Simple Summary
Recently, the study of amphibian behavioral ecology has received increased interest from ethologists and evolutionary biologists. In fact, plethodontid salamanders (family Plethodontidae) are often used as model organisms to better understand different aspects of behavioral adaptation. We reviewed the recent scientific literature published on the behavioral ecology of European cave salamanders belonging to the genus Speleomantes, to highlight recent advancements and possible future directions for successful research. We found that, in recent years, several aspects of Speleomantes behavior were investigated, such as trophic strategies and parental care, while others were neglected, in particular, chemical communication at the intraspecfic level. Finally, we propose European cave salamanders as useful models to understand the gradual adaptation of behaviors that facilitate the permanent colonization of subterranean habitats.
Abstract
There is a recent growing interest in the study of evolutionary and behavioral ecology of amphibians. Among salamanders, Plethodontidae is the most speciose family, with more than 500 species, while in Europe, there are only 8 species, all belonging to the genus Speleomantes. European plethodontids recently received increasing attention with regard to the study of their natural history, ecology and behavior; however, the lack of standardized data, especially for the latter, hampers comparative analysis with the species from the New World. We here synthetized the recent advances in Speleomantes behavioral ecology, considering as a starting point the comprehensive monography of Lanza and colleagues published in 2006. We identified the behavioral categories that were investigated the most, but we also highlighted knowledge gaps and provided directions for future studies. By reviewing the scientific literature published within the period 2006–2022, we observed a significant increase in the number of published articles on Speleomantes behavior, overall obtaining 36 articles. Behavioral studies on Speleomantes focused mainly on trophic behavior (42%), and on intraspecific behavior (33%), while studies on pheromonal communication and interspecific behavioral interactions were lacking. In addition, most of the studies were observational (83%), while the experimental method was rarely used. After providing a synthesis of the current knowledge, we suggest some relevant topics that need to be considered in future research on the behavioral ecology of European plethodontids, highlighting the importance of a more integrative approach in which both field observations and planned experiments are used.
Keywords:
behavior; comparative ethology; experiments; Plethodontidae; salamanders; Hydromantes; Speleomantes 1. Introduction
The behavioral ecology of amphibians remains largely understudied [1,2], although there is growing interest in the study of ecological processes and adaptive behaviors that shape the evolutionary dynamics of amphibian species and populations [3]. Amphibians represent a highly diverse group that include species with different habitus (from fossorial to exclusively aquatic), different reproductive strategies (oviparous and ovoviviparous) and even individuals that revolutionize their shape through life (larva vs. adult form). According to the intrinsic features of the species, but also to the local environmental conditions, the behavior of amphibians may change dramatically. For examples, in species adopting an r reproductive strategy, eggs are produced and laid by females in mass, which are often externally fertilized. On the other hand, in K-selected species, parents may spend substantial time taking care of their eggs and newborns, actively fighting against potential predators, driving them to a safer environment, or even feeding their brood [4]. The evolution of parental care in amphibians indeed seems to have played an important role for their colonization of different habitats. Looking at bi-phasic species, the behaviors of aquatic larvae can be completely different from those adopted by terrestrial adults. It is, therefore, quite complicated to standardize and compare amphibians’ behavior, unless specific groups with a similar ecology and life traits are considered.
Among Urodela, the family Plethodontidae (Gray, 1850) is the most speciose [4]. This family comprises more than 500 species out of 800 worldwide salamander and newt species, of which the overwhelming majority is found in Northern, Central and Southern America, while there are just 8 species in Europe and only 1 in Asia [4,5,6]. Plethodontids are characterized by the absence of lungs at the adult stage, and by the presence of naso-labial grooves that connect the upper lip to the external naris [4]. The main function of these peculiar structures is to canalize chemicals towards the olfactory structures [7,8].The wide diversity and the high local abundances that New World Plethodontids can reach make them relevant nodes within local ecological webs, where functional guilds are composed of many different interacting species [9,10,11,12]. Indeed, epigean plethodontids are predators which often occupy an intermediate position in the local food web, meaning that they hold the critical role of being prey and predators at the same time. Many species of New World plethodontids occur sympatrically, a condition that allowed several studies on species habitat selection and competition to be performed. Plethodontid salamanders have been even used as proxy for biodiversity assessments and monitoring in North American forests, cases that further highlight the importance of this group of amphibians and the need to deepen our knowledge on their biology and life traits [3].
On the other hand, the discovery of the Old World plethodontid species is relatively recent, and studies on their ecology and behavior lagged behind compared to their American relatives. Some not exclusive causes may contribute to this knowledge gap. As far as we know, Asian and European plethodontid species usually do not occur in sympatry, preventing the completion of studies on competitive behaviors of species living sympatrically. To this purpose, syntopy between two European plethodontid species has been artificially created, but such unnatural conditions do not provide any substantial information for advancing our knowledge on their behavior. Furthermore, although eight species of European plethodontids are currently known, they have been historically considered equivalent in their biology and ecology, meaning that single-species studies were often translated to the whole genus (see [13] and references therein). Indeed, only recently has more emphasis been given on evaluating interspecific (or even intraspecific) divergences among European plethodontid species, emphasis that strongly contributed in raising the number of studies performed. Furthermore, European plethodontids usually occur in subterranean environments, habitats where prolonged studies and monitoring activities are more challenging compared to surface ones.
American plethodontids became one of the main amphibian model systems used to better understand ecological questions in amphibians such as interspecific competition, predation and hybridization. Starting from field observations, classical ecological manipulative experiments were carried out, initially in natural woodland habitats [14,15,16]. However, in natural settings, there are many external uncontrollable factors that may influence the observed outcomes. For these reasons, plethodontids are often tested in laboratories to facilitate the avoidance of external confounding factors, while simultaneously controlling biological variables such as age, sex or reproductive status [17,18,19].
In this context, the current understanding of the behavioral ecology of European plethodontids, known as European cave salamanders, remains poorly developed in comparison to American plethodontids. The eight European species belong to the genus Speleomantes Dubois, 1984, although they are sometimes referred to as Hydromantes [5,20,21]. However, the use of Speleomantes better highlights the phylogenetic independence of European plethodontids, which form a well-defined monophyletic group with a large genetic distance from the five Californian species of Hydromantes [22,23,24]. All European cave salamanders are fully terrestrial and usually inhabit habitats where specific microclimatic conditions (e.g., high humidity and relatively low temperature variations) occur, such as humid crevices, forest floor environments and natural or artificial subterranean habitats [13,25,26]. Historically, behavioral studies on European plethodontids were sporadic because zoologists were focusing mainly on species description at the morphological and genetical level [25,27].
The monography dedicated to the taxonomy, biogeography and ecology of European cave salamanders written by Lanza et al. [13] constitutes the main reference concerning the natural history, ecology and behavior of the genus Speleomantes. This monograph lists under the chapter “Ethology” four sub-sections: “Feeding behaviour”, “Activity, habitat use and displacement”, “Antipredatory adaptations” and “Communication”. Concerning “Feeding behaviour”, most of the studies dealt with the anatomical structure of the eye [28] and of the tongue [29,30]. This sub-section summarizes the extraordinary tongue protrusion capability of Speleomantes and the ability to capture static prey in complete darkness [31,32]. Finally, the trophic strategy of Speleomantes is described as an “ambush strategy”, although this assertion seems based more on sporadic observations than on robust evidence [13]. The second sub-section, “Activity, habitat use and displacement”, relates exclusively to the species’ auto-ecological requirements, such as habitat features and seasonal activity, with little connection to behavior ecology sensu [3]. The third sub-section, “Antipredatory adaptations”, describes the presence of yellow-reddish or ochre dorsal colorations and the production of toxic skin secretion observed in many Speleomantes individuals and populations [13]. Finally, in the fourth sub-section, “Communication”, the presence of chemical intraspecific interactions was inferred from anatomical and histological studies on mating glands, but without providing any experimental evidence of their use during courtship and mating. In any case, the typical “nose tapping” behavior of plethodontids was reported in S. strinatii, suggesting that this specific behavior related to chemical communication is present [33]. Consequently, the relative paucity of the literature on European cave salamanders’ behavior does not allow robust comparative studies with American plethodontids [3,34,35].
The principal goal of this work is to provide a synthesis of the current knowledge on Speleomantes behavioral traits, and to stimulate new research aimed on this topic to produce a substantial amount of research that enables comparative studies with New World plethodontid species. Furthermore, we aim to propose this genus as a model to test eco-evolutionary hypotheses. For example, Speleomantes are facultative cave species that use subterranean environments to avoid unsuitable climatic conditions [36]. The microclimatic conditions offered by subterranean environments promote high efficiency for Speleomantes cutaneous respiration; therefore, European cave salamanders often establish stable populations in these habitats. The colonization of subterranean environments by epigean populations likely begins with a series of behavioral adjustments, allowing individuals to better exploit the new environment [37]. In this framework, Speleomantes could be considered as troglophiles sensu [38] and should be useful models to study the dynamics of meta-populations connecting subterranean and epigean ecosystems.
2. Materials and Methods
Peer-reviewed full papers, including proceedings from national meetings, published from 2006 to 2022, were considered in this study. Papers dealing exclusively, or giving relevant information, on European plethodontids were selected from online databases (i.e., Scopus and Clarivate Web of Sciences), searching for “Speleomantes” or “Hydromantes”. General articles just citing Speleomantes (or Hydromantes), faunistic lists, atlases and abstracts from scientific meetings were excluded. Papers dealing with the American Hydromantes species were discarded. Therefore, only European plethodontid were taken into consideration and analyzed.
Initially, we evaluated the existence of a recent trend in publications concerning the European genus Speleomantes, both in their absolute number and in relation to the continuous growth of global scientific production [39]. To obtain this latter information, we calculated the ratio between the number of papers dealing with Speleomantes or Hydromantes, but referring only to European species, and the number of papers with the word “amphibians” in the title, abstract and keywords retrieved each year from Scopus (accessed 10 August 2023). Temporal trends were analyzed using the Mann–Kendall non-parametric test [40], while we assessed an equal article distribution among Speleomantes species using a χ2 test for homogeneity [41], counting multi-species papers as one entry for each species.
In addition, we grouped European plethodontid studies according to their general subject, using as inspiration the chapter titles of the book Behavioral ecology of the Eastern Red-back Salamander—50 years of research by Jaeger et al. [3] as a benchmark. This choice was made for two reasons: (i) to use a clear operational definition of “behavioural ecology” that was already applied to salamanders by a leading research group for over 40 years, and (ii) to allow a better comparison with the research subjects on Plethodon cinereus, which is the most studied North American plethodontid species. However, we made some modifications or additions to better describe the studies published on European cave salamanders (see Table 1). For example, the first chapter “Interspecific competition between P. cinereus and P. shenandoah” was too specific and could not be applied to Speleomantes, while papers on courtship and parental behavior (topics not treated in [3]) were added to the category “Intraspecific social behaviour”. In order to better understand the types of studies within each behavioral category, we distinguished the two sub-categories “experimental” and “observational”. Papers were considered “experimental”, if they were carried out in controlled environments (i.e., in laboratory enclosures and terraria) or in the field with some type of intervention planned to discriminate among alternative hypotheses. In contrast, studies were considered “observational”, if they were based on data obtained from field populations without any previous manipulative intervention.

Table 1.
Behavioral ecology categories and related predominant species considered in this study (modified from [3]).
3. Results
We retrieved 26,180 peer-reviewed papers published from 2006 to 2022 with the word “amphibians” from the Scopus database. During the same time frame, 106 full papers dealing with European plethodontids were published (Supplementary Material, Table S1). During the study period, there was a positive trend in both the general scientific output on “amphibians” and for European plethodontids, expressed as the absolute number of papers published per year (Mann–Kendall trend test, Z = 3.831, p = 0.0001, and Z = 3.253, p = 0.001, for amphibian and Speleomantes papers, respectively). The relative proportion of Speleomantes papers showed a positive trend, even considering the general increase in overall papers published on amphibians (Figure 1; Mann–Kendall trend test, Z = 2.595, p = 0.009).
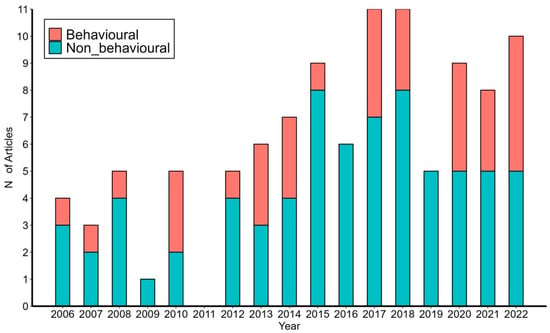
Figure 1.
Speleomantes papers published from 2006 to 2022. Red subgroups correspond to articles dealing with the behavioral ecology of Speleomantes, while blue ones cover the other topics.
Among these European plethodontid peer-reviewed papers, 34% (36/106) could be classified as broadly dealing with behavioral ecology as defined in this study (Figure 1), while the complete list of citations is given in Supplementary Material (Table S1). Behavioral papers mainly focused on two subject categories that accounted for 75% of the retrieved articles: “Foraging tactics” (42%, 15/36) and “Intraspecific social behaviour” (33%, 12/36; Figure 2). In the last category, six articles dealt with parental care in the genus Speleomantes, while the remaining articles focused on other intraspecific interactions. There were no studies on “Pheromonal glands and pheromonal communication” and on “Interspecific behavioural interactions”. Most studies were observational (83%, 30/36), while the remaining studies used an experimental approach in controlled or natural environments. During the study period, the lowest number of research articles was dedicated to S. ambrosii (n = 4) and the highest to S. strinatii (n = 15; Table 1), with a relatively homogeneous distribution among the eight Speleomantes species (χ2 = 13.271, df = 7, p = 0.066).
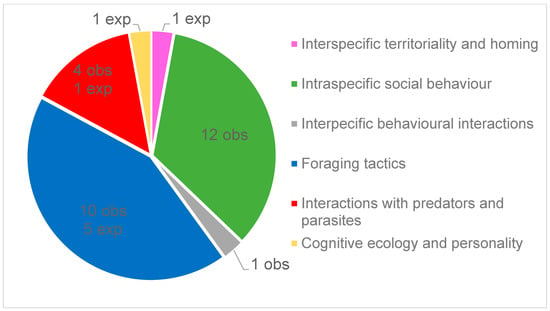
Figure 2.
Distribution of behavioral ecology subjects of papers published on the eight Speleomantes species, together with the detail of how many of these studies are observational (obs) or experimental (exp).
4. Discussion
Despite the increasing number of peer-reviewed papers published since the review of Lanza et al. [13], the behavioral ecology of European plethodontids belonging to the genus Speleomantes remains poorly known.
From our analysis, it is clear that recent research interests were focusing on three main topics in particular, related to foraging tactics, intraspecific social behavior (but the majority of papers were dealing with clutch guarding and parental care rather than on interactions among free-living conspecifics) and with predators or parasites. Foraging ecology is very relevant when assessing the role of salamanders as top predators on invertebrate communities that inhabit the superficial soil stratum [3]. Therefore, the dietary habits and feeding behavior of all eight species of Speleomantes were investigated at least once (Table S1). Researchers mostly explored the trophic spectrum of Speleomantes with the analysis of stomach contents, identifying diet composition in multiple conspecific populations and assessing how seasonality contributes to defining the type and amount of consumed prey [42,43]. In some instances, assessments of individuals’ food specialization were also performed [19,44]. On the other hand, many other aspects of Speleomantes foraging behavior are still unexplored or just hypothesized [45]. For example, we still do not know which foraging strategies these species adopt, or where they forage the most. Furthermore, do Speleomantes show prey preferences? Studies of gut contents were usually not supported by analyses aiming to quantify the local trophic supply, but see [46,47], making research topics related to individuals’ prey preferences virtually unexplored in many species. To date, all trophic ecology studies were performed using a stomach flushing technique on live individuals, a relatively simple technique that has been often performed in the field on different species of salamanders [46], while stable isotopes or DNA barcoding are rarely used [48].
Most of the studies regarding predation and parasitism on Speleomantes deal with the observations of these events in both epigean and ipogean environments. Consequently, the knowledge regarding this topic is still lacking (e.g., potential predators and predation avoidance) and more experimental studies should be carried out to explore the subject.
The second most explored research topic was related to reproductive behavior and parental care. Considering the cryptic behavior of Speleomantes, especially when they reproduce, the discovery of some nests in the wild is of high importance to comprehend their reproductive behavior, although it is limited to some species and mostly concentrated in the spring–summer period [36,49]. Indeed, most of the available information identifies the beginning of spring as the time in which females usually lay eggs, which are then protected until they hatch at the end of summer [50]. Different is the case of parental care. Using a facility provided by a semi-natural laboratory, some researchers were able to study female parental care in S. strinatii. Researchers observed an active protection of the mothers of both their eggs and newborns [51]. Future studies aiming to deepen research on this topic should include more species and (hopefully) include wild observations, as individuals may change their behavior when translocated into different environments [52].
The lack of interspecific behavioral studies may be due to the allopatric distribution characterizing the genus Speleomantes. This condition, to the best of our knowledge, does not occur in only two narrow contact zones of mainland Italy, where, inter alia, hybridization between species is present [53,54]. In the past, a couple of experiments involving the creation of artificial sympatry of two mainland Speleomantes species were performed; their aim was to study their habitat selection and competition [55]. However, data from such unnatural conditions are useless for this review; therefore, those studies were not considered. In addition, only two other terrestrial salamanders are found in sympatry with one Speleomantes species at a time, i.e., the fire salamander Salamandra salamandra and the Northern spectacled salamander Salamandrina perspicillata [13]. Such co-occurrence is mostly realized in epigean environments (e.g., in forested areas), although both species can occasionally exploit subterranean environments [56,57]. A few studies performed on this system provided the first information related to the potential mechanisms (e.g., temporal and trophic niche partitioning) that these species adopt to avoid competition [58,59]. However, more natural observations and experimental tests are needed to shed light on this topic.
The behavioral trait for which our knowledge is the poorest is Speleomantes communication and social interactions at an intraspecific level. We still have uncertainty on the nature of intraspecific interactions between individuals. Some authors highlighted the potential occurrence of agonistic interactions between age classes [60] within subterranean environments, while some others found no evidence of such behavior [61]. The single study performed on surface population does not report any evidence of agonistic interactions [62].
Finally, our review highlighted the relative low number of experimental studies published on European plethodontids. New World plethodontids are currently used as laboratory models for experimental studies on comparative and adaptive behavior [3]. Characteristics promoting the use of plethodontids as model species are their easy maintenance, survival and breeding in animal research facilities [3]. In controlled environments, selected individuals can be exposed to experimental conditions to test for multiple hypotheses. In fact, the experimental hypothesis-testing approach seems very informative in the study of behavioral ecology, since it allows researchers to infer causality [3,17,18]. Although, caveats should be always considered when extrapolating results observed in simplified, and possibly stressing, settings compared to natural habitats [19]. This type of approach was performed mostly on the continental species S. strinatii, with only one exception concerning S. italicus (see Table S1). However, similarly to many New World plethodontids, Speleomantes are medium-sized terrestrial salamanders that can be easily hosted in the laboratory within terraria or small containers for relatively long periods of time. However, reproducing a suitable microhabitat for these species may be not trivial. In fact, due to their strictly cutaneous respiration, European plethodontids are highly dependent on microclimatic conditions, which should be characterized by high air relative humidity, and air temperatures usually lower than 18 °C [26]. Therefore, maintaining these environmental features within such ranges would assure animal welfare. These environmental conditions can be reproduced only in laboratory cold rooms or in natural or artificial subterranean habitats, thus limiting the experimental approach, as we, in fact, observed. Based on our analysis of the recent published research on European cave salamanders, we propose a preliminary list of possible topics for future research (Table 2).
Table 2.
Suggested topics for future research on European cave salamanders species.
5. Future Directions
Our analysis also highlighted the scarcity of comparative studies, where the behavioral adaptations of the Speleomantes species are framed in an evolutionary perspective. These studies are useful to determine if variation in behavioral traits is correlated with the phylogenetic relatedness or with local ecological conditions experienced by the different species [63]. This comparative approach should be one of the most fruitful lines of research, because the phylogenetic relationships among the eight European cave salamander species have been relatively well investigated and quantified using different approaches [64,65].
Another relevant issue concerns understanding the evolutionary processes that allowed European cave salamanders to colonize subterranean environments [66]. This, in fact, is one of the hottest topics in evolutionary biology and for biospeleologists; although, it is not always easy to accomplish [52]. Most of the effort focused on understanding the adaptive traits characterizing the most adapted subterranean species (i.e., troglobites), while little attention was given to facultative cave species such as troglophiles and trogloxenes [38,67]. Specific traits characterizing troglobites (e.g., anophtalmia, lack of pigmentation, high longevity) are peculiar and very attractive for investigative purposes; indeed, researchers very often compare subterranean species with such characters with their genetically close-related epigean species [68,69,70]. However, the appearance (or disappearance) of a specific character can be a long stepwise process, where species not completely adapted to subterranean environments may provide key information on the intermediate processes from one end to the other. Improving behavioral studies on Speleomantes can be very useful to understanding such evolutionary processes, as behavioral adjustments likely take place at the very beginning of the colonization of subterranean environments [37,71]. Using Speleomantes as model species provides the (very rare) possibility to employ in experiments both epigean and subterranean conspecific populations, a condition that will help in producing more robust inferences and more clear results as it reduces potential divergences due to different life histories. A similar approach has already been successfully used by comparing the surface-dwelling and the cave-adapted populations of the fish Astyanax mexicanum [72,73] and those of the Pyrenean brook salamander Calotriton asper. The adults of this latter salamander are terrestrial but mate and lay eggs in streams and produce aquatic larvae. In this species, both epigean and hypogean populations are known [74], and many studies aiming to assess behavioral divergences between surface and subterranean populations have been performed. For example, to test the hypothesis that cave-adapted individuals evolved increased food-finding abilities, the prey detection performances of surface and subterranean populations were compared [72]. This study showed that epigean Pyrenean brook salamanders possess a predisposition to successfully forage at night and that these salamanders improve their ability to forage in total darkness when experimentally maintained in cave conditions for long periods of time. Therefore, this plasticity in foraging behavior may facilitate the permanent colonization of subterranean habitats, in which food is limited, and light is reduced or completely absent [72].
In conclusion, our analysis showed the many knowledge gaps hampering a proper understanding of the behavioral ecology of Speleomantes species. Starting from the many observational studies published in recent years, an experimental approach should be promoted to better understand the behavioral adaptations described for the different species. For example, under controlled conditions, such as when using terraria or mesocosms, the existence and relative strength of intra- and interspecific chemical communication could be successfully tested. Finally, a more evolutionary and comparative approach should also be encouraged, as is the case of Lunghi et al., (2018) [50], which is the only study comparing the reproductive behaviors of all eight species of European cave salamanders.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ani13233667/s1, Table S1: Complete list of Speleomantes paper 2006–2022 papers highlieted in yellow were analised.
Author Contributions
Conceptualization A.C., G.R. and S.S.; methodology, S.S; formal analysis, S.S.; data curation, G.R., E.L. and S.S.; writing—original draft preparation, S.S.; writing—review and editing, A.C. and E.L.; visualization, G.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in the Supplementary material.
Acknowledgments
We are grateful to Mauro Valerio Pastorino, from the Gruppo Speleologico Ligure “Arturo Issel”, for providing access to the Biospeleological Station “A. Issel” and for the many useful discussions over the years on the biology of cave salamanders.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Owens, I.P. Where Is Behavioural Ecology Going? Trends Ecol. Evol. 2006, 21, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, S.R.; Silla, A.J.; Byrne, P.G. Animal Personality and Behavioral Syndromes in Amphibians: A Review of the Evidence, Experimental Approaches, and Implications for Conservation. Behav. Ecol. Sociobiol. 2018, 72, 79. [Google Scholar] [CrossRef]
- Jaeger, R.G.; Gollmann, B.; Anthony, C.D.; Gabor, C.R.; Kohn, N.R. Behavioral Ecology of the Eastern Red-Backed Salamander: 50 Years of Research; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Wake, D.B. Lungless Salamanders (Plethodontidae). In Grzimek’s Animal Life Encyclopaedia; Hutchins, M., Duellman, W.E., Schlager, N., Eds.; Gale/Cengage Learning: Farmington Hills, MI, USA, 2003; Volume 6, pp. 389–404. [Google Scholar]
- Wake, D.B. Taxonomy of Salamanders of the Family Plethodontidae (Amphibia: Caudata). Zootaxa 2012, 3484, 75–82. [Google Scholar] [CrossRef]
- Min, M.S.; Yang, S.-Y.; Bonett, R.; Vieites, D.; Brandon, R.; Wake, D. Discovery of the First Asian Plethodontid Salamander. Nature 2005, 435, 87–90. [Google Scholar] [CrossRef] [PubMed]
- Dawley, E.M. Recognition of Individual, Sex and Species Odours by Salamanders of the Plethodon glutinosus-P. jordani Complex. Anim. Behav. 1984, 32, 353–361. [Google Scholar] [CrossRef]
- Dawley, E.M. Comparative Morphology of Plethodontid Olfactory and Vomeronasal Organs: How Snouts Are Packed. Herpetol. Monogr. 2017, 31, 169–209. [Google Scholar] [CrossRef]
- Burton, T.M.; Likens, G.E. Salamander Populations and Biomass in the Hubbard Brook Experimental Forest, New Hampshire. Copeia 1975, 541–546. [Google Scholar] [CrossRef]
- Welsh, H.H., Jr.; Droege, S. A Case for Using Plethodontid Salamanders for Monitoring Biodiversity and Ecosystem Integrity of North American Forests. Conserv. Biol. 2001, 15, 558–569. [Google Scholar] [CrossRef]
- Petranka, J.W. Salamanders of the United States and Canada; Smithsonian Institution Press: Washington, DC, USA, 1998. [Google Scholar]
- Cody, M.L.; Smallwood, J.A. Long-Term Studies of Vertebrate Communities; Academic Press: New York, NY, USA, 1996; ISBN 0-08-053562-3. [Google Scholar]
- Lanza, B. A Review of Systematics, Taxonomy, Genetics, Biogeography and Natural History of the Genus Speleomantes Dubois, 1984 (Amphibia Caudata Plethodontidae). Trieste Mus. Civ. Stor. Nat. 2006, 52, 5–135. [Google Scholar]
- Hairston, N.G. Evolution under Interspecific Competition: Field Experiments on Terrestrial Salamanders. Evolution 1980, 34, 409–420. [Google Scholar] [CrossRef]
- Hairston, N.G. Community Ecology and Salamander Guilds; Cambridge University Press: Cambridge, UK, 1987; ISBN 0-521-32578-1. [Google Scholar]
- Hairston, N.G. The Experimental Test of an Analysis of Field Distributions: Competition in Terrestrial Salamanders. Ecology 1980, 61, 817–826. [Google Scholar] [CrossRef]
- Campbell, D.L.; Weiner, S.A.; Starks, P.T.; Hauber, M.E. Context and Control: Behavioural Ecology Experiments in the Laboratory. Ann. Zool. Fennici 2009, 46, 112–123. [Google Scholar] [CrossRef]
- Ylönen, H.; Wolff, J.O. Experiments in Behavioural Ecology and the Real World. Trends Ecol. Evol. 1999, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Bailey, J. Does the Stress of Laboratory Life and Experimentation on Animals Adversely Affect Research Data? A Critical Review. Altern. Lab. Anim. 2018, 46, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; Manenti, R.; Cianferoni, F.; Ceccolini, F.; Veith, M.; Corti, C.; Ficetola, G.F.; Mancinelli, G. Interspecific and Interpopulation Variation in Individual Diet Specialization: Do Environmental Factors Have a Role? Ecology 2020, 101, e03088. [Google Scholar] [CrossRef]
- Bruni, G. Tail-Straddling Walk and Spermatophore Transfer in Hydromantes italicus: First Observations for the Genus and Insights about Courtship Behavior in Plethodontid Salamanders. Herpetol. Rev. 2020, 51, 673–680. [Google Scholar]
- Speybroeck, J.; Beukema, W.; Crochet, P.-A. A Tentative Species List of the European Herpetofauna (Amphibia and Reptilia)—An Update. Zootaxa 2010, 2492, 1–27. [Google Scholar] [CrossRef]
- Speybroeck, J.; Beukema, W.; Dufresnes, C.; Fritz, U.; Jablonski, D.; Lymberakis, P.; Martínez-Solano, I.; Razzetti, E.; Vamberger, M.; Vences, M. Species List of the European Herpetofauna–2020 Update by the Taxonomic Committee of the Societas Europaea Herpetologica. Amphib.-Reptil. 2020, 41, 139–189. [Google Scholar] [CrossRef]
- Bingham, R.E.; Papenfuss, T.J.; Lindstrand, L.; Wake, D.B. Phylogeography and Species Boundaries in the Hydromantes shastae Complex, with Description of Two New Species (Amphibia; Caudata; Plethodontidae). Bull. Mus. Comp. Zool. 2018, 161, 403–427. [Google Scholar] [CrossRef]
- Lanza, B.; Andreone, F.; Bologna, M.; Corti, C.; Razzetti, E. Amphibia—Fauna d’Italia; Edizioni Calderini: Bologna, Italy, 2007; Volume 42. [Google Scholar]
- Ficetola, G.F.; Lunghi, E.; Canedoli, C.; Padoa-Schioppa, E.; Pennati, R.; Manenti, R. Differences between Microhabitat and Broad-Scale Patterns of Niche Evolution in Terrestrial Salamanders. Sci. Rep. 2018, 8, 10575. [Google Scholar] [CrossRef]
- Lanza, B.; Caputo Barucchi, V.; Nascetti, G.; Bullini, L. Morphologic and Genetic Studies on the European Plethodontid Salamanders: Taxonomic Inferences (Genus Hydromantes); Museo Regionale Scienze Naturali: Torino, Italy, 1995; Volume 16, ISBN 88-86041-10-1. [Google Scholar]
- Serra, G.; Crnjar, R.; Lilliu, V.; Cardinale Ciccotti, F.; Spiga, M.A. Morphometric Analysis of the Retinic Photoreceptors in the Cave Salamander Hydromantes Genei (Speleomantes) (Temm. and Schl.): Functional and Anatomical Considerations. Ital. J. Anat. Embryol. 1995, 100, 99–110. [Google Scholar]
- Lombard, R.E.; Wake, D.B. Tongue Evolution in the Lungless Salamanders, Family Plethodontidae I. Introduction, Theory and a General Model of Dynamics. J. Morphol. 1976, 148, 265–286. [Google Scholar] [CrossRef] [PubMed]
- Eric Lombard, R.; Wake, D.B. Tongue Evolution in the Lungless Salamanders, Family Plethodontidae. II. Function and Evolutionary Diversity. J. Morphol. 1977, 153, 39–79. [Google Scholar] [CrossRef] [PubMed]
- Roth, G. Experimental Analysis of the Prey Catching Behavior of Hydromantes italicus Dunn (Amphibia, Plethodontidae). J. Comp. Physiol. 1976, 109, 47–58. [Google Scholar] [CrossRef]
- Roth, G. The Role of Stimulus Movement Patterns in the Prey Catching Behavior of Hydromantes genei (Amphibia, Plethodontidae). J. Comp. Physiol. 1978, 123, 261–264. [Google Scholar] [CrossRef]
- Zanetti, L.; Salvidio, S. Preliminary Data on the Territorial Behaviour of Speleomantes strinatii; Museo Civico di Zoologia: Roma, Italy, 2006; pp. 160–161. [Google Scholar]
- Jaeger, R.G.; Forester, D.C. Social Behavior of Plethodontid Salamanders. Herpetologica 1993, 49, 163–175. [Google Scholar]
- Bruce, R. Sexual Size Dimorphism in the Plethodontidae. In The Biology of Plethodontid Salamanders; Bruce, R.C., Jaeger, R.G., Houck, L.D., Eds.; Klucer Academic: New York, NY, USA, 2000; pp. 243–260. [Google Scholar]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Do Cave Features Affect Underground Habitat Exploitation by Non-Troglobite Species? Acta Oecologica 2014, 55, 29–35. [Google Scholar] [CrossRef]
- Lunghi, E.; Martinez, A.; Hesselberg, T.; Mammola, S. Behavioural Adjustments Enable the Colonization of Subterranean Environments. Zool. J. Linn. Soc. 2023. [Google Scholar] [CrossRef]
- Mammola, S. Finding Answers in the Dark: Caves as Models in Ecology Fifty Years after Poulson and White. Ecography 2019, 42, 1331–1351. [Google Scholar] [CrossRef]
- Fire, M.; Guestrin, C. Over-Optimization of Academic Publishing Metrics: Observing Goodhart’s Law in Action. GigaScience 2019, 8, giz053. [Google Scholar] [CrossRef]
- Gilbert, R. Skewed Distributions and Goodness-of-Fit Tests. In Statistical Methods for Environmental Pollution Monitoring; Wiley: Cambridge, MA, USA, 1987; pp. 152–163. [Google Scholar]
- Siegel, S. Nonparametric Statisticsfor the Behavioral Sciences; McGraw-Hill Companies: New York, NY, USA, 1956. [Google Scholar]
- Lunghi, E.; Cianferoni, F.; Ceccolini, F.; Veith, M.; Manenti, R.; Mancinelli, G.; Corti, C.; Ficetola, G.F. What Shapes the Trophic Niche of European Plethodontid Salamanders? PLoS ONE 2018, 13, e0205672. [Google Scholar] [CrossRef]
- Lunghi, E.; Cianferoni, F.; Corti, C.; Zhao, Y.; Manenti, R.; Ficetola, G.F.; Mancinelli, G. The Trophic Niche of Subterranean Populations of Speleomantes italicus. Sci. Rep. 2022, 12, 18257. [Google Scholar] [CrossRef] [PubMed]
- Salvidio, S.; Oneto, F.; Ottonello, D.; Costa, A.; Romano, A. Trophic Specialization at the Individual Level in a Terrestrial Generalist Salamander. Can. J. Zool. 2015, 93, 79–83. [Google Scholar] [CrossRef]
- Cianferoni, F.; Lunghi, E. Inferring on Speleomantes Foraging Behavior from Gut Contents Examination. Animals 2023, 13, 2782. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Romano, A.; Rosa, G.; Salvidio, S. Weighted Individual-Resource Networks in Prey–Predator Systems: The Role of Prey Availability on the Emergence of Modular Structures. Integr. Zool. 2020, 17, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Salvidio, S.; Romano, A.; Oneto, F.; Ottonello, D.; Michelon, R. Different Season, Different Strategies: Feeding Ecology of Two Syntopic Forest-Dwelling Salamanders. Acta Oecologica 2012, 43, 42–50. [Google Scholar]
- Gillespie, J.H. Application of Stable Isotope Analysis to Study Temporal Changes in Foraging Ecology in a Highly Endangered Amphibian. PLoS ONE 2013, 8, e53041. [Google Scholar] [CrossRef] [PubMed]
- Lunghi, E.; De Falco, G.; Buschettu, S.; Murgia, R.; Mulas, C.; Mulargia, M.; Canedoli, C.; Ficetola, G.F.; Manenti, R. First Data on Nesting Ecology and Behaviour in the Imperial Cave Salamander Hydromantes imperialis. North-West. J. Zool. 2015, 11, 324–330. [Google Scholar]
- Lunghi, E.; Corti, C.; Manenti, R.; Barzaghi, B.; Buschettu, S.; Canedoli, C.; Cogoni, R.; De Falco, G.; Fais, F.; Manca, A.; et al. Comparative Reproductive Biology of European Cave Salamanders (Genus Hydromantes): Nesting Selection and Multiple Annual Breeding. Salamandra 2018, 54, 101–108. [Google Scholar]
- Oneto, F.; Ottonello, D.; Pastorino, M.V.; Salvidio, S. Posthatching Parental Care in Salamanders Revealed by Infrared Video Surveillance. J. Herpetol. 2010, 44, 649–653. [Google Scholar] [CrossRef]
- Mammola, S.; Lunghi, E.; Bilandžija, H.; Cardoso, P.; Grimm, V.; Schmidt, S.I.; Hesselberg, T.; Martínez, A. Collecting Eco-evolutionary Data in the Dark: Impediments to Subterranean Research and How to Overcome Them. Ecol. Evol. 2021, 11, 5911–5926. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Lunghi, E.; Cimmaruta, R.; Manenti, R. Transgressive Niche across a Salamander Hybrid Zone Revealed by Microhabitat Analyses. J. Biogeogr. 2019, 46, 1342–1354. [Google Scholar] [CrossRef]
- Bruni, G.; Chiocchio, A.; Nascetti, G.; Cimmaruta, R. Different Patterns of Introgression in a Three Species Hybrid Zone among European Cave Salamanders. Ecol. Evol. 2023, 13, e10437. [Google Scholar] [CrossRef]
- Cimmaruta, R.; Forti, G.; Lucente, D.; Nascetti, G. Thirty Years of Artificial Syntopy between Hydromantes italicus and H. ambrosii ambrosii (Amphibia, Plethodontidae). Amphib.-Reptil. 2013, 34, 413–420. [Google Scholar] [CrossRef]
- Manenti, R.; Ficetola, G. Salamanders Breeding in Subterranean Habitats: Local Adaptations or Behavioural Plasticity? J. Zool. 2013, 289, 182–188. [Google Scholar] [CrossRef]
- Angelini, C.; Vanni, S.; Vignoli, L. Salamandrina Terdigitata (Bonnaterre, 1789) Salamandrina Perspicillata (Savi, 1821). Fauna D’italia 2007, 42, 228–237. [Google Scholar]
- Rosa, G.; Salvidio, S.; Costa, A. Disentangling Exploitative and Interference Competition on Forest Dwelling Salamanders. Animals 2023, 13, 2003. [Google Scholar] [CrossRef]
- Lunghi, E.; Corti, C.; Biaggini, M.; Zhao, Y.; Cianferoni, F. The Trophic Niche of Two Sympatric Species of Salamanders (Plethodontidae and Salamandridae) from Italy. Animals 2022, 12, 2221. [Google Scholar] [CrossRef]
- Salvidio, S. Spatial Segregation in the European Plethodontid Speleomantes strinatii in Relation to Age and Sex. Amphib.-Reptil. 2002, 23, 505–510. [Google Scholar]
- Ficetola, G.F.; Pennati, R.; Manenti, R. Spatial Segregation among Age Classes in Cave Salamanders: Habitat Selection or Social Interactions? Popul. Ecol. 2013, 55, 217–226. [Google Scholar] [CrossRef]
- Rosa, G.; Salvidio, S.; Costa, A. European Plethodontid Salamanders on the Forest Floor: Testing for Age-Class Segregation and Habitat Selection. J. Herpetol. 2022, 56, 27–33. [Google Scholar] [CrossRef]
- Bloomberg, S.P.; Garland, T.; Ives, A.R. Testing for Phylogenetic Signal in Comparative Data: Behavioral Traits Are More Labile. Evolution 2003, 57, 717–745. [Google Scholar]
- van der Meijden, A.; Chiari, Y.; Mucedda, M.; Carranza, S.; Corti, C.; Veith, M. Phylogenetic Relationships of Sardinian Cave Salamanders, Genus Hydromantes, Based on Mitochondrial and Nuclear DNA Sequence Data. Mol. Phylogenet. Evol. 2009, 51, 399–404. [Google Scholar] [CrossRef]
- Chiari, Y.; van der Meijden, A.; Mucedda, M.; Lourenco, J.M.; Hochkirch, A.; Veith, M. Phylogeography of Sardinian Cave Salamanders (Genus Hydromantes) Is Mainly Determined by Geomorphology. PLoS ONE 2012, 7, e32332. [Google Scholar] [CrossRef]
- Mammola, S.; Amorim, I.R.; Bichuette, M.E.; Borges, P.A.; Cheeptham, N.; Cooper, S.J.; Cardoso, P. Fundamental Research Questions in Subterranean Biology. Biol. Rev. 2020, 95, 1855–1872. [Google Scholar] [CrossRef]
- Howarth, F.G.; Moldovan, O.T. The Ecological Classification of Cave Animals and Their Adaptations. In Cave Ecology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2018; pp. 41–67. [Google Scholar]
- Hervant, F.; Mathieu, J.; Durand, J. Behavioural, Physiological and Metabolic Responses to Long-Term Starvation and Refeeding in a Blind Cave-Dwelling (Proteus anguinus) and a Surface-Dwelling (Euproctus asper) Salamander. J. Exp. Biol. 2001, 204, 269–281. [Google Scholar] [CrossRef]
- Beasley-Hall, P.G.; Bertozzi, T.; Bradford, T.M.; Foster, C.S.P.; Jones, K.; Tierney, S.M.; Humphreys, W.F.; Austin, A.D.; Cooper, S.J.B. Differential Transcriptomic Responses to Heat Stress in Surface and Subterranean Diving Beetles. Sci. Rep. 2022, 12, 16194. [Google Scholar] [CrossRef]
- Lunghi, E.; Bilandžija, H. Longevity in Cave Animals. Front. Ecol. Evol. 2022, 10, 874123. [Google Scholar] [CrossRef]
- Lipovšek, S.; Leitinger, G.; Janžekovič, F.; Kozel, P.; Dariš, B.; Perc, M.; Devetak, D.; Weiland, N.; Novak, T. Towards Understanding Partial Adaptation to the Subterranean Habitat in the European Cave Spider, Meta menardi: An Ecocytological Approach. Sci. Rep. 2019, 9, 9121. [Google Scholar] [CrossRef]
- Borowsky, R. Selection maintains the phenotypic divergence of cave and surface fish. Am. Nat. 2023, 202, 55–63. [Google Scholar] [CrossRef]
- Gross, J.B.; Boggs, T.E.; Rétaux, S.; Torres-Paz, J. Developmental and genetic basis of troglomorphic traits in the teleost fish Astyanax mexicanus. In Groundwater Ecology and Evolution; Malard, F., Griebler, C., Rétaux, S., Eds.; Academic Press: New York, NY, USA, 2023; pp. 351–371. [Google Scholar]
- Guillaume, O. Surface Newt Calotriton Asper Acclimation to Cave Conditions Improved Their Foraging Ability in Darkness. Front. Ecol. Evol. 2022, 10, 1057023. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).