
Association of SLIT3 and ZNF280B Gene Polymorphisms with Wool Fiber Diameter
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Traits
2.2. Genomic DNA Extraction and Wool Fiber Diameter Detection
2.3. Genetic Typing
2.4. SNP Validation
2.5. Real-Time Quantitative PCR
2.6. Statistical Analysis
3. Results
3.1. Genotyping Results for SLIT3 and ZNF280B and Population Index Analysis of the Loci in Alpine Merino Sheep
3.2. Association Analysis between ZNF280B and SLIT3 Genotypes and Wool Fiber Diameter of Alpine Merino Sheep
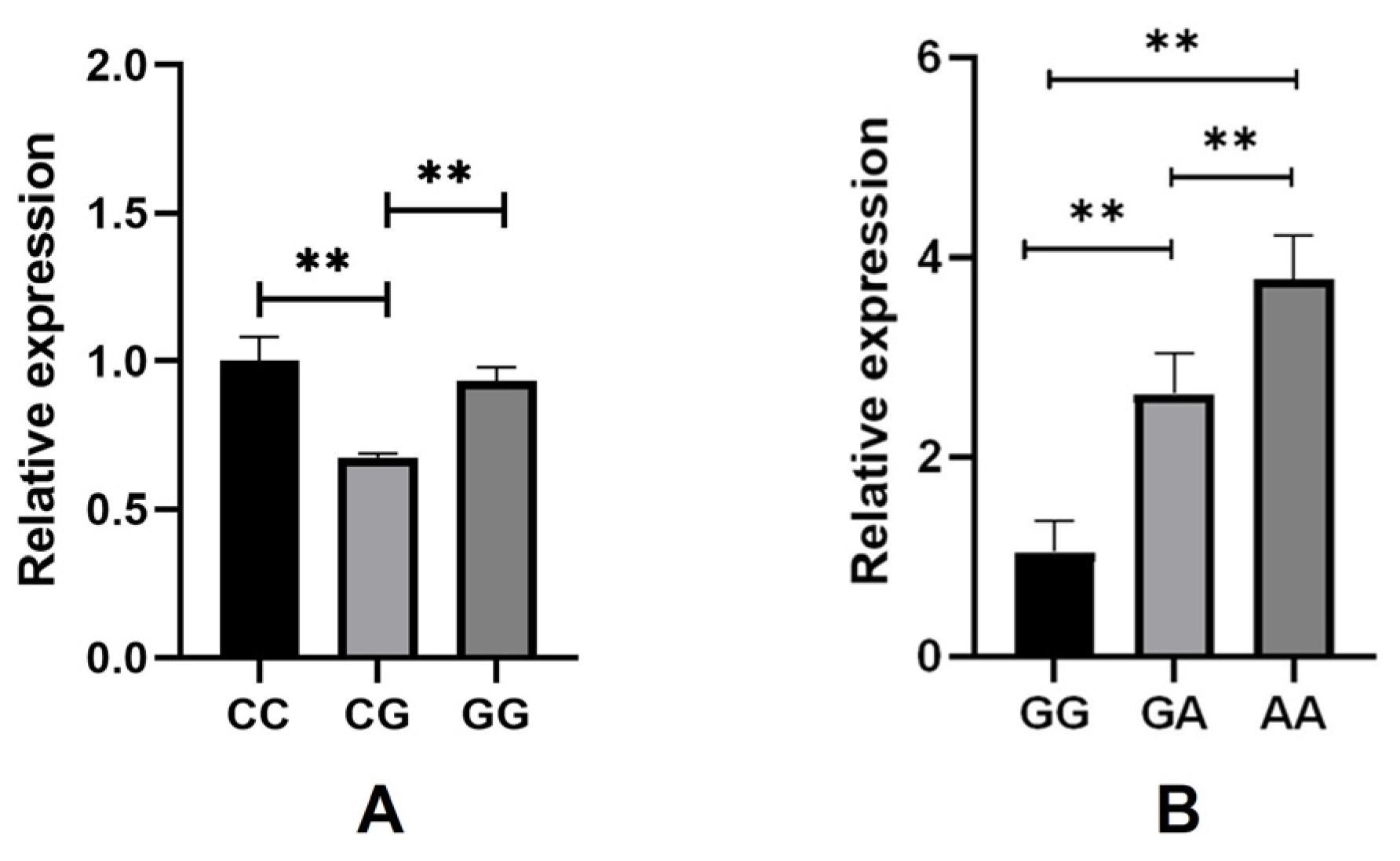
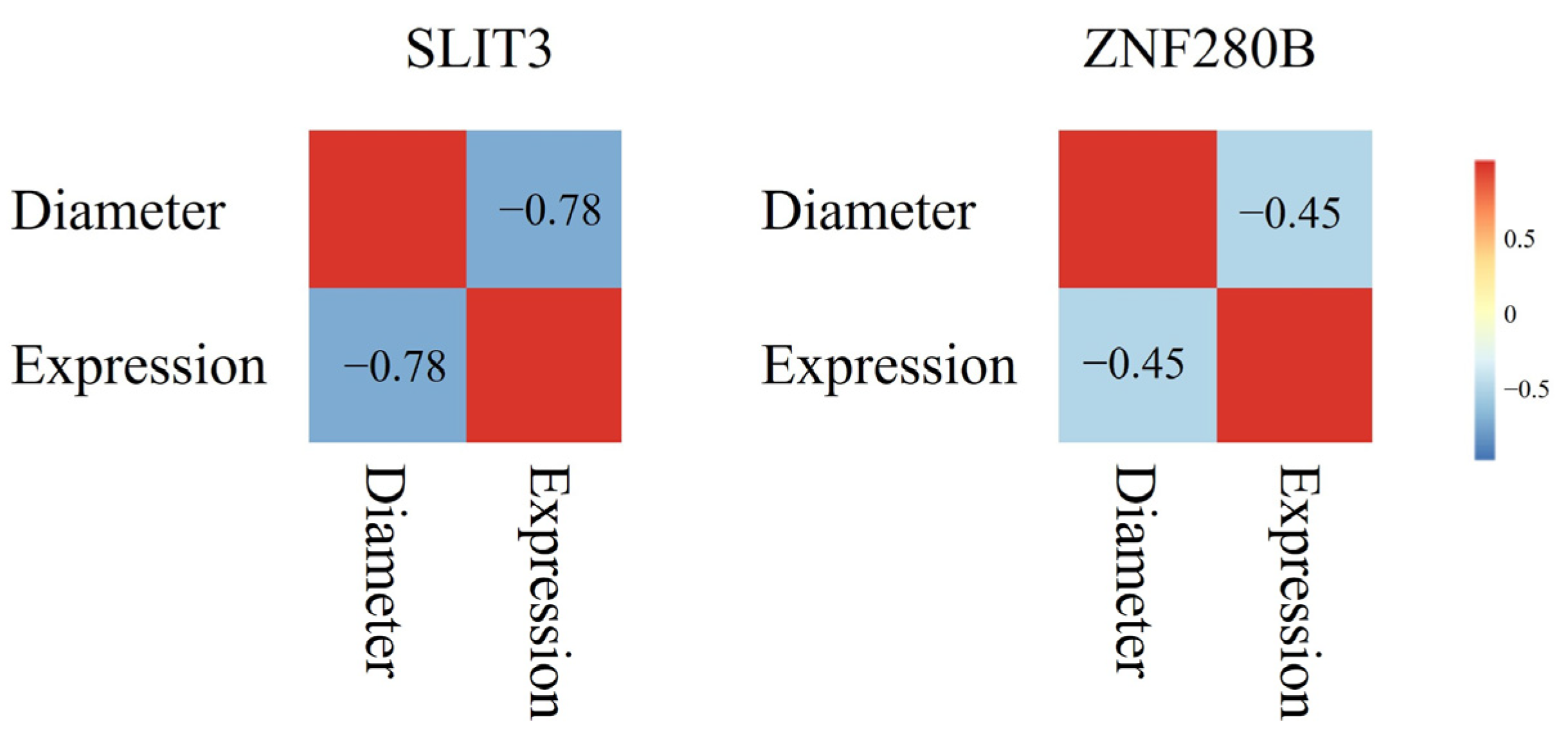
3.3. Expression Analysis of SLIT3 and ZNF280B in Skin Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.; Xiao, H. Research on the Development Strategy of China’s Wool Industry. J. Wuhan Textile Univ. 2016, 29, 14–18. [Google Scholar]
- Dry, F.W. The dominant N gene in New Zealand Romney sheep. Aust. J. Agric. Res. 1955, 6, 725–769. [Google Scholar] [CrossRef]
- Deng, X.; Liu, K. Wool quality inspection and sheep breeding and wool spinning product quality. Grass-Feed. Livest. 2000, 1, 15–17. [Google Scholar]
- Zhao, J.; Liu, N.; Liu, K.; He, J.; Yu, J.; Bu, R.; Cheng, M.; De, W.; Liu, J.; Li, H. Identification of genes and proteins associated with anagen wool growth. Anim. Genet. 2017, 48, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Purvis, I.W.; Franklin, I.R. Major genes and QTL influencing wool production and quality: A review. Genet. Sel. Evol. 2005, 37, 97–107. [Google Scholar] [CrossRef]
- Zhao, J.; Qin, H.; Xin, J.; Liu, N.; Han, R.; Perez-Campo, F.M.; Li, H. Discovery of genes and proteins possibly regulating mean wool fibre diameter using cDNA microarray and proteomic approaches. Sci. Rep. 2020, 10, 7726. [Google Scholar] [CrossRef] [PubMed]
- Aumailley, M. The laminin family. Cell Adh. Migr. 2013, 7, 48–55. [Google Scholar] [CrossRef]
- Di, J.; Xu, X.; Zhang, Y.; Tian, K.; Tian, Y.; Wu, W.; Yu, L. Study on differentially expressed genes in skin tissue of fine wool sheep with different wool fiber diameters. Acta Vet. Zootech. Sin. 2013, 44, 681–689. [Google Scholar]
- Zhao, H.; Hu, R.; Li, F.; Yue, X. Five SNPs Within the FGF5 Gene Significantly Affect Both Wool Traits and Growth Performance in Fine-Wool Sheep (Ovis aries). Front. Genet. 2021, 12, 732097. [Google Scholar] [CrossRef]
- Li, S.; Zhou, H.; Gong, H. Variation in the Ovine KAP6-3 Gene (KRTAP6-3) Is Associated with Variation in Mean Fibre Diameter-Associated Wool Traits. Genes 2017, 8, 204. [Google Scholar] [CrossRef]
- Song, Y.; Luo, Y.; Zhou, H.; Liu, X.; Li, S.; Hao, Z.; Shen, J.; Zhen, H.; Li, L.; Wang, J.; et al. Variation in caprine KRTAP1-3 and its association with cashmere fibre diameter. Gene 2022, 823, 146341. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhou, H.; Luo, Y.; Zhao, M.; Gong, H.; Hao, Z.; Hu, J.; Hickford, J.G.H. Variation in the Caprine KAP24-1 Gene Affects Cashmere Fibre Diameter. Animals 2019, 9, 15. [Google Scholar] [CrossRef]
- Zhao, M.; Zhou, H.; Luo, Y.; Wang, J.; Hu, J.; Liu, X.; Li, S.; Hao, Z.; Jin, X.; Song, Y.; et al. Variation in the Caprine Keratin-Associated Protein 27-1 Gene is Associated with Cashmere Fiber Diameter. Genes 2020, 11, 934. [Google Scholar] [CrossRef]
- Min, S.; Wang, W.J.; Yang, Y.L.; Gan, S.Q.; Zhang, Y.S.; Wang, J.H.; Liu, S.R.; Li, N. A novel polymorphism of IGFBP-3 gene and its relationship with several wool traits in Chinese Merino sheep. Hereditas 2008, 30, 1182–1186. [Google Scholar]
- Zhao, H.; Guo, T.; Lu, Z.; Liu, J.; Zhu, S.; Qiao, G.; Han, M.; Yuan, C.; Wang, T.; Li, F.; et al. Genome-wide association studies detects candidate genes for wool traits by re-sequencing in Chinese fine-wool sheep. BMC Genom. 2021, 22, 127. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Zhu, S.; Guo, T.; Han, M.; Chen, B.; Qiao, G.; Wu, Y.; Yuan, C.; Liu, J.; Lu, Z.; et al. Whole-genome re-sequencing association study on yearling wool traits in Chinese fine-wool sheep. J. Anim. Sci. 2021, 99, skab210. [Google Scholar] [CrossRef]
- Jen, J.; Wang, Y. Zinc finger proteins in cancer progression. J. Biomed. Sci. 2016, 23, 53. [Google Scholar] [CrossRef]
- Cai, C.; Omwancha, J.; Hsieh, C.L.; Shemshedini, L. Androgen induces expression of the multidrug resistance protein gene MRP4 in prostate cancer cells. Prostate Cancer Prostatic Dis. 2007, 10, 39–45. [Google Scholar] [CrossRef]
- Cai, C.; Hsieh, C.L.; Gao, S.; Kannan, A.; Bhansali, M.; Govardhan, K.; Dutta, R.; Shemshedini, L. Soluble guanylyl cyclase α1 and p53 cytoplasmic sequestration and down-regulation in prostate cancer. Mol. Endocrinol. 2012, 26, 292–307. [Google Scholar] [CrossRef]
- Vassilev, L.T.; Vu, B.T.; Graves, B.; Carvajal, D.; Podlaski, F.; Filipovic, Z.; Kong, N.; Kammlott, U.; Lukacs, C.; Klein, C.; et al. In vivo activation of the p53 pathway by small-molecule antagonists of MDM2. Science 2004, 303, 844–848. [Google Scholar] [CrossRef]
- Piper, M.; Georgas, K.; Yamada, T.; Little, M. Expression of the vertebrate Slit gene family and their putative receptors, the Robo genes, in the developing murine kidney. Mech. Dev. 2000, 94, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ding, M. Robo and Ror function in a common receptor complex to regulate Wnt-mediated neurite outgrowth in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2018, 115, E2254–E2263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, C.; Zhang, C.; Peng, G. Wnt Signaling Regulates Ipsilateral Pathfinding in the Zebrafish Forebrain through slit3. Neuroscience 2020, 449, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Greaves, E.; Collins, F.; Esnal-Zufiaurre, A.; Giakoumelou, S.; Horne, A.W.; Saunders, P.T. Estrogen receptor (ER) agonists differentially regulate neuroangiogenesis in peritoneal endometriosis via the repellent factor SLIT3. Endocrinology 2014, 155, 4015–4026. [Google Scholar] [CrossRef]
- Zhang, B.; Dietrich, U.M.; Geng, J.G.; Bicknell, R.; Esko, J.D.; Wang, L. Repulsive axon guidance molecule Slit3 is a novel angiogenic factor. Blood 2009, 114, 4300–4309. [Google Scholar] [CrossRef]
- Yuan, W.; Rao, Y.; Babiuk, R.P.; Greer, J.J.; Wu, J.; Ornitz, D.M. A genetic model for a central (septum transversum) congenital diaphragmatic hernia in mice lacking Slit3. Proc. Natl. Acad. Sci. USA 2003, 100, 5217–5222. [Google Scholar] [CrossRef]
- Kenneth, J.; Livak, T.D.S. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar]
- Wang, Z.; Li, M.; Lan, X.; Li, M.; Lei, C.; Chen, H. Tetra-primer ARMS-PCR identifies the novel genetic variations of bovine HNF-4α gene associating with growth traits. Gene 2014, 546, 206–213. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Q.; Zhang, S.; Zhang, X.; Pan, C.; Chen, H.; Zhu, H.; Lan, X. Genetic Effects of Single Nucleotide Polymorphisms in the Goat GDF9 Gene on Prolificacy: True or False Positive? Animals 2019, 9, 886. [Google Scholar] [CrossRef]
- Jia, C. Genome-Wide Association Study for Growth and Serum Biochemical Parameters and Construction of CNV Map in Yak. Ph.D. Thesis, Northwest A&F University, Xianyang, China, 2020. [Google Scholar]
- Soemedi, R.; Cygan, K.J.; Rhine, C.L.; Wang, J.; Bulacan, C.; Yang, J.; Bayrak-Toydemir, P.; McDonald, J.; Fairbrother, W.G. Pathogenic variants that alter protein code often disrupt splicing. Nat. Genet. 2017, 49, 848–855. [Google Scholar] [CrossRef]
- Dybus, A.; Kulig, H.; Yu, Y.; Lanckriet, R.; Proskura, W.; Cheng, Y. CRY1 Gene Polymorphism and Racing Performance of Homing Pigeons. Animals 2021, 11, 2632. [Google Scholar] [CrossRef] [PubMed]
- Devran, Z.; Goknur, A.B.; Mesci, L. Development of molecular markers for the Mi-1 gene in tomato using the KASP genotyping assay. Hortic. Environ. Biotechnol. 2016, 57, 156–160. [Google Scholar] [CrossRef]
- Kusza, S.; Cziszter, L.T.; Ilie, D.E.; Sauer, M.; Padeanu, I.; Gavojdian, D. Kompetitive Allele Specific PCR (KASP™) genotyping of 48 polymorphisms at different caprine loci in French Alpine and Saanen goat breeds and their association with milk composition. PeerJ 2018, 6, e4416. [Google Scholar] [CrossRef]
- Feng, S.G.; Liu, S.; Yang, L. Keratin family and its regulation on wool growth and development. Chem. Life 2007, 154, 92–94. [Google Scholar]
- Gong, H.; Zhou, H.; McKenzie, G.W.; Yu, Z.; Clerens, S.; Dyer, J.M.; Plowman, J.E.; Wright, M.W.; Arora, R.; Bawden, C.S.; et al. An updated nomenclature for keratin-associated proteins (KAPs). Int. J. Biol. Sci. 2012, 8, 258. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| SNPs | Primer Sequence (5′-3′) | Reaction Component |
|---|---|---|
| F1: GAAGGTGACCAAGTTCATGCTCAGGAAAGCAGGGAAGCG | A-FAM | |
| SLIT3 g.478807C>G | F2: GAAGGTCGGAGTCAACGGATTCAGGAAAGCAGGGAAGCC | G-HEX |
| (chr16:945395) | R: GCTGGCTTTCTCTGATTCAGC | Common |
| F1: GAAGGTGACCAAGTTCATGCTGCATACATAGGGCATTTCGC | G-FAM | |
| ZNF280B g.677G>A | F2: GAAGGTCGGAGTCAACGGATTGGCATACATAGGGCATTTCGT | A-HEX |
| (chr17:70269373) | R: CTGCGGTCTGTAAAATCTGTGAA | Common |
| SNPs | Primer Sequence (5′-3′) | Product Size | Position |
|---|---|---|---|
| SLIT3 g.478807C>G | F: TTTCCCAGCAGGAGTGTCTA | 663 bp | Intron |
| R: GCCTTCTTGTTTCCGTTTCA | |||
| ZNF280B g.677G>A | F: TGGAACACTGGTGAAAACTC | 280 bp | Exon |
| R: CCCTCTGCGGTCTGTAAAAT |
| Gene | Primer Sequence (5′-3′) | Product Size |
|---|---|---|
| SLIT3 | F: CCTGGAGTGAAGAAGTGGGA | 320 bp |
| R: CATGTTCATCCTTTTGTTTT | ||
| ZNF280B | F: CAAATTCTTAGTGTTTCCATG | 247 bp |
| R: CCAGTTTCCCACCCCATTCC | ||
| GAPDH | F: AGAAGGCTGGGGCTCATTTG | 258 bp |
| R: AGGGGCCATCCACAGTCTTC |
| SNPs | Genotypic Frequencies | Allelic Frequencies | He | Ne | PIC | χ2 | |||
|---|---|---|---|---|---|---|---|---|---|
| SLIT3 g.478807C>G | CC(781) | CG(270) | GG(30) | C | G | 0.259 | 1.349 | 0.225 | 1.282 |
| 0.722 | 0.250 | 0.028 | 0.847 | 0.153 | |||||
| ZNF280B g.677G>A | GG(408) | GA(670) | AA(3) | G | A | 0.430 | 1.754 | 0.337 | 211.188 * |
| 0.377 | 0.620 | 0.003 | 0.687 | 0.313 | |||||
| SNPs | Genotype | Wool Fiber Diameter (μm) | F |
|---|---|---|---|
| SLIT3 g.478807C>G | CC | 19.8 ± 1.6 a | 2.613 |
| CG | 20.1 ± 1.7 b | ||
| GG | 19.9 ± 1.7 ab | ||
| ZNF280B g.677G>A | GG | 19.9 ± 1.6 a | 2.451 |
| GA | 19. 8 ± 1.6 a | ||
| AA | 17.9 ± 0.6 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, L.; Lu, Z.; Guo, T.; Liu, J.; Yuan, C.; Yang, B. Association of SLIT3 and ZNF280B Gene Polymorphisms with Wool Fiber Diameter. Animals 2023, 13, 3552. https://doi.org/10.3390/ani13223552
Yue L, Lu Z, Guo T, Liu J, Yuan C, Yang B. Association of SLIT3 and ZNF280B Gene Polymorphisms with Wool Fiber Diameter. Animals. 2023; 13(22):3552. https://doi.org/10.3390/ani13223552
Chicago/Turabian StyleYue, Lin, Zengkui Lu, Tingting Guo, Jianbin Liu, Chao Yuan, and Bohui Yang. 2023. "Association of SLIT3 and ZNF280B Gene Polymorphisms with Wool Fiber Diameter" Animals 13, no. 22: 3552. https://doi.org/10.3390/ani13223552
APA StyleYue, L., Lu, Z., Guo, T., Liu, J., Yuan, C., & Yang, B. (2023). Association of SLIT3 and ZNF280B Gene Polymorphisms with Wool Fiber Diameter. Animals, 13(22), 3552. https://doi.org/10.3390/ani13223552