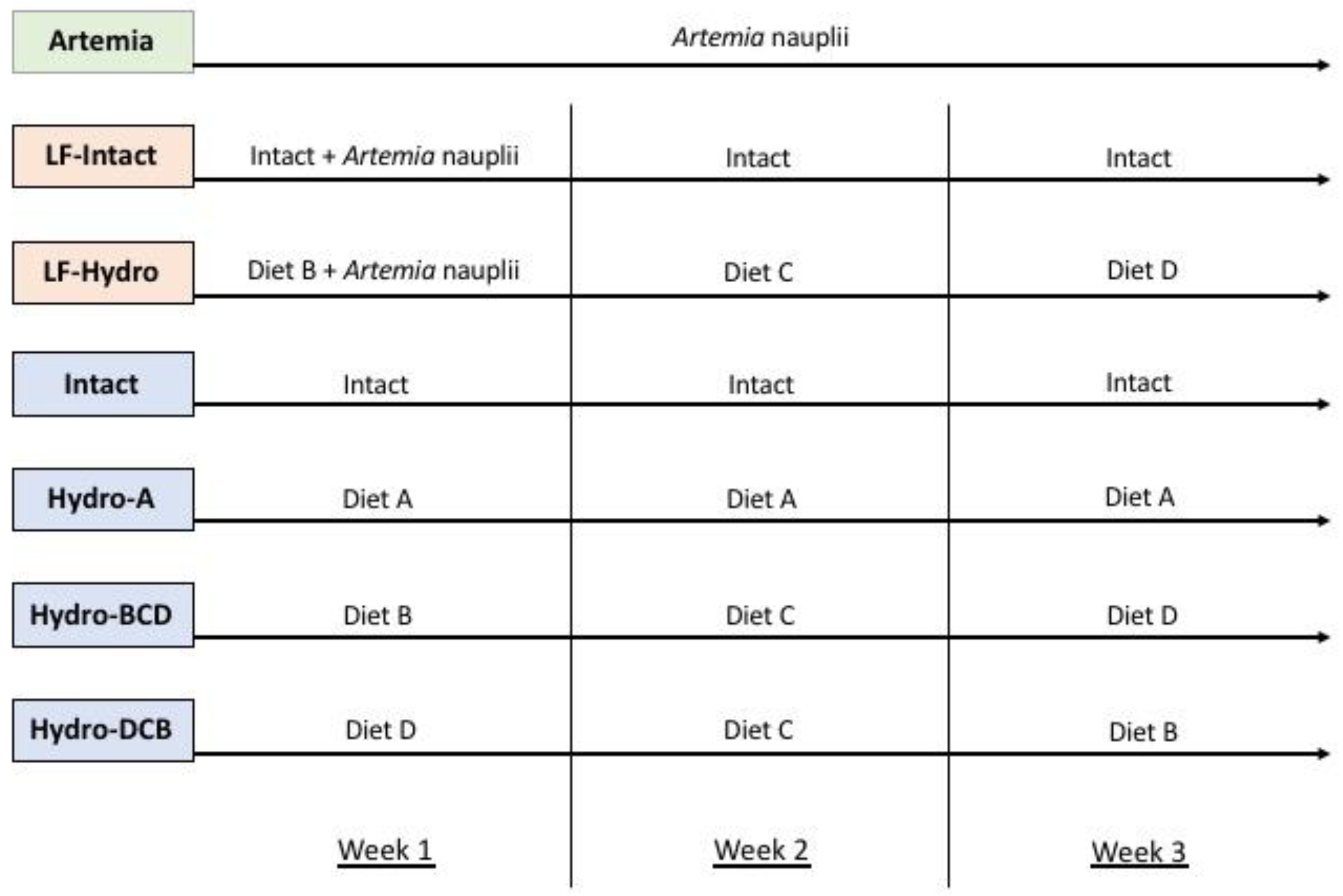
2.4. Experimental Design
2.5. Sampling and Measuring
2.6. Free Amino Acid Analysis
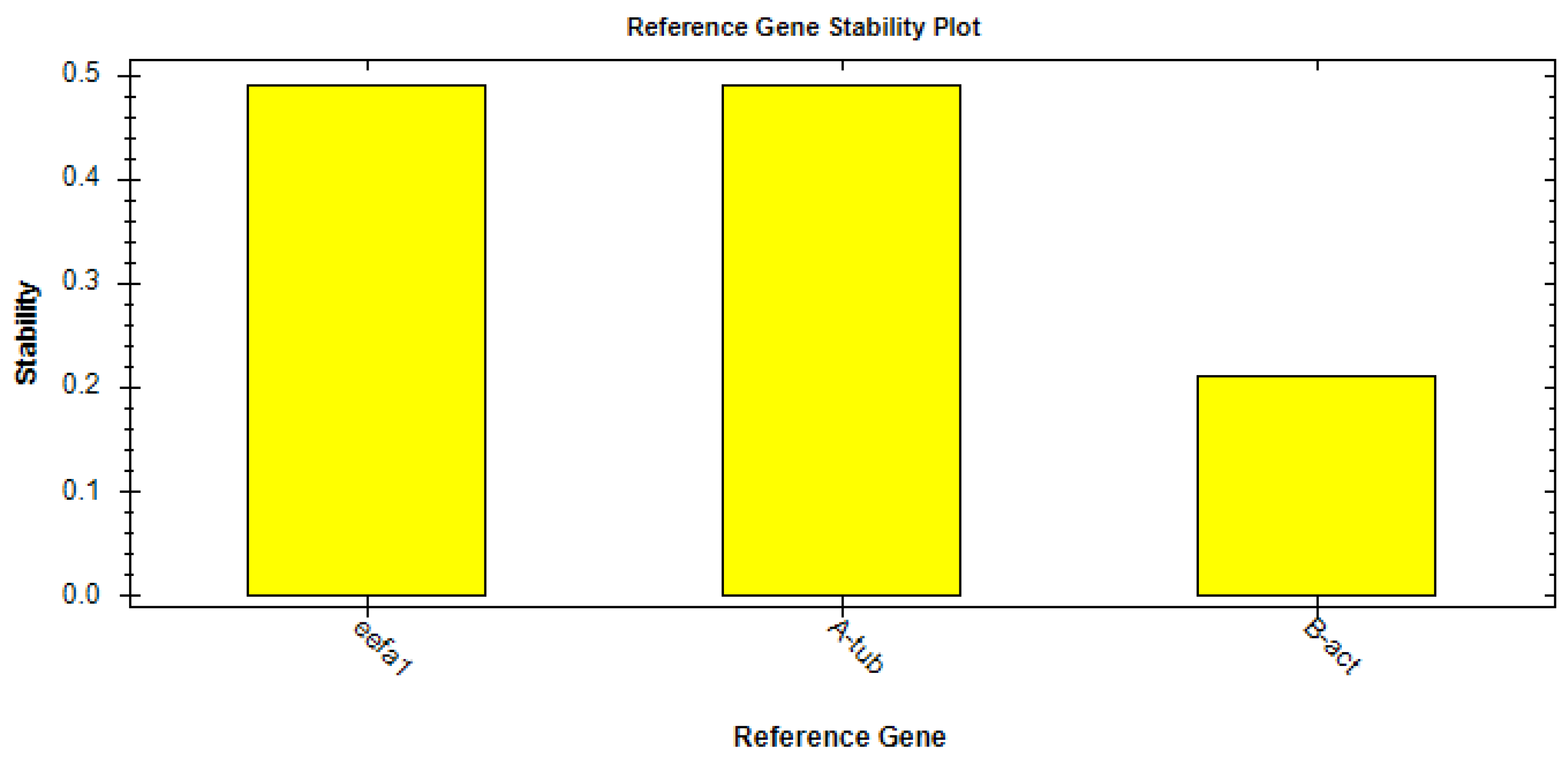
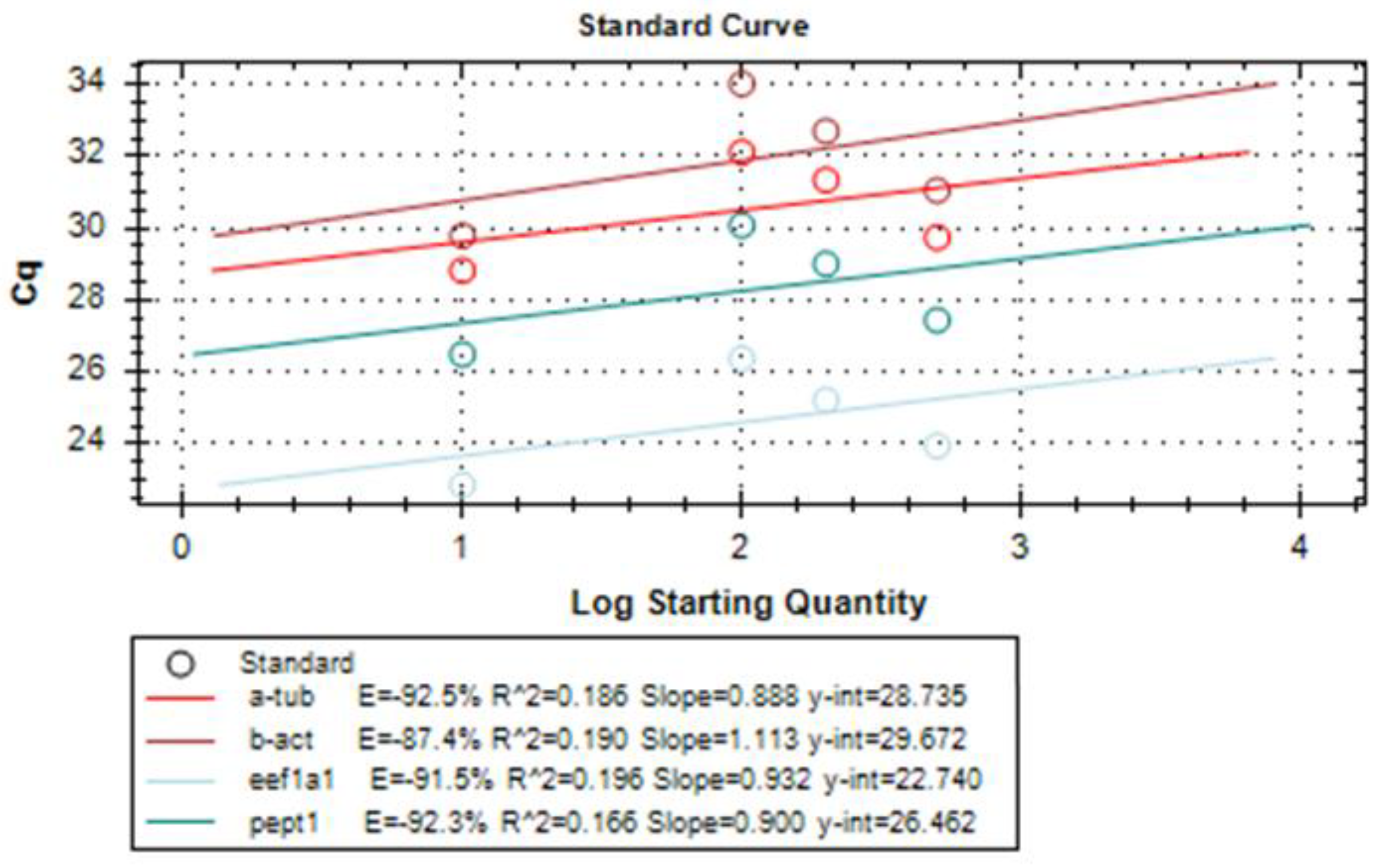
2.7. Gene Expression
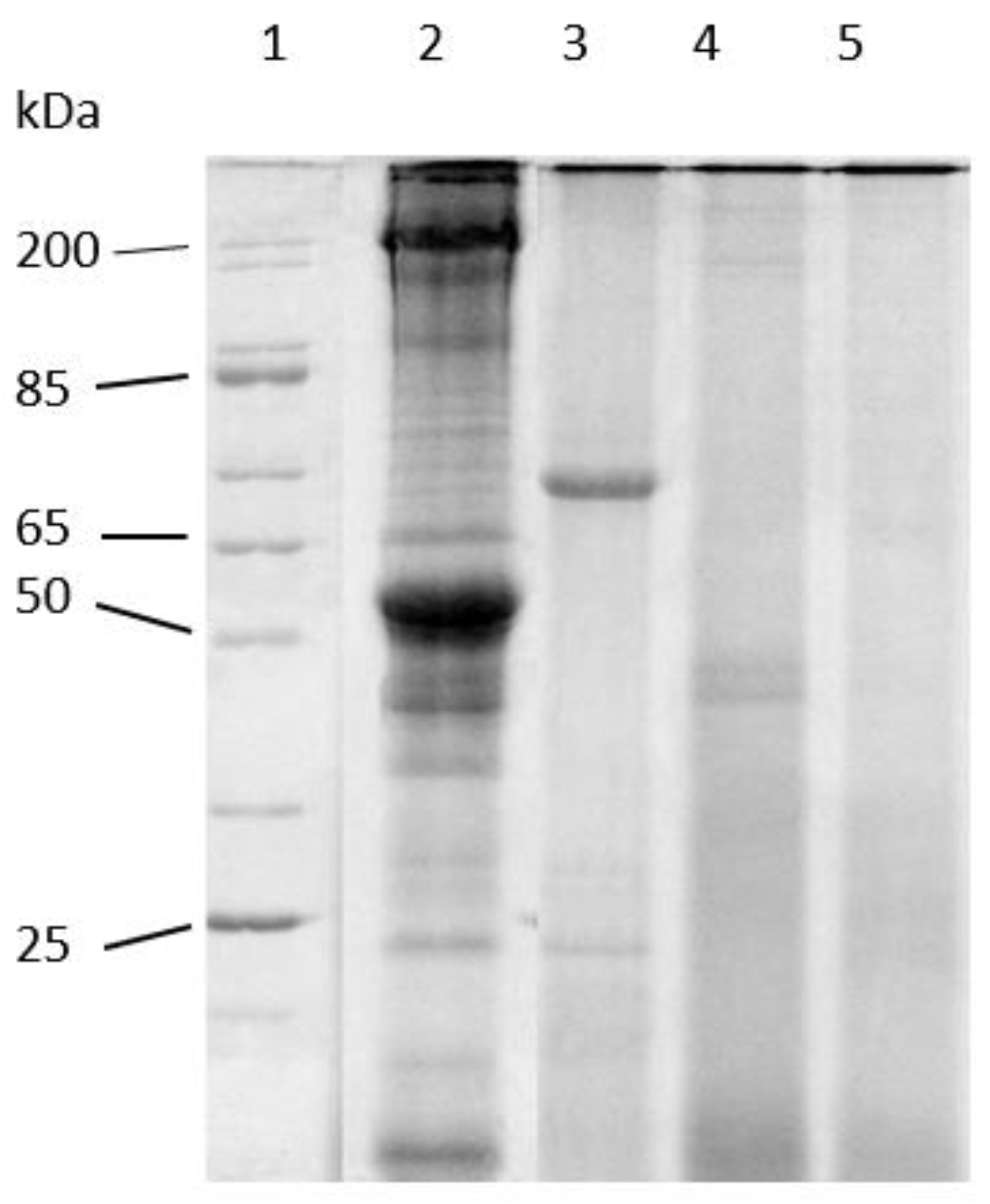
2.8. SDS-PAGE
2.9. Statistical Analysis
3. Results
3.1. LMB Muscle Hydrolysates
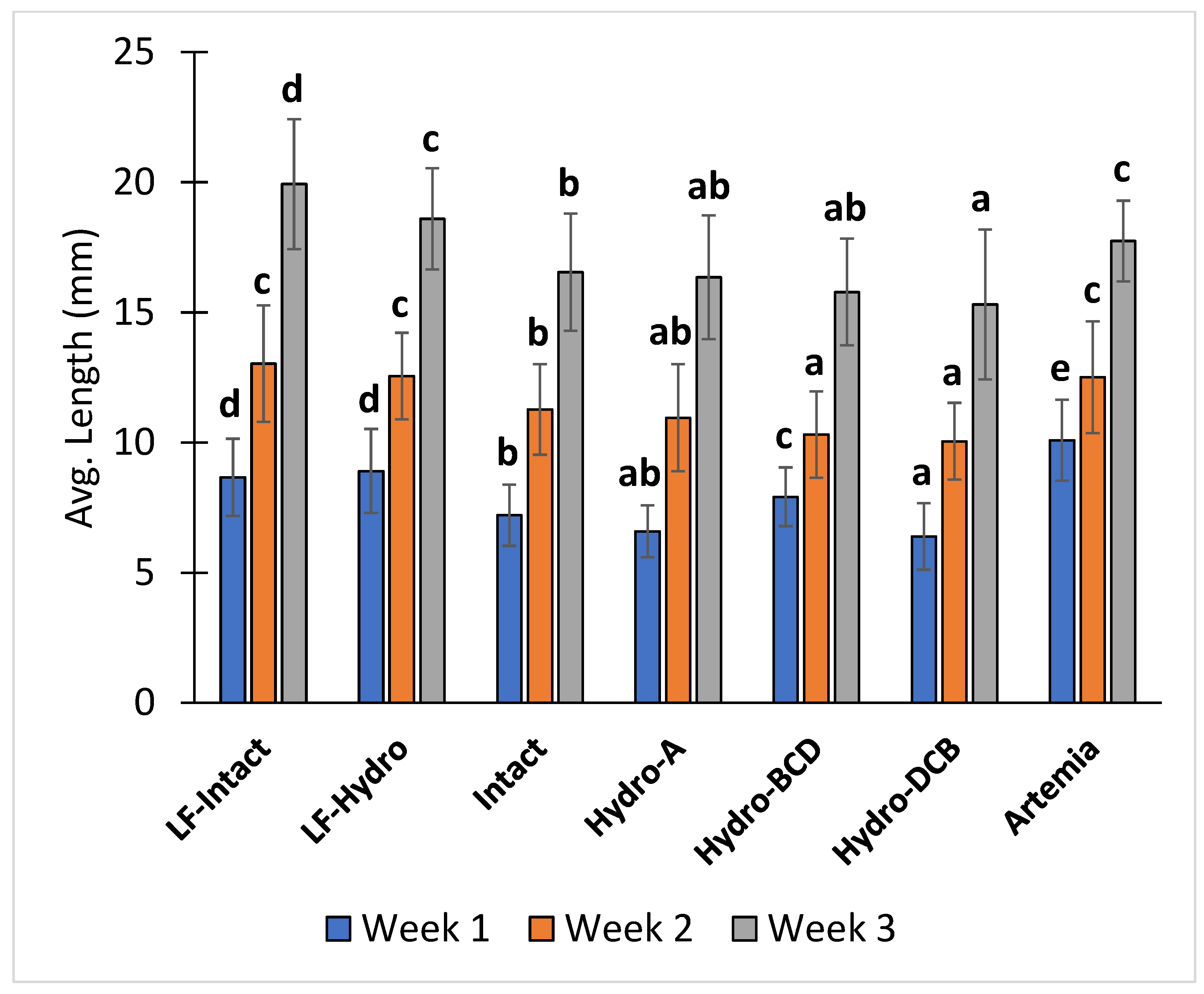
3.2. Growth and Survival
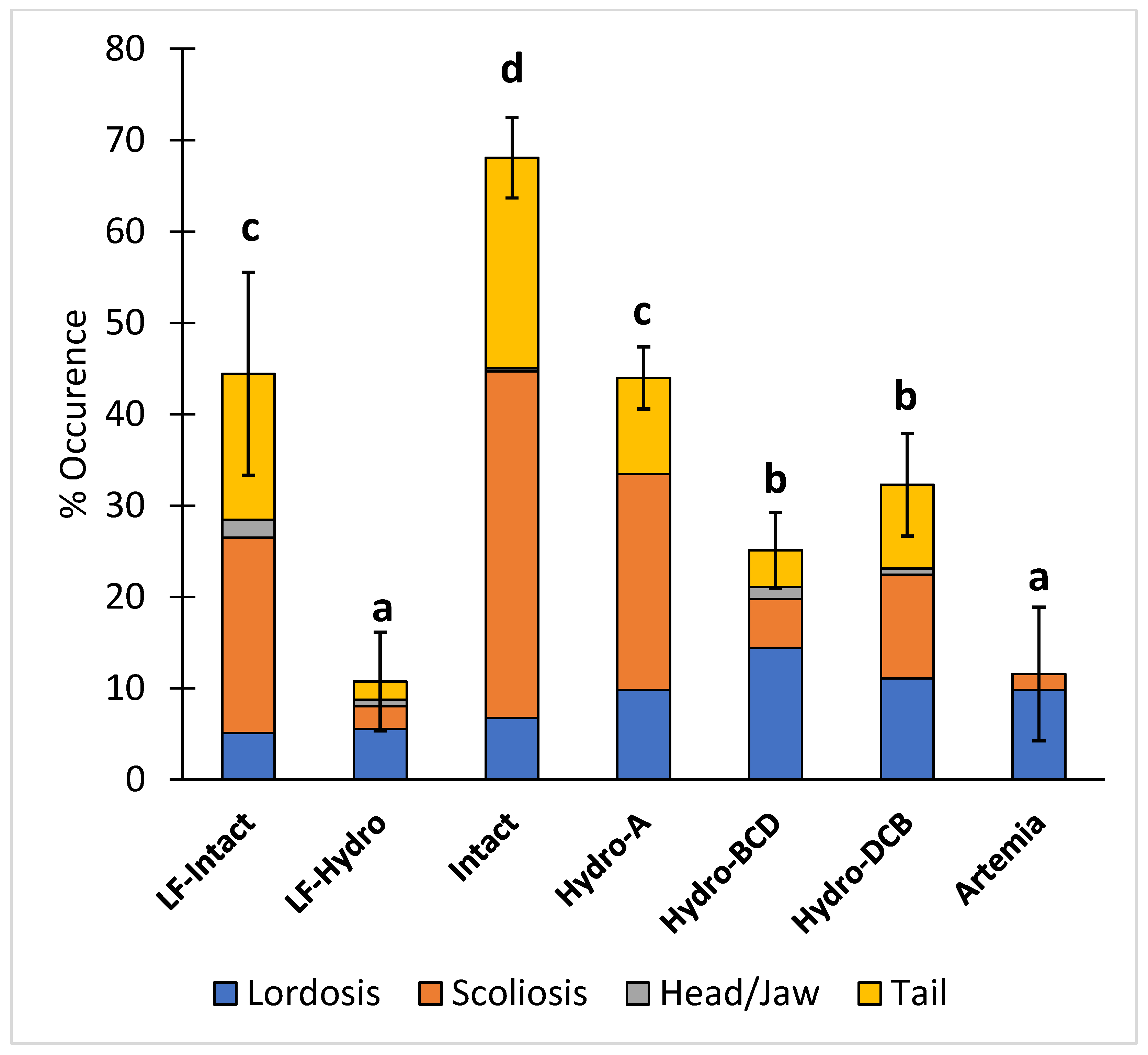
3.3. Skeletal Deformities
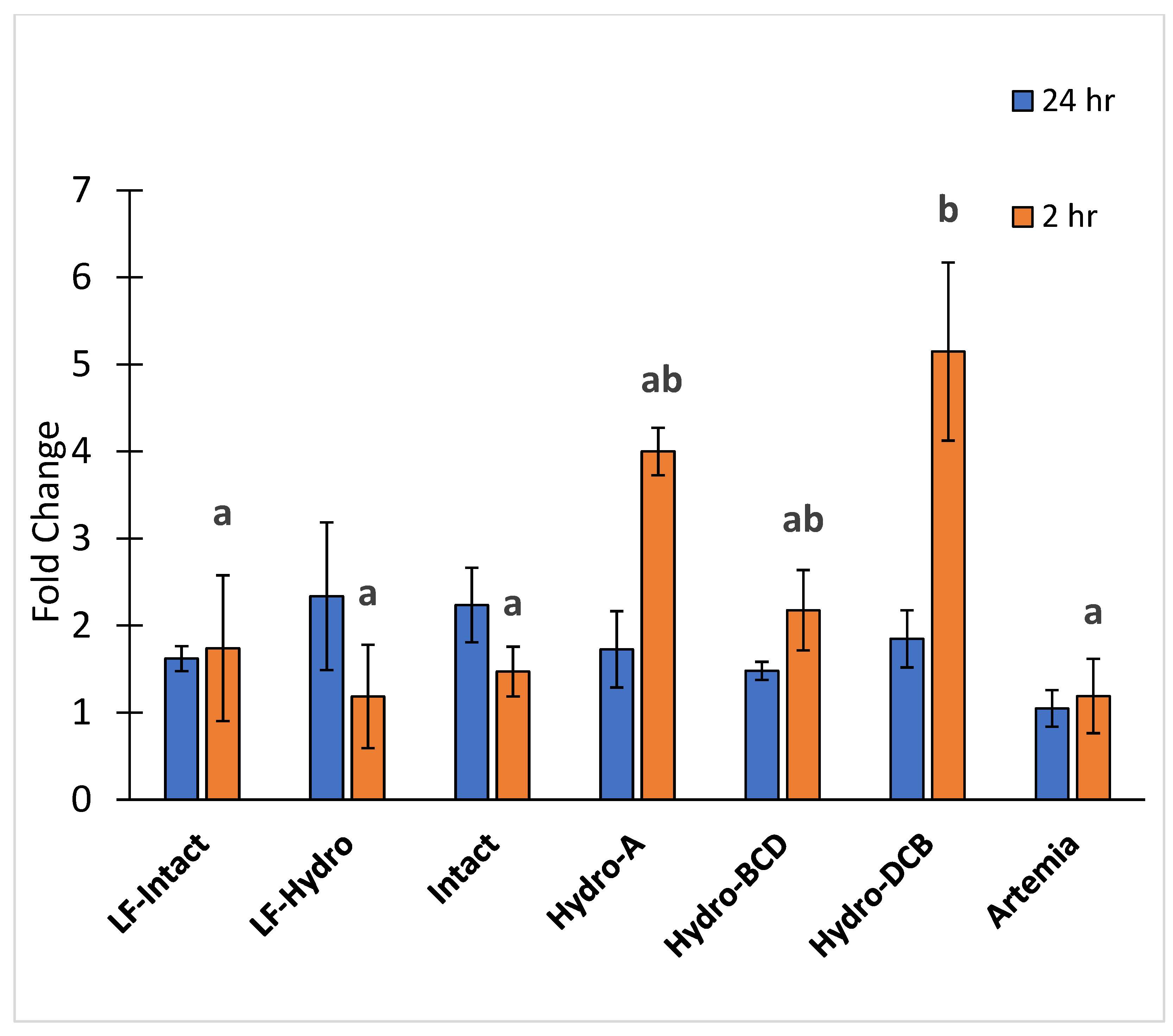
3.4. PepT1 Expression
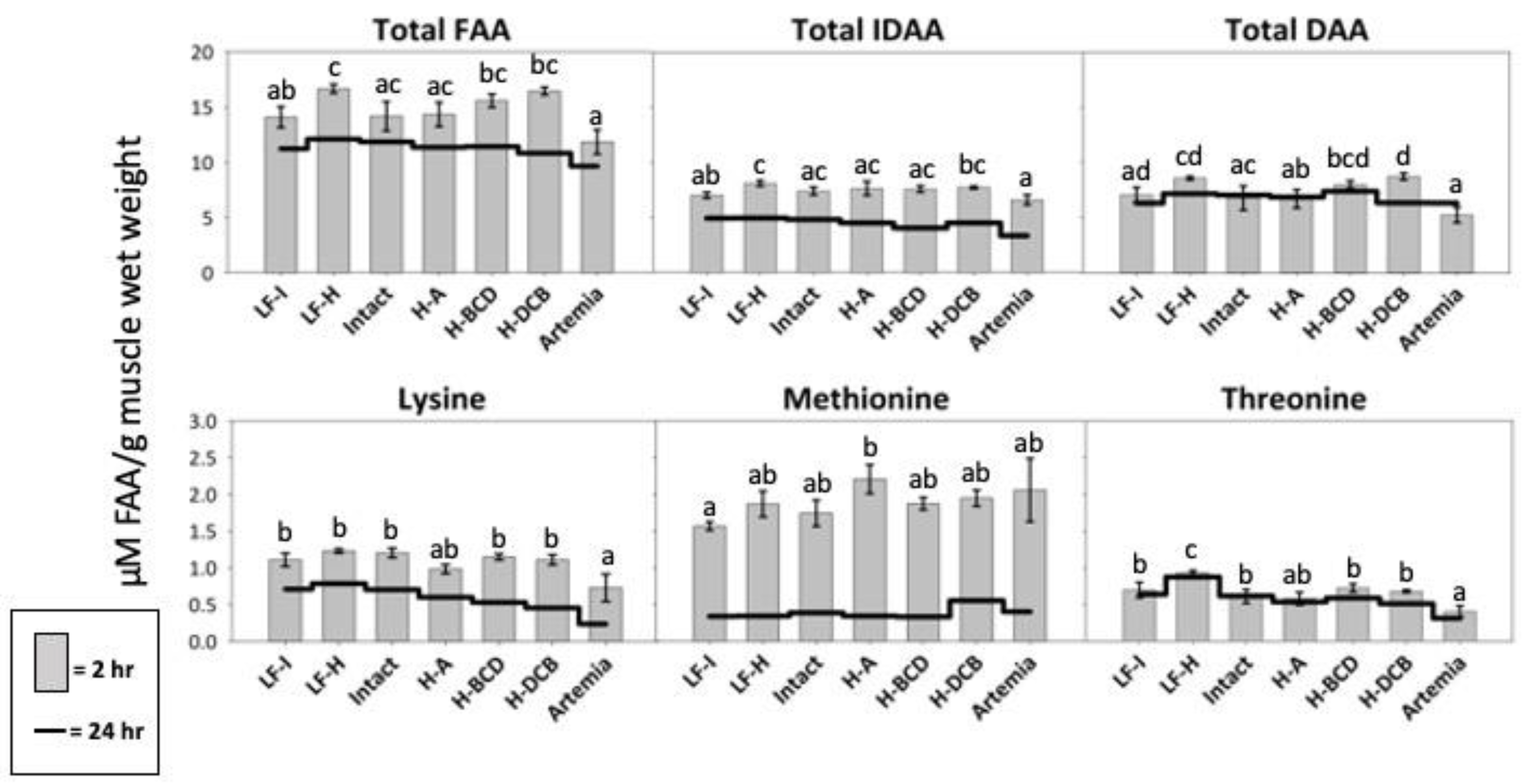
3.5. Free Amino Acid Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Samat, N.A.; Yusoff, F.M.; Rasdi, N.W.; Karim, M. Enhancement of Live Food Nutritional Status with Essential Nutrients for Improving Aquatic Animal Health: A Review. Animals 2020, 10, 2457. [Google Scholar] [CrossRef] [PubMed]
- Cahu, C.; Infante, J.Z. Substitution of live food by formulated diets in marine fish larvae. Aquaculture 2001, 200, 161–180. [Google Scholar] [CrossRef]
- Callan, C.; Jordaan, A.; Kling, L.J. Reducing Artemia use in the culture of Atlantic cod (Gadus morhua). Aquaculture 2003, 219, 585–595. [Google Scholar] [CrossRef]
- Fletcher, R.C., Jr.; Roy, W.; Davie, A.; Taylor, J.; Robertson, D.; Migaud, H. Evaluation of new microparticulate diets for early weaning of Atlantic cod (Gadus morhua): Implications on larval performances and tank hygiene. Aquaculture 2007, 263, 35–51. [Google Scholar] [CrossRef]
- Conceição, L.E.; Yúfera, M.; Makridis, P.; Morais, S.; Dinis, M.T. Live foods for early stages of fish rearing. Aquac. Res. 2010, 41, 613–640. [Google Scholar] [CrossRef]
- Kandathil Radhakrishnan, D.; AkbarAli, I.; Schmidt, B.V.; John, E.M.; Sivanpillai, S.; Thazhakot Vasunambesan, S. Improvement of nutritional quality of live food for aquaculture: An overview. Aquac. Res. 2020, 51, 1–17. [Google Scholar] [CrossRef]
- Walford, J.; Lim, T.M.; Lam, T.J. Replacing live foods with microencapsulated diets in the rearing of seabass (Lates calcarifer) larvae: Do the larvae ingest and digest protein-membrane microcapsules? Aquaculture 1991, 92, 225–235. [Google Scholar] [CrossRef]
- People Le Ruyet, J.; Alexandre, J.C.; Thébaud, L.; Mugnier, C. Marine fish larvae feeding: Formulated diets or live prey? J. World Aquac. Soc. 1993, 24, 211–224. [Google Scholar] [CrossRef]
- Lazo, J.P.; Holt, G.J.; Arnold, C.R. Ontogeny of pancreatic enzymes in larval red drum Sciaenops ocellatus. Aquac. Nutr. 2000, 6, 183–192. [Google Scholar] [CrossRef]
- Sarvi, B.; Matinfar, A.; Eskandary, G.R.; Sethi, U.S. Substitution of rotifer by a microencapsulated diet in yellow fin seabream (Acanthopagrus ltus) larviculture. Iran. Sci. Fish. J. 2009, 18, 53–66. [Google Scholar]
- Khoa, T.N.D.; Waqalevu, V.; Honda, A.; Shiozaki, K.; Kotani, T. Comparative study on early digestive enzyme activity and expression in red sea bream (Pagrus major) fed on live feed and micro-diet. Aquaculture 2020, 519, 734721. [Google Scholar] [CrossRef]
- Mozanzadeh, M.T.; Bahabadi, M.N.; Morshedi, V.; Azodi, M.; Agh, N.; Gisbert, E. Weaning strategies affect larval performance in yellowfin seabream (Acanthopagrus latus). Aquaculture 2021, 539, 736673. [Google Scholar] [CrossRef]
- Goolish, E.M.; Okutake, K.; Lesure, S. Growth and survivorship of larval zebrafish Danio rerio on processed diets. N. A. J. Aquac. 1999, 61, 189–198. [Google Scholar] [CrossRef]
- Adekunle, A.I.; Joyce, A.O.B. Effect of partial and total replacement of live food (artemia) with formulated diets in early stage growth of hybrid catfish (Heterobranchus bidorsalis x Heterobranchus longifilis) fry. Int. J. Fish. Aquat. Studies 2014, 1, 143–146. [Google Scholar]
- Jamali, H.; Meshkini, S. Study of Sudden Replacement of Live Feed by Commercial Feed Affects Growth, Survival Rate and Resistance to Environmental Stress in Oscar Fish (Astronotus ocellatus) Larvae. Fish. Sci. Technol. 2018, 7, 279–286. [Google Scholar]
- Lipscomb, T.N.; Patterson, J.T.; Wood, A.L.; Watson, C.A.; DiMaggio, M.A. Larval growth, survival, and partial budget analysis related to replacing Artemia in larval culture of six freshwater ornamental fishes. J. World Aquac. Soc. 2020, 51, 1132–1144. [Google Scholar] [CrossRef]
- Cahu, C.; Infante, J.Z.; Takeuchi, T. Nutritional components affecting skeletal development in fish larvae. Aquaculture 2003, 227, 245–258. [Google Scholar] [CrossRef]
- Mambrini, M.; Kaushik, S.J. Indispensable amino acid requirements of fish: Correspondence between quantitative data and amino acid profiles of tissue proteins. J. Appl. Ichthyol. 1995, 11, 240–247. [Google Scholar] [CrossRef]
- Akiyama, T.; Oohara, I.; Yamamoto, T. Comparison of essential amino acid requirements with A/E ratio among fish species. Fish. Sci. 1997, 63, 963–970. [Google Scholar] [CrossRef]
- Meyer, G.; Fracalossi, D.M. Estimation of jundiá (Rhamdia quelen) dietary amino acid requirements based on muscle amino acid composition. Sci. Agric. 2005, 62, 401–405. [Google Scholar] [CrossRef]
- Cole, D.J.A. The amino acid requirements of pigs-the concept of an ideal protein. Pig News Inf. 1980, 1, 201–205. [Google Scholar]
- Wilson, R.P.; Cowey, C.B. Amino acid composition of whole body tissue of rainbow trout and Atlantic salmon. Aquaculture 1985, 48, 373–376. [Google Scholar] [CrossRef]
- Dabrowski, K. The feeding of fish larvae: Present « state of the art» and perspectives. Reprod. Nutr. Développement 1984, 24, 807–833. [Google Scholar] [CrossRef]
- Kolkovski, S. Digestive enzymes in fish larvae and juveniles—Implications and applications to formulated diets. Aquaculture 2001, 200, 181–201. [Google Scholar] [CrossRef]
- Zambonino Infante, J.L.; Cahu, C.L.; Peres, A. Partial substitution of di-and tripeptides for native proteins in sea bass diet improves Dicentrarchus labrax larval development. J. Nutr. 1997, 127, 608–614. [Google Scholar] [CrossRef]
- Fernández, I.; Hontoria, F.; Ortiz-Delgado, J.B.; Kotzamanis, Y.; Estévez, A.; Zambonino-Infante, J.L.; Gisbert, E. Larval performance and skeletal deformities in farmed gilthead sea bream (Sparus aurata) fed with graded levels of Vitamin A enriched rotifers (Brachionus plicatilis). Aquaculture 2008, 283, 102–115. [Google Scholar] [CrossRef]
- Kwasek, K.; Gonzalez, C.; Wick, M.; Molinari, G.S.; Wojno, M. Fish muscle hydrolysate obtained using largemouth bass Micropterus salmoides digestive enzymes improves largemouth bass performance in its larval stages. PLoS ONE 2021, 16, e0261847. [Google Scholar] [CrossRef]
- Kotzamanis, Y.P.; Gisbert, E.; Gatesoupe, F.J.; Infante, J.Z.; Cahu, C. Effects of different dietary levels of fish protein hydrolysates on growth, digestive enzymes, gut microbiota, and resistance to Vibrio anguillarum in European sea bass (Dicentrarchus labrax) larvae. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 205–214. [Google Scholar] [CrossRef]
- Swanepoel, J.C.; Goosen, N.J. Evaluation of fish protein hydrolysates in juvenile African catfish (Clarias gariepinus) diets. Aquaculture 2018, 496, 262–269. [Google Scholar] [CrossRef]
- Srichanun, M.; Tantikitti, C.; Kortner, T.M.; Krogdahl, Å.; Chotikachinda, R. Effects of different protein hydrolysate products and levels on growth, survival rate and digestive capacity in Asian seabass (Lates calcarifer Bloch) larvae. Aquaculture 2014, 428, 195–202. [Google Scholar] [CrossRef]
- Canada, P.; Conceição, L.E.; Mira, S.; Teodósio, R.; Fernandes, J.M.; Barrios, C.; Millán, F.; Pedroche, J.; Valente, L.M.; Engrola, S. Dietary protein complexity modulates growth, protein utilisation and the expression of protein digestion-related genes in Senegalese sole larvae. Aquaculture 2017, 479, 273–284. [Google Scholar] [CrossRef]
- Dabrowski, K.; Miller, M. Contested paradigm in raising zebrafish (Danio rerio). Zebrafish 2018, 15, 295–309. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Nutrient Requirements of Fish and Shrimp; National Academies Press: Washington, DC, USA, 2011. [Google Scholar]
- Terova, G.; Corà, S.; Verri, T.; Rimoldi, S.; Bernardini, G.; Saroglia, M. Impact of feed availability on PepT1 mRNA expression levels in sea bass (Dicentrarchus labrax). Aquaculture 2009, 294, 288–299. [Google Scholar] [CrossRef]
- Updike, M.S.; Zerby, H.; Utrata, K.L.; Lilburn, M.; Kaletunc, G.; Wick, M. Proteins associated with thermally induced gelation of turkey breast meat. J. Food Sci. 2006, 71, E398–E402. [Google Scholar] [CrossRef]
- Sheng, Z.; Turchini, G.M.; Xu, J.; Fang, Z.; Chen, N.; Xie, R.; Zhang, H.; Li, S. Functional properties of protein hydrolysates on growth, digestive enzyme activities, protein metabolism, and intestinal health of larval largemouth bass (Micropterus salmoides). Front. Immunol. 2022, 13, 3517. [Google Scholar] [CrossRef]
- Canavate, J.P.; Fernández-Dıaz, C. Influence of co-feeding larvae with live and inert diets on weaning the sole Solea senegalensis onto commercial dry feeds. Aquaculture 1999, 174, 255–263. [Google Scholar] [CrossRef]
- Rasdi, N.W.; Qin, J.G. Improvement of copepod nutritional quality as live food for aquaculture: A review. Aquac. Res. 2016, 47, 1–20. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Sá, R.; Oliva-Teles, A.; Bergot, P. Solubility and peptide profile affect the utilization of dietary protein by common carp (Cyprinus carpio) during early larval stages. Aquaculture 2004, 234, 319–333. [Google Scholar] [CrossRef]
- Önal, U.; Langdon, C. Potential delivery of water-soluble protein hydrolysates to marine suspension feeders by three different microbound particle types. Aquaculture 2009, 296, 174–178. [Google Scholar] [CrossRef]
- Martínez-Alvarez, O.; Chamorro, S.; Brenes, A. Protein hydrolysates from animal processing by-products as a source of bioactive molecules with interest in animal feeding: A review. Food Res. Int. 2015, 73, 204–212. [Google Scholar] [CrossRef]
- de Vareilles, M.; Richard, N.; Gavaia, P.J.; Silva, T.S.; Cordeiro, O.; Guerreiro, I.; Yúfera, M.; Batista, I.; Pires, C.; Pousão-Ferreira, P.; et al. Impact of dietary protein hydrolysates on skeleton quality and proteome in Diplodus sargus larvae. J. Appl. Ichthyol. 2012, 28, 477–487. [Google Scholar] [CrossRef]
- Rønnestad, I.; Conceição, L.E.; Aragão, C.; Dinis, M.T. Free amino acids are absorbed faster and assimilated more efficiently than protein in postlarval Senegal sole (Solea senegalensis). J. Nutr. 2000, 130, 2809–2812. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dabrowski, K.; Hliwa, P.; Gomulka, P. Indispensable amino acid concentrations decrease in tissues of stomachless fish, common carp in response to free amino acid-or peptide-based diets. Amino Acids 2006, 31, 165–172. [Google Scholar] [CrossRef]
- Ambardekar, A.A.; Reigh, R.C.; Williams, M.B. Absorption of amino acids from intact dietary proteins and purified amino acid supplements follows different time-courses in channel catfish (Ictalurus punctatus). Aquaculture 2009, 291, 179–187. [Google Scholar] [CrossRef]
- Roy, P.K.; Lall, S.P. Vitamin K deficiency inhibits mineralization and enhances deformity in vertebrae of haddock (Melanogrammus aeglefinus L.). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2007, 148, 174–183. [Google Scholar] [CrossRef]
- Kanazawa, A.; Teshima, S.; Inamori, S.; Iwashita, T.; Nagao, A. Effects of phospholipids [in diets] on growth, survival rate, and incidence of malformation in the larval ayu [Plecoglossus altivelis]. Mem. Fac. Fish. -Kagoshima Univ. (Jpn.) 1982, 30, 301–309. [Google Scholar]
- Akiyama, T.; Murai, T.; Nose, T. Oral administration of serotonin against spinal deformity of chum salmon [Onchorhynchus keta] fry induced by tryptophan deficiency. Bull. Jpn. Soc. Sci. Fish. (Jpn.) 1986, 52, 1249–1254. [Google Scholar] [CrossRef]
- Boursiaki, V.; Theochari, C.; Zaoutsos, S.P.; Mente, E.; Vafidis, D.; Apostologamvrou, C.; Berillis, P. Skeletal deformity of scoliosis in gilthead seabreams (Sparus aurata): Association with changes to calcium-phosphor hydroxyapatite salts and collagen fibers. Water 2019, 11, 257. [Google Scholar] [CrossRef]
- Cahu, C.L.; Infante, J.Z.; Quazuguel, P.; Le Gall, M.M. Protein hydrolysate vs. fish meal in compound diets for 10-day old sea bass Dicentrarchus labrax larvae. Aquaculture 1999, 171, 109–119. [Google Scholar] [CrossRef]
- Johannsdottir, J.; Heimisdottir, H.L.; Hakonardottir, K.; Hrolfsdottir, L.; Steinarsson, A.; Imsland, A.K.; Thorarensen, H.; Bergsson, A.B.; Bjornsdottir, R. Improved performance of A tlantic cod (Gadus morhua L.) larvae following enhancement of live feed using a fish protein hydrolysate. Aquac. Nutr. 2014, 20, 314–323. [Google Scholar] [CrossRef]
- Peruzzi, S.; Puvanendran, V.; Riesen, G.; Seim, R.R.; Hagen, Ø.; Martinez-Llorens, S.; Falk-Petersen, I.B.; Fernandes, J.M.; Jobling, M. Growth and development of skeletal anomalies in diploid and triploid Atlantic salmon (Salmo salar) fed phosphorus-rich diets with fish meal and hydrolyzed fish protein. PLoS ONE 2018, 13, e0194340. [Google Scholar] [CrossRef] [PubMed]
- Kwasek, K.; Zhang, Y.; Dabrowski, K. Utilization of Dipeptide/Protein Based Diets in Larval and Juvenile Koi Carp Post-Prandial Free Amino Acid Levels. J. Anim. Physiol. Anim. Nutr. 2009, 94, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, S.J.; Luquet, P. Influence of bacterial protein incorporation and of sulphur amino acid supplementation to such diets on growth of rainbow trout, Salmo gairdnerii Richardson. Aquaculture 1980, 19, 163–175. [Google Scholar] [CrossRef]
- Mente, E.; Deguara, S.; Santos, M.B.; Houlihan, D. White muscle free amino acid concentrations following feeding a maize gluten dietary protein in Atlantic salmon (Salmo salar L.). Aquaculture 2003, 225, 133–147. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Intact | A | B | C | D |
|---|---|---|---|---|---|
| Fish meal (Intact) | 74.00 | 37.00 | 37.00 | 37.00 | 37.00 |
| Hydrolysate 1.5 h | - | 12.30 | 3.70 | 7.40 | 22.20 |
| Hydrolysate 3 h | - | 12.30 | 11.10 | 22.20 | 11.10 |
| Hydrolysate 6 h | - | 12.30 | 22.20 | 7.40 | 3.70 |
| CPSP 90 1 | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 |
| Krill Meal 2 | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 |
| Fish oil 3 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 |
| Lecithin 4 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 |
| Mineral mix 5 | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 |
| Vitamin mix 6 | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 |
| CaHPO4 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Taurine | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Choline chloride | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| Vitamin C 7 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Proximate Comp. (%) | |||||
| Crude Protein (N × 6.25) | 60.97 (±0.08) | 60.99 (±0.17) | 60.96 (±0.20) | 61.02 (±0.03) | 60.96 (±0.19) |
| Lipids | 18.05 (±0.18) | 18.06 (±0.08) | 18.00 (±0.10) | 17.98 (±0.16) | 17.97 (±0.15) |
| Ash | 12.01 (±0.07) | 11.99 (±0.17) | 12.02 (±0.15) | 12.07 (±0.13) | 12.06 (±0.11) |
| Amino Acid Composition (%) | Intact | A | B | C | D |
|---|---|---|---|---|---|
| Alanine | 3.90 (±0.10) | 3.73 (±0.01) | 3.72 (±0.10) | 3.73 (±0.09) | 3.89 (±0.10) |
| Arginine | 3.54 (±0.08) | 3.38 (±0.02) | 3.41 (±0.08) | 3.38 (±0.08) | 3.49 (±0.06) |
| Aspartic Acid | 5.69 (±0.11) | 5.49 (±0.02) | 5.49 (±0.10) | 5.53 (±0.17) | 5.35 (±0.09) |
| Cysteine | 0.57 (±0.02) | 0.54 (±0.02) | 0.57 (±0.01) | 0.55 (±0.04) | 0.50 (±0.04) |
| Glutamic Acid | 8.02 (±0.19) | 7.76 (±0.06) | 7.64 (±0.20) | 7.70 (±0.16) | 7.84 (±0.10) |
| Glycine | 4.73 (±0.20) | 4.33 (±0.08) | 4.48 (±0.15) | 4.45 (±0.15) | 4.70 (±0.18) |
| Histidine | 1.35 (±0.02) | 1.35 (±0.02) | 1.32 (±0.02) | 1.32 (±0.05) | 1.26 (0.05) |
| Hydroxyproline | 0.87 (±0.07) | 0.74 (±0.05) | 0.83 (±0.04) | 0.79 (±0.18) | 1.13 (±0.20) |
| Isoleucine | 2.65 (±0.05) | 2.62 (±0.03) | 2.56 (±0.05) | 2.55 (±0.11) | 2.43 (±0.10) |
| Leucine | 4.29 (±0.09) | 4.32 (±0.09) | 4.15 (±0.08) | 4.18 (±0.15) | 3.99 (±0.17) |
| Lysine | 4.81 (±0.10) | 4.85 (±0.11) | 4.65 (±0.09) | 4.65 (±0.18) | 4.45 (±0.20) |
| Methionine | 1.57 (±0.04) | 1.51 (±0.01) | 1.49 (±0.04) | 1.52 (±0.05) | 1.47 (±0.02) |
| Phenylalanine | 2.27 (±0.04) | 2.26 (±0.04) | 2.19 (±0.04) | 2.21 (±0.05) | 2.13 (±0.02) |
| Proline | 2.71 (±0.14) | 2.42 (±0.07) | 2.55 (±0.09) | 2.54 (±0.17) | 2.89 (±0.24) |
| Serine | 2.12 (±0.04) | 2.04 (±0.03) | 2.07 (±0.03) | 2.10 (±0.04) | 2.05 (±0.01) |
| Taurine | 1.95 (±0.27) | 2.50 (±0.17) | 2.19 (±0.15) | 2.23 (±0.14) | 2.06 (±0.23) |
| Threonine | 2.49 (±0.05) | 2.40 (±0.03) | 2.43 (±0.03) | 2.46 (±0.07) | 2.36 (±0.03) |
| Tryptophan | 0.62 (±0.08) | 0.54 (±0.05) | 0.45 (±0.08) | 0.52 (±0.08) | 0.46 (±0.05) |
| Tyrosine | 1.79 (±0.06) | 1.86 (±0.06) | 1.73 (±0.03) | 1.77 (±0.06) | 1.68 (±0.09) |
| Valine | 2.98 (±0.05) | 2.94 (±0.03) | 2.88 (±0.05) | 2.90 (±0.09) | 2.81 (±0.07) |
| Primer | Nucleotide Sequence (5′-3′) | Amplicon Size (Base Pairs) | |
|---|---|---|---|
| PepT1 | Forward | AAGCTGGGACCGAGAAGATT | 120 |
| Reverse | CGGCCAATAAAGTGGTTTCA | ||
| eEF1a1 | Forward | GTTGCTGCTGGTGTTGGTGAG | 156 |
| Reverse | GAAACGCTTCTGGCTGTAAGG | ||
| α-tubulin | Forward | GACTCGACCACAAGTTTGACC | 142 |
| Reverse | GTTCCCACCTCTTCGTAATCC | ||
| β-actin | Forward | GTATTGTCATGGACTCTGGTG | 182 |
| Reverse | ACGTACGATTTCACGCTCAGC |
| Group | Avg. Weight (mg) | Avg. Total Length (mm) | Survival (%) |
|---|---|---|---|
| LF-Intact | 79.50 c (±30.00) | 19.93 d (±2.49) | 46.29 c (±9.20) |
| LF-Hydro | 70.67 bc (±23.71) | 18.60 c (±1.95) | 33.78 b (±6.72) |
| Intact | 51.67 ac (±14.74) | 16.55 b (±2.25) | 23.94 b (±8.69) |
| Hydro-A | 44.33 ab (±4.04) | 16.35 ab (±2.38) | 11.69 a (±2.11) |
| Hydro-BCD | 35.83 a (±6.37) | 15.78 ab (±2.05) | 11.33 a (±4.89) |
| Hydro-DCB | 41.00 a (±7.81) | 15.31 a (±2.88) | 9.96 a (±2.85) |
| Artemia | 72.32 bc (±10.98) | 17.74 c (±1.55) | 86.89 d (±1.81) |
| Amino Acid (μM/g) | LF-Intact | LF-Hydro | Intact | Hydro-A | Hydro-BCD | Hydro-DCB | Artemia |
|---|---|---|---|---|---|---|---|
| Aspartic Acid | 0.64 ab (±0.02) | 0.65 ab (±0.01) | 0.61 ab (±0.06) | 0.61 ab (±0.05) | 0.63 ab (±0.01) | 0.70 b (±0.05) | 0.53 a (±0.08) |
| Glutamic Acid | 1.56 bc (±0.08) | 1.63 c (±0.05) | 1.41 ac (±0.05) | 1.14 a (±0.11) | 1.40 ac (±0.07) | 1.35 ac (±0.03) | 1.25 ab (±0.29) |
| Asparagine | 0.38 a (±0.04) | 0.50 ab (±0.02) | 0.54 ab (±0.08) | 0.46 ab (±0.09) | 0.53 ab (±0.04) | 0.57 b (±0.08) | 0.45 ab (±0.05) |
| Serine | 0.55 a (±0.11) | 0.74 ab (±0.10) | 0.58 a (±0.13) | 0.60 a (±0.12) | 0.88 b (±0.07) | 0.93 b (±0.04) | 0.55 a (±0.07) |
| Glutamine | 0.63 ab (±0.04) | 0.72 ab (±0.03) | 0.79 b (±0.05) | 0.53 a (±0.09) | 0.71 ab (±0.06) | 0.76 ab (±0.17) | 0.64 ab (±0.05) |
| Histidine | 0.73 b (±0.12) | 0.69 b (±0.05) | 0.61 b (±0.14) | 0.61 b (±0.01) | 0.56 b (±0.04) | 0.60 b (±0.01) | 0.28 a (±0.11) |
| Glycine | 1.91 bc (±0.36) | 2.55 c (±0.15) | 1.35 b (±0.44) | 1.31 ab (±0.22) | 1.69 b (±0.30) | 1.75 b (±0.03) | 0.61 a (±0.04) |
| Threonine | 0.70 b (±0.11) | 0.94 c (±0.03) | 0.61 b (±0.09) | 0.58 ab (±0.09) | 0.73 b (±0.06) | 0.68 b (±0.02) | 0.41 a (±0.08) |
| Arginine | 0.09 b (±0.01) | 0.12 c (±0.01) | 0.10 b (±0.00) | 0.09 b (±0.01) | 0.11 bc (±0.01) | 0.10 b (±0.01) | 0.04 a (±0.01) |
| Alanine | 0.83 abc (±0.19) | 1.06 bd (±0.10) | 0.82 ab (±0.23) | 1.00 bc (±0.13) | 1.22 cd (±0.03) | 1.41 d (±0.02) | 0.54 a (±0.13) |
| Tyrosine | 0.20 bc (±0.01) | 0.25 d (±0.01) | 0.19 bc (±0.02) | 0.17 b (±0.03) | 0.22 cd (±0.01) | 0.19 bc (±0.01) | 0.09 a (±0.02) |
| Lysine | 1.11 b (±0.09) | 1.24 b (±0.03) | 1.21 b (±0.07) | 0.98 ab (±0.06) | 1.16 b (±0.04) | 1.12 b (±0.07) | 0.73 a (±0.19) |
| Methionine | 1.57 a (±0.06) | 1.87 ab (±0.18) | 1.74 ab (±0.18) | 2.21 b (±0.20) | 1.87 ab (±0.08) | 1.95 ab (±0.11) | 2.06 ab (±0.43) |
| Valine | 1.95 (±0.07) | 2.32 (±0.15) | 2.01 (±0.05) | 2.34 (±0.52) | 1.99 (±0.09) | 2.29 (±0.10) | 2.41 (±0.13) |
| Cysteine | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Tryptophan | 0.12 ab (±0.03) | 0.11 a (±0.01) | 0.12 ab (±0.00) | 0.08 a (±0.01) | 0.16 b (±0.03) | 0.12 ab (±0.00) | 0.10 a (±0.01) |
| Phenylalanine | 0.28 b (±0.02) | 0.30 b (±0.02) | 0.33 bc (±0.02) | 0.28 b (±0.01) | 0.39 c (±0.04) | 0.33 b (±0.01) | 0.19 a (±0.02) |
| Isoleucine | 0.24 (±0.17) | 0.15 (±0.03) | 0.34 (±0.22) | 0.11 (±0.03) | 0.19 (±0.04) | 0.14 (±0.03) | 0.16 (±0.02) |
| Leucine | 0.24 (±0.14) | 0.38 (±0.05) | 0.33 (±0.21) | 0.35 (±0.05) | 0.44 (±0.05) | 0.40 (±0.04) | 0.25 (±0.04) |
| Proline | 0.35 a (±0.02) | 0.47 ab (±0.07) | 0.49 ab (±0.17) | 0.87 cd (±0.03) | 0.71 bc (±0.08) | 1.07 d (±0.04) | 0.58 ab (±0.12) |
| IDAA | 7.03 ab (±0.28) | 8.11 c (±0.26) | 7.40 ac (±0.36) | 7.65 ac (±0.63) | 7.60 ac (±0.29) | 7.74 bc (±0.13) | 6.62 a (±0.45) |
| DAA | 7.06 ad (±0.67) | 8.57 cd (±0.16) | 6.78 ac (±1.10) | 6.70 ab (±0.84) | 7.99 bcd (±0.34) | 8.73 d (±0.31) | 5.24 a (±0.71) |
| TFAA | 14.10 ab (±0.95) | 16.68 c (±0.41) | 14.18 ac (±1.33) | 14.35 ac (±1.10) | 15.59 bc (±0.60) | 16.46 bc (±0.37) | 11.87 a (±1.11) |
| Amino Acid (μM/g) | LF-Intact | LF-Hydro | Intact | Hydro-A | Hydro-BCD | Hydro-DCB | Artemia |
|---|---|---|---|---|---|---|---|
| Aspartic Acid | 0.62 (±0.02) | 0.70 (±0.06) | 0.75 (±0.03) | 0.73 (±0.02) | 0.72 (±0.06) | 0.66 (±0.01) | 0.46 (±0.01) |
| Glutamic Acid | 1.34 (±0.05) | 1.29 (±0.09) | 1.45 (±0.09) | 1.25 (±0.03) | 1.45 (±0.08) | 1.19 (±0.06) | 0.93 (±0.05) |
| Asparagine | 0.45 (±0.03) | 0.42 (±0.04) | 0.57 (±0.04) | 0.50 (±0.01) | 0.51 (±0.06) | 0.43 (±0.01) | 0.59 (±0.09) |
| Serine | 0.30 (±0.04) | 0.29 (±0.01) | 0.51 (±0.06) | 0.52 (±0.05) | 0.66 (±0.09) | 0.42 (±0.02) | 0.72 (±0.13) |
| Glutamine | 0.55 (±0.03) | 0.52 (±0.04) | 0.83 (±0.04) | 0.68 (±0.02) | 0.86 (±0.09) | 0.52 (±0.02) | 0.70 (±0.07) |
| Histidine | 0.73 (±0.07) | 0.54 (±0.09) | 0.56 (±0.07) | 0.42 (±0.07) | 0.41 (±0.03) | 0.34 (±0.02) | 0.25 (±0.08) |
| Glycine | 1.89 (±0.45) | 2.64 (±0.49) | 1.60 (±0.26) | 1.43 (±0.25) | 1.53 (±0.23) | 1.47 (±0.14) | 0.70 (±0.13) |
| Threonine | 0.64 (±0.06) | 0.88 (±0.13) | 0.62 (±0.04) | 0.54 (±0.06) | 0.59 (±0.04) | 0.51 (±0.04) | 0.31 (±0.04) |
| Arginine | 0.07 (±0.01) | 0.05 (±0.00) | 0.07 (±0.01) | 0.06 (±0.01) | 0.05 (±0.00) | 0.04 (±0.00) | 0.03 (±0.00) |
| Alanine | 0.50 (±0.06) | 0.36 (±0.04) | 0.50 (±0.04) | 0.57 (±0.07) | 0.51 (±0.11) | 0.40 (±0.01) | 0.84 (±0.17) |
| Tyrosine | 0.13 (±0.02) | 0.19 (±0.02) | 0.16 (±0.01) | 0.14 (±0.01) | 0.18 (±0.00) | 0.15 (±0.01) | 0.04 (±0.00) |
| Lysine | 0.71 (±0.06) | 0.79 (±0.05) | 0.70 (±0.04) | 0.60 (±0.05) | 0.53 (±0.02) | 0.45 (±0.05) | 0.24 (±0.02) |
| Methionine | 0.34 (±0.04) | 0.34 (±0.04) | 0.39 (±0.03) | 0.34 (±0.05) | 0.33 (±0.07) | 0.55 (±0.04) | 0.40 (±0.01) |
| Valine | 1.91 (±0.09) | 1.85 (±0.06) | 1.83 (±0.08) | 1.95 (±0.08) | 1.63 (±0.04) | 2.17 (±0.12) | 1.80 (±0.03) |
| Cysteine | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Tryptophan | 0.05 (±0.00) | 0.06 (±0.00) | 0.08 (±0.00) | 0.08 (±0.00) | 0.10 (±0.01) | 0.08 (±0.00) | 0.06 (±0.01) |
| Phenylalanine | 0.18 (±0.01) | 0.21 (±0.02) | 0.23 (±0.03) | 0.22 (±0.00) | 0.23 (±0.01) | 0.22 (±0.01) | 0.12 (±0.00) |
| Isoleucine | 0.10 (±0.00) | 0.06 (±0.01) | 0.10 (±0.01) | 0.10 (±0.01) | 0.05 (±0.02) | 0.03 (±0.00) | 0.04 (±0.00) |
| Leucine | 0.21 (±0.01) | 0.17 (±0.02) | 0.24 (±0.01) | 0.20 (±0.01) | 0.14 (±0.01) | 0.12 (±0.01) | 0.09 (±0.00) |
| Proline | 0.54 (±0.06) | 0.74 (±0.13) | 0.66 (±0.03) | 1.05 (±0.25) | 0.98 (±0.16) | 1.08 (±0.10) | 1.33 (±0.07) |
| IDAA | 4.94 (±0.26) | 4.96 (±0.24) | 4.82 (±0.26) | 4.51 (±0.10) | 4.05 (±0.13) | 4.51 (±0.02) | 3.35 (±0.11) |
| DAA | 6.30 (±0.54) | 7.16 (±0.68) | 7.04 (±0.28) | 6.86 (±0.63) | 7.39 (±0.31) | 6.36 (±0.27) | 6.32 (±0.66) |
| TFAA | 11.24 (±0.80) | 12.12 (±0.92) | 11.86 (±0.53) | 11.37 (±0.63) | 11.44 (±0.41) | 10.85 (±0.25) | 9.66 (±0.77) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molinari, G.S.; Wojno, M.; Terova, G.; Wick, M.; Riley, H.; Caminiti, J.T.; Kwasek, K. A Novel Approach in the Development of Larval Largemouth Bass Micropterus salmoides Diets Using Largemouth Bass Muscle Hydrolysates as the Protein Source. Animals 2023, 13, 373. https://doi.org/10.3390/ani13030373
Molinari GS, Wojno M, Terova G, Wick M, Riley H, Caminiti JT, Kwasek K. A Novel Approach in the Development of Larval Largemouth Bass Micropterus salmoides Diets Using Largemouth Bass Muscle Hydrolysates as the Protein Source. Animals. 2023; 13(3):373. https://doi.org/10.3390/ani13030373
Chicago/Turabian StyleMolinari, Giovanni S., Michal Wojno, Genciana Terova, Macdonald Wick, Hayden Riley, Jeffrey T. Caminiti, and Karolina Kwasek. 2023. "A Novel Approach in the Development of Larval Largemouth Bass Micropterus salmoides Diets Using Largemouth Bass Muscle Hydrolysates as the Protein Source" Animals 13, no. 3: 373. https://doi.org/10.3390/ani13030373
APA StyleMolinari, G. S., Wojno, M., Terova, G., Wick, M., Riley, H., Caminiti, J. T., & Kwasek, K. (2023). A Novel Approach in the Development of Larval Largemouth Bass Micropterus salmoides Diets Using Largemouth Bass Muscle Hydrolysates as the Protein Source. Animals, 13(3), 373. https://doi.org/10.3390/ani13030373