
What Drives the Alien Parrot Richness and Occurrence in Urban Green Spaces along the Annual Cycle in Buenos Aires City, Argentina?


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Species Analized
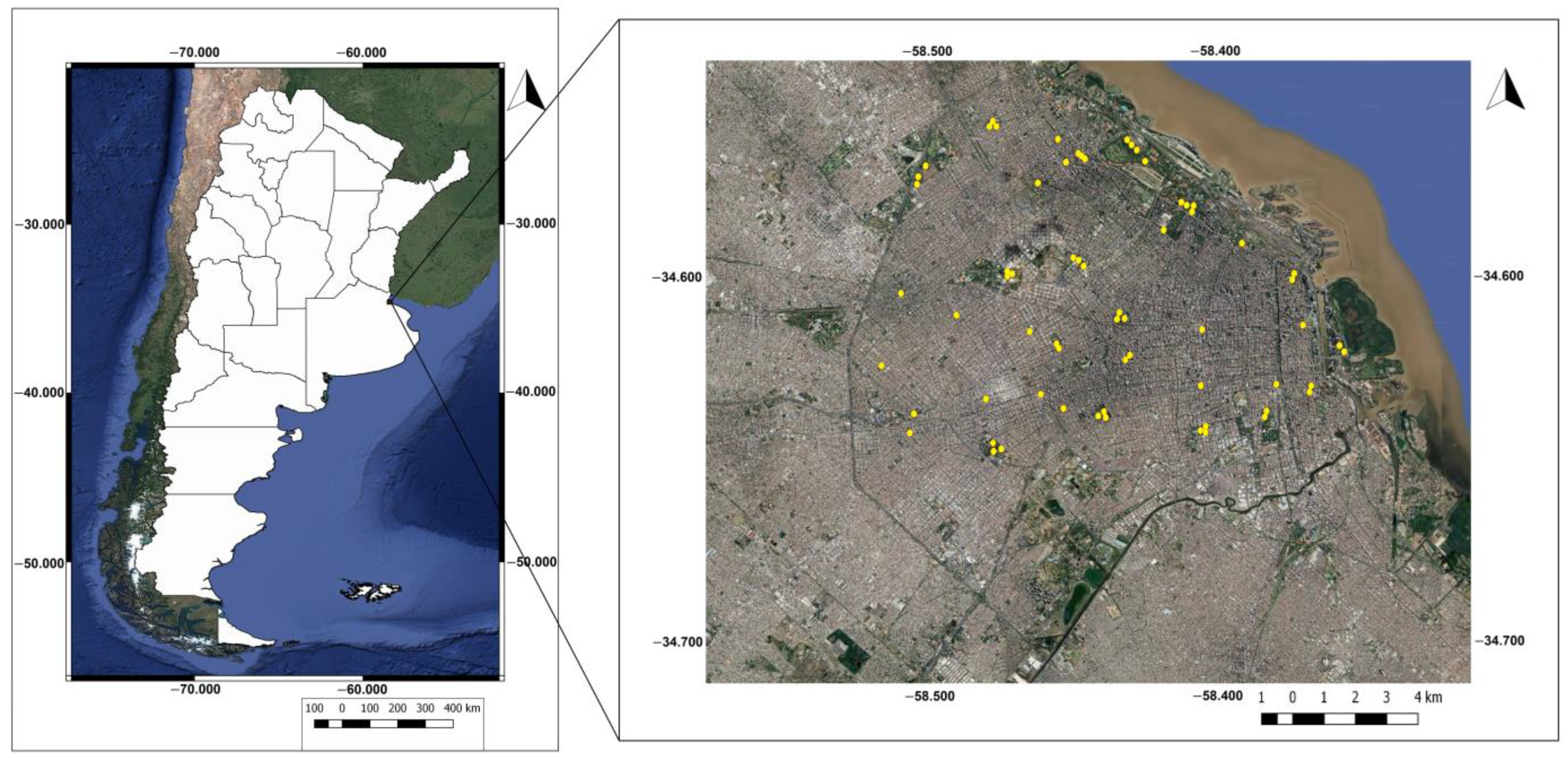
2.2. Study Area
2.3. Selection of Sampling Sites
2.4. Bird Counts
2.5. Environmental Variables
2.6. Variables of Parrot Diversity and Occurrence
2.7. Statistical Analysis
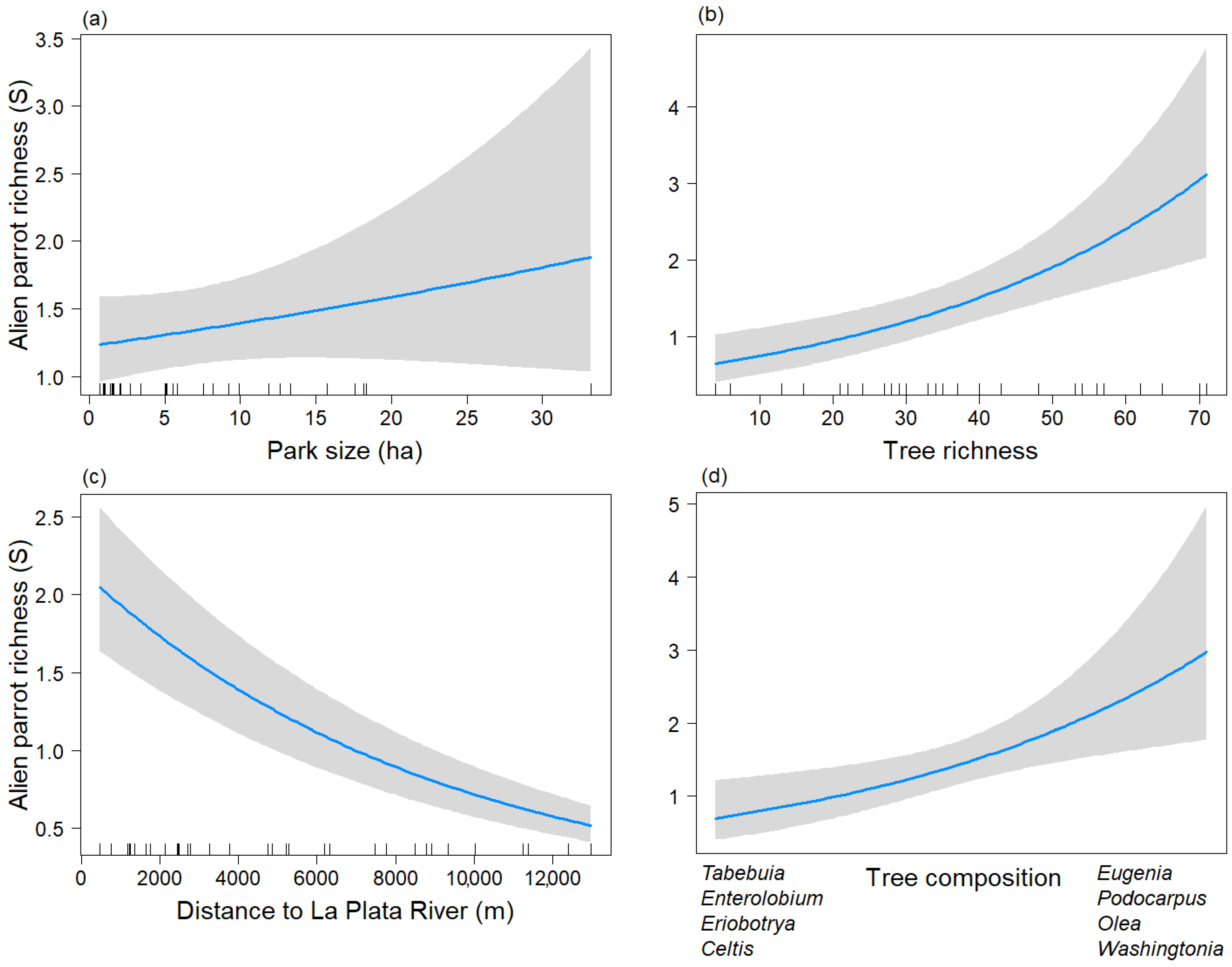
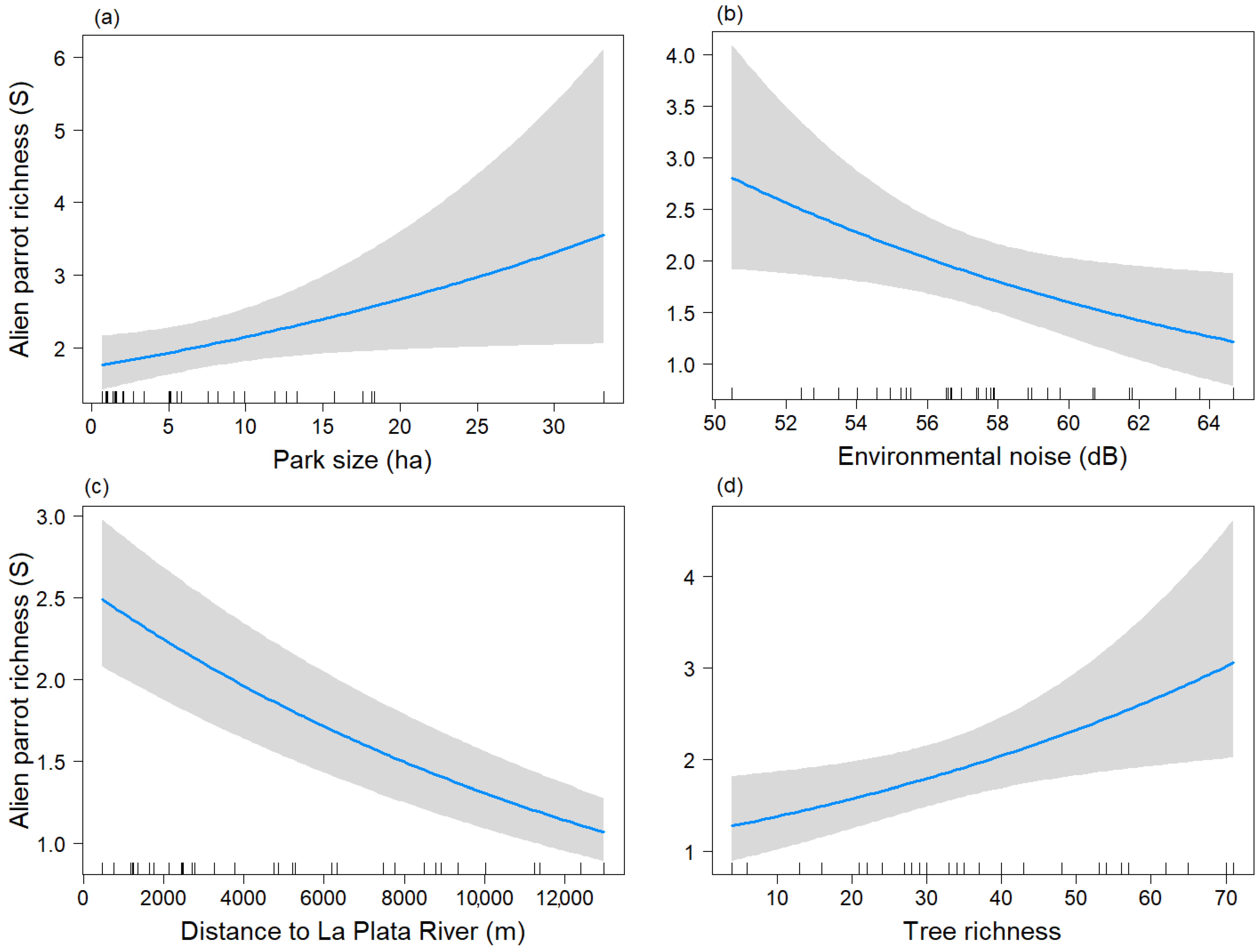
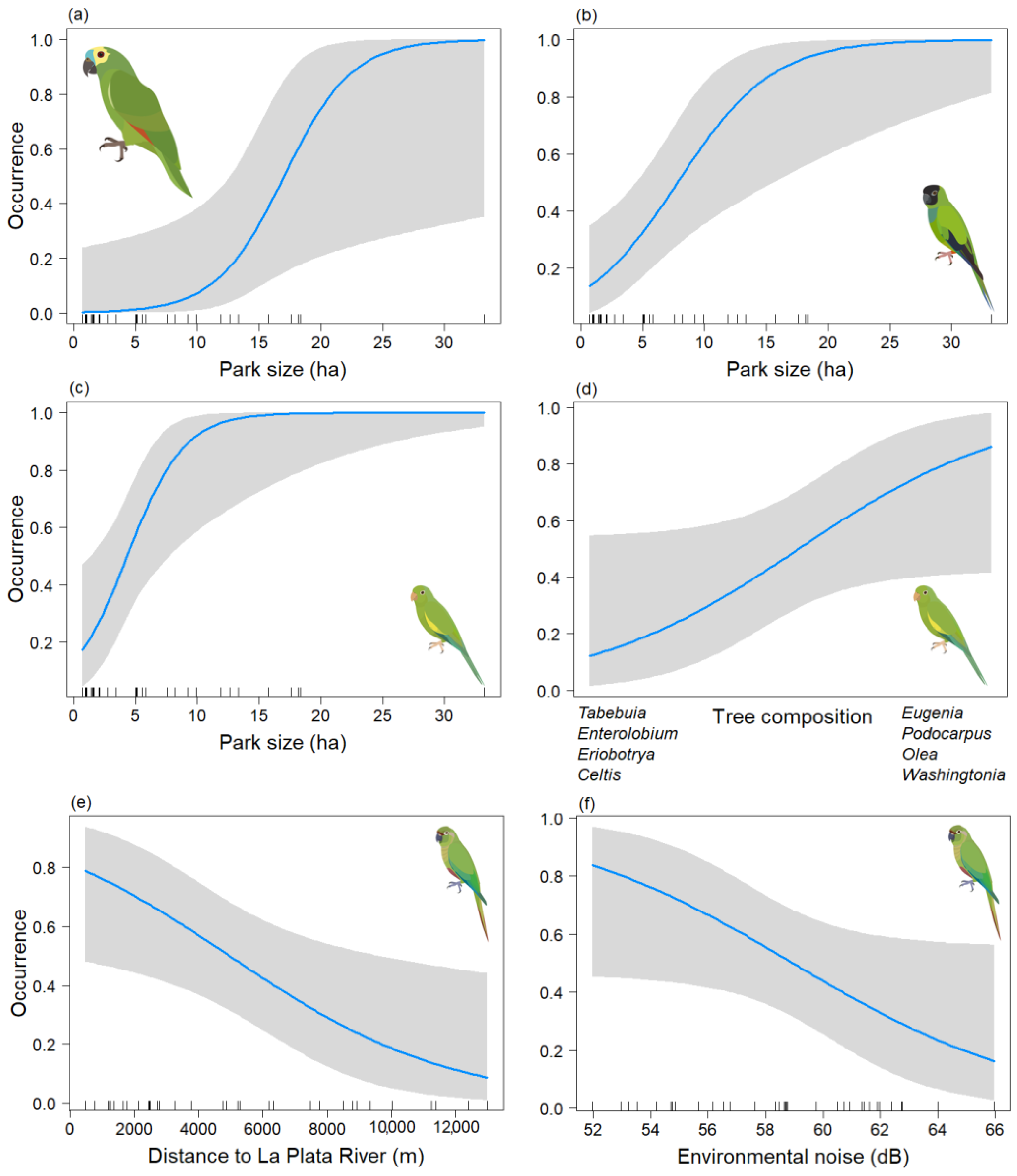
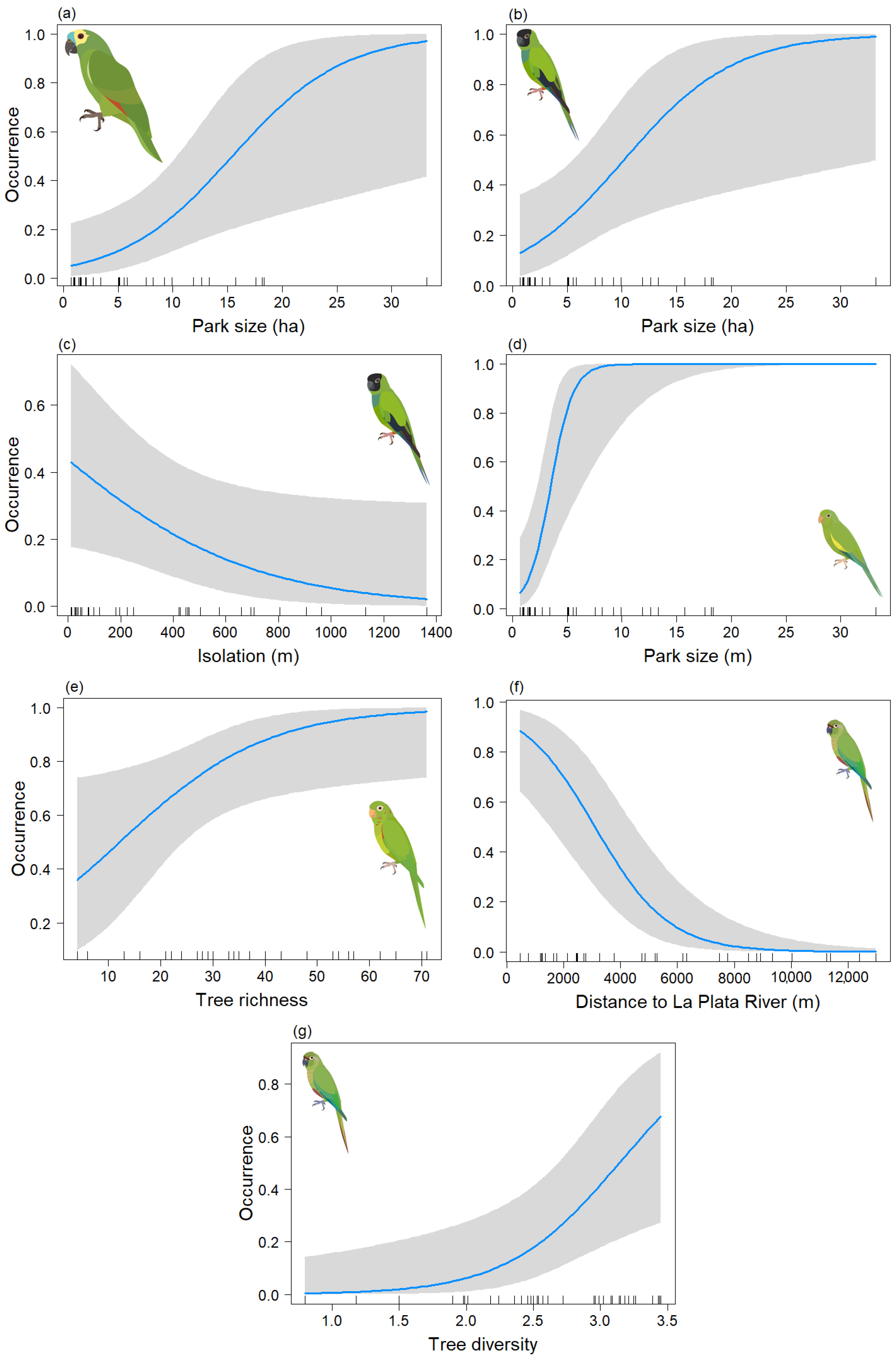
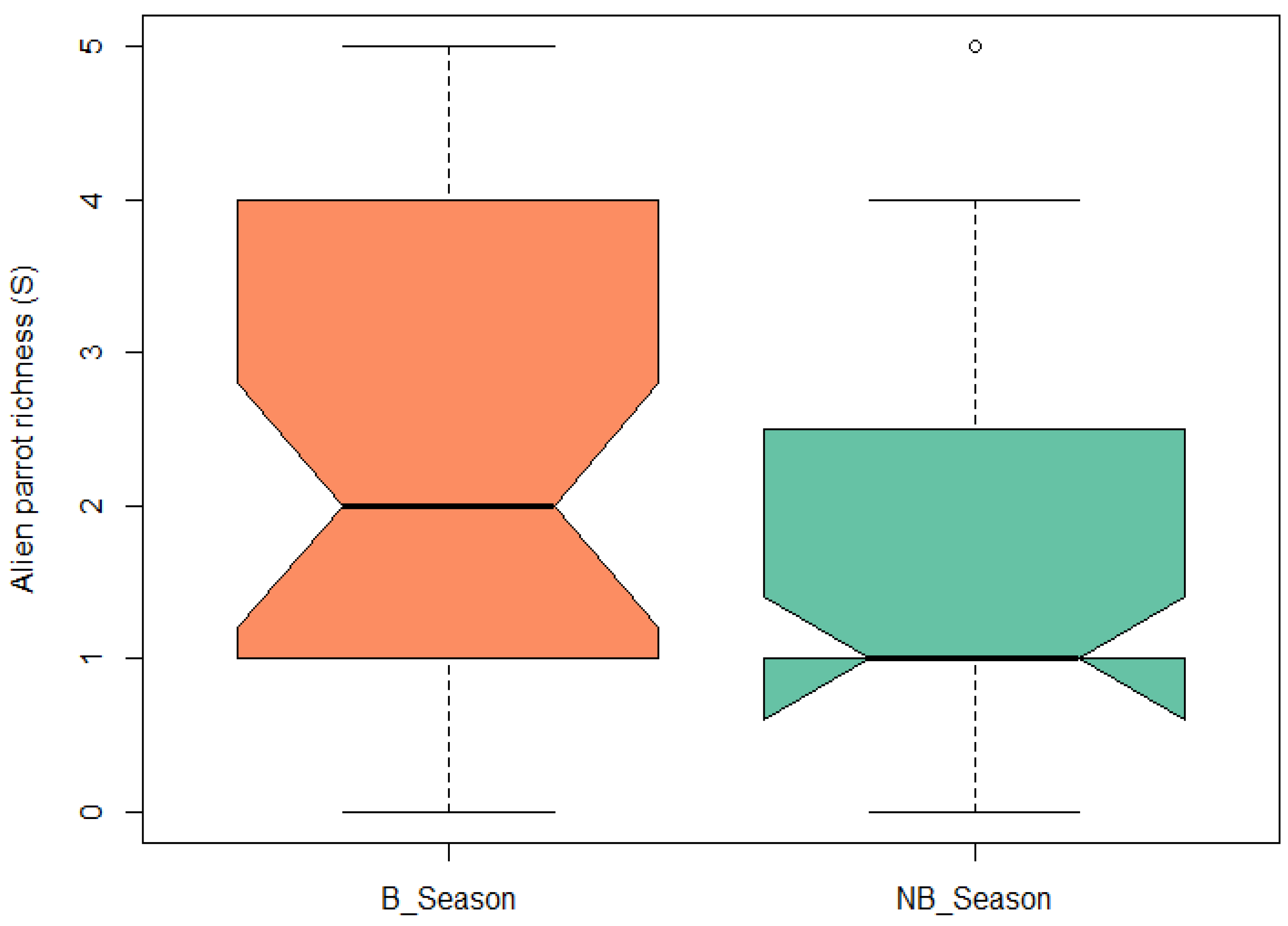
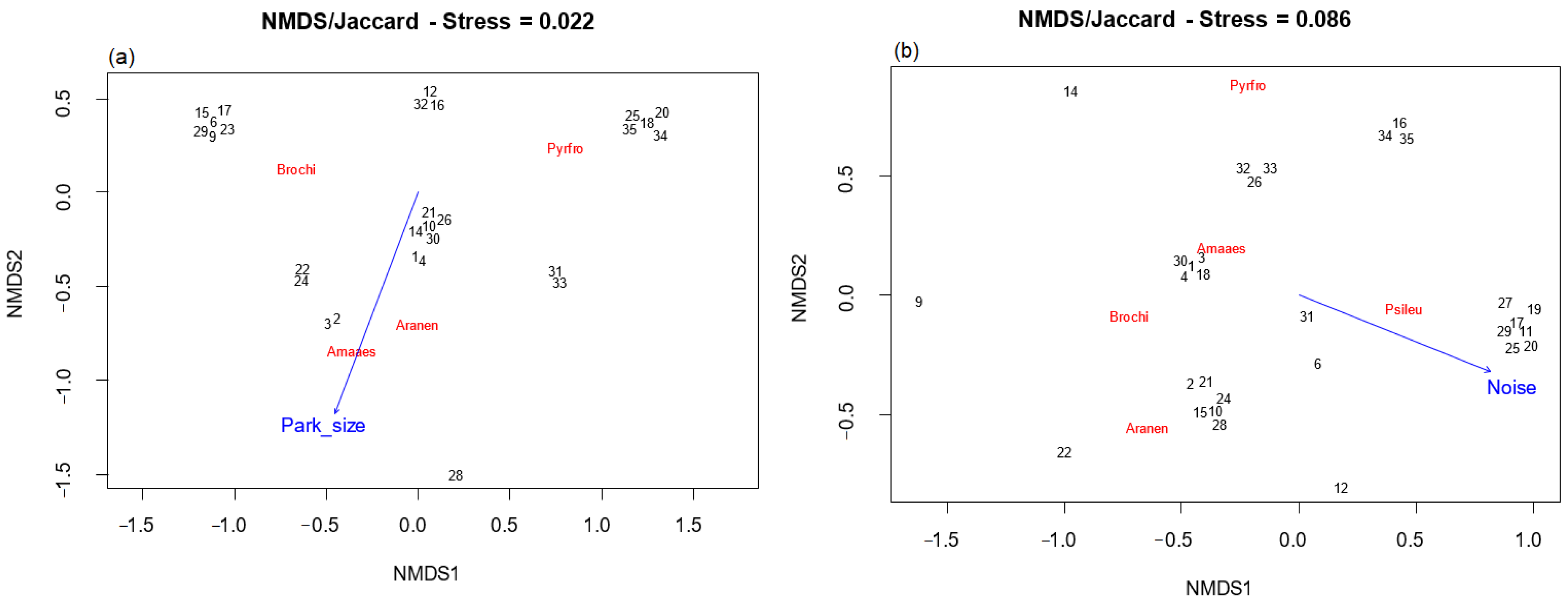
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blackburn, T.M.; Bellard, C.; Ricciardi, A. Alien versus native species as drivers of recent extinctions. Front. Ecol. Environ. 2019, 17, 203–207. [Google Scholar] [CrossRef]
- IUCN (International Union for Conservation of Nature). IUCN guidelines for the prevention of biodiversity loss caused by alien invasive species. In Proceedings of the 51st Meeting of the IUCN Council, Gland, Switzerland, 9 February 2000. [Google Scholar]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Carrete, M.; Tella, J. Wild-bird trade and exotic invasions: A new link of conservation concern? Front. Ecol. Environ. 2008, 6, 207–211. [Google Scholar] [CrossRef]
- Lockwood, J.L.; Cassey, P.; Blackburn, T.M. The more you introduce the more you get: The role of colonization pressure and propagule pressure in invasion ecology. Divers. Distrib. 2009, 15, 904–910. [Google Scholar] [CrossRef]
- Stanley, M.C.; McNaughton, E.J.; Fewster, R.M.; Galbraith, J.A. Cumulative propagule pressure exerted by escaped pet parrots. J. Appl. Ecol. 2023, 60, 384–392. [Google Scholar] [CrossRef]
- Peh, K.S.-H. Invasive species in Southeast Asia: The knowledge so far. Biodivers. Conserv. 2010, 19, 1083–1099. [Google Scholar] [CrossRef]
- Chan, D.T.C.; Poon, E.S.K.; Wong, A.T.C.; Sin, S.Y.W. Global trade in parrots–Influential factors of trade and implications for conservation. Glob. Ecol. Conserv. 2021, 30, e01784. [Google Scholar] [CrossRef]
- Bush, E.R.; Baker, S.E.; Macdonald, D.W. Global trade in exotic pets 2006–2012. Conserv. Biol. 2014, 28, 663–676. [Google Scholar] [CrossRef]
- Haene, E. Invasión de loros en la ciudad. Nat. Conserv. 2006, 19, 16–23. [Google Scholar]
- Cassey, P.; Blackburn, T.M.; Russell, G.J.; Jones, K.E.; Lockwood, J.L. Influences on the transport and establishment of exotic bird species: An analysis of the parrots (Psittaciformes) of the world. Glob. Chang. Biol. 2004, 10, 417–426. [Google Scholar] [CrossRef]
- Menchetti, M.; Mori, E. Worldwide impact of alien parrots. (Aves Psittaciformes) on native biodiversity and environment: A review. Ethol. Ecol. Evol. 2014, 26, 172–194. [Google Scholar] [CrossRef]
- Martin-Albarracin, V.L.; Amico, G.C.; Simberloff, D.; Nuñez, M.A. Impact of Non-Native Birds on Native Ecosystems: A Global Analysis. PLoS ONE 2015, 10, e0143070. [Google Scholar] [CrossRef] [PubMed]
- Strubbe, D.; Matthysen, E.; Graham, C.H. Assessing the potential impact of invasive ring-necked parakeets Psittacula krameri on native nuthatches Sitta europeae in Belgium. J. Appl. Ecol. 2010, 47, 549–557. [Google Scholar] [CrossRef]
- Strubbe, D.; Matthysen, E. Invasive ring-necked parakeets Psittacula krameri in Belgium: Habitat selection and impact on native birds. Ecography 2007, 30, 578–588. [Google Scholar]
- Hernández-Brito, D.; Carrete, M.; Ibáñez, C.; Juste, J.; Tella, J.L. Nest-site competition and killing by invasive parakeets cause the decline of a threatened bat population. R. Soc. Open Sci. 2018, 5, 172477. [Google Scholar] [CrossRef]
- Parr, M.; Juniper, T. Parrots: A Guide to Parrots of the World; Bloomsbury Publishing: London, UK, 2010. [Google Scholar]
- Francis, R.A.; Chadwick, M.A. Urban invasions: Non-native and invasive species in cities. Geography 2015, 100, 144–151. [Google Scholar] [CrossRef]
- Diamond, J.M.; Ross, M.S. Exotic parrots breeding in urban tree cavities: Nesting requirements, geographic distribution, and potential impacts on cavity nesting birds in southeast Florida. Avian Res. 2019, 10, 39. [Google Scholar] [CrossRef]
- Álvarez-Castillo, C.; MacGregor-Fors, I.; Arriaga-Weiss, S.L.; Mota-Vargas, C.; Santiago-Alarcon, D. Abundance of White-fronted Parrots and diet of an urban parrot assemblage (Aves: Psittaciformes) in a green Neotropical city. Avian Res. 2022, 13, 100019. [Google Scholar] [CrossRef]
- Senar, J.C.; Carrillo-Ortiz, J.G.; Ortega-Segalerva, A.; Dawson Pell, F.S.E.; Pascual, J.; Arroyo, L.; Mazzoni, D.; Montalvo, T.; Hatchwell, B.J. The reproductive capacity of Monk Parakeets Myiopsitta monachus is higher in their invasive range. Bird Study 2019, 66, 136–140. [Google Scholar] [CrossRef]
- Clergeau, P.; Vergnes, A. Bird feeders may sustain feral Rose-ringed parakeets Psittacula krameri in temperate Europe. Wildl. Biol. 2011, 17, 248–252. [Google Scholar] [CrossRef]
- Davis, A.; Taylor, C.E.; Major, R.E. Seasonal abundance and habitat use of Australian parrots in an urbanized landscape. Landsc Urban Plan 2012, 106, 191–198. [Google Scholar] [CrossRef]
- Leveau, L.M.; Ruggiero, A.; Matthews, T.J.; Bellocq, M.I. A global consistent positive effect of urban green area size on bird richness. Avian Res. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Barbosa, K.V.d.C.; Rodewald, A.D.; Ribeiro, M.C.; Jahn, A.E. Noise level and water distance drive resident and migratory bird species richness within a Neotropical megacity. Landsc. Urban Plan. 2020, 197, 103769. [Google Scholar] [CrossRef]
- Narosky, T.; Di Giacomo, A. Las Aves de la Provincia de Buenos Aires: Distribución y Estatus; Asociación Ornitológica del Plata, Vázquez Mazzini Editores y LOLA: Buenos Aires, Argentina, 1993. [Google Scholar]
- Ibañez, L.M.; Girini, J.M.; Palacio, F.X.; Montalti, D. Nidificación y alimentación de psittácidos introducidos en el noreste de la provincia de Buenos Aires, Argentina. Hornero 2014, 29, 13–22. [Google Scholar] [CrossRef]
- eBird. eBird Basic Dataset. Cornell Lab of Ornithology; Ithaca: New York, NY, USA, 2022. [Google Scholar]
- Narosky, T.; Henschke, C. Aves de la ciudad de Buenos Aires; Vásquez Mazzini Editores: Buenos Aires, Argentina, 2005. [Google Scholar]
- Cabrera, A.L. Regiones Fitogeográficas Argentinas; Acme: Buenos Aires, Argentina, 1976; p. 85. [Google Scholar]
- Narosky, T.; Yzurieta, D. Guía Para la Identificación de las aves de Argentina y Uruguay; Vazquez Mazzini: Buenos Aires, Argentina, 1987; p. 345. [Google Scholar]
- Peris, S.; Montelongo, T. Birds and small urban parks: A study in a high plateau city. Turk. J. Zool. 2014, 38, 316–325. [Google Scholar] [CrossRef]
- de Toledo, M.C.B.; Donatelli, R.J.; Batista, G.T. Relation between green spaces and bird community structure in an urban area in Southeast Brazil. Urban Ecosyst. 2012, 15, 111–131. [Google Scholar] [CrossRef]
- Urquiza, A.; Mella, J.E. Riqueza y diversidad de aves en parques de Santiago durante el período estival. Boletín Chil. De Ornitol. 2002, 9, 12–21. [Google Scholar]
- Melo, M.A.; Sanches, P.M.; Filho, D.F.S.; Piratelli, A.J. Influence of habitat type and distance from source area on bird taxonomic and functional diversity in a Neotropical megacity. Urban Ecosyst. 2022, 25, 545–560. [Google Scholar] [CrossRef]
- Faggi, A.; Perepelizin, P. Riqueza de aves a lo largo de un gradiente de urbanización en la ciudad de Buenos Aires. Rev. Del Mus. Argent. De Cienc. Nat. Nueva Ser. 2006, 8, 289–297. [Google Scholar] [CrossRef]
- Haene, E. Biocorredores de la Ciudad Autónoma de Buenos Aires, un Modelo Demostrativo Para la Argentina; Universidad de Belgrano: Buenos Aires, Argentina, 2020. [Google Scholar]
- Fitzsimons, J.A.; Palmer, G.C.; Antos, M.J.; White, J.G. Refugees and residents: Densities and habitat preferences of lorikeets in urban Melbourne. Aust. Field Ornithol. 2003, 20, 2–7. [Google Scholar]
- Shwartz, A.; Shirley, S.; Kark, S. How do habitat variability and management regime shape the spatial heterogeneity of birds within a large Mediterranean urban park? Landsc. Urban Plan. 2008, 84, 219–229. [Google Scholar] [CrossRef]
- Chávez Almonacid, C.A. Relación Entre la Avifauna, la Vegetación y las Construcciones en Plazas y Parques de la Ciudad de Valdivia. Doctoral Dissertation, Universidad Austral de Chile, Los Ríos, Chile, 2014. [Google Scholar]
- Curzel, F.E.; Bellocq, M.I.; Leveau, L.M. Local and landscape features of wooded streets influenced bird taxonomic and functional diversity. Urban For. Urban Green. 2021, 66, 127369. [Google Scholar] [CrossRef]
- Miller, J.R. Biodiversity conservation and the extinction of experience. Trends Ecol. Evol. 2005, 20, 430–434. [Google Scholar] [CrossRef] [PubMed]
- Leveau, L.M.; Leveau, C.M. Street design in suburban areas and its impact on bird communities: Considering different diversity facets over the year. Urban For. Urban Green. 2020, 48, 126578. [Google Scholar] [CrossRef]
- Dooling, R.J.; Popper, A.N. Some lessons from the effects of highway noise on birds. In Proceedings of the Fourth International Conference on the Effects of Noise on Aquatic Life, Dublin, Ireland, 10–16 July 2016; p. 010004. [Google Scholar]
- González-Oreja, J.A.; De La Fuente-Díaz-Ordaz, A.A.; Hernández-Santín, L.; Bonache-Regidor, C.; Buzo-Franco, D. Can human disturbance promote nestedness? Songbirds and noise in urban parks as a case study. Landsc. Urban Plan. 2012, 104, 9–18. [Google Scholar] [CrossRef]
- Leveau, L.M.; Leveau, C.M. Does urbanization affect the seasonal dynamics of bird communities in urban parks? Urban Ecosyst. 2016, 19, 631–647. [Google Scholar] [CrossRef]
- Hildén, O. Habitat selection in birds: A review. Ann. Zool. Fenn. 1965, 2, 53–75. [Google Scholar]
- Sagario, M.C.; Cueto, V.R. Seasonal Space use and Territory Size of Resident Sparrows in the Central Monte Desert, Argentina. Ardeola 2014, 61, 153–159. [Google Scholar] [CrossRef]
- BirdLife International. IUCN Red List for Birds. 2023. Available online: http://datazone.birdlife.org (accessed on 10 October 2023).
- Collar, N.; Kirwan, G.M.; Boesman, P.F.D. Turquoise-fronted Parrot (Amazona aestiva), version 1.0. In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Arellano, C. Nanday Parakeet (Aratinga nenday), version 1.0. In Birds of the World; Schulenberg, T.S., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Brightsmith, D.; Burgio, K.R.; Hiller, B.J.; Block, K.E.; Pyle, P.; Patten, M.A. Yellow-chevroned Parakeet (Brotogeris chiriri), version 1.0. In Birds of the World; Rodewald, P.G., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Collar, N.; Boesman, P.F.D.; Sharpe, C.J. White-eyed Parakeet (Psittacara leucophthalmus), version 1.0. In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Collar, N.; Boesman, P.F.D.; Kirwan, G.M. Maroon-bellied Parakeet (Pyrrhura frontalis), version 1.0. In Birds of the World; del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E., Eds.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Censo Nacional de Población, Hogares y Viviendas 2022. Resultados Provisionales; INDEC: Buenos Aires, Argentina, 2023.
- Soriano, A. Río de la Plata grasslands. In Natural Grasslands: Introduction and Western Hemisphere; Elsevier: Amsterdam, The Netherlands, 1992; pp. 367–407. [Google Scholar]
- Halffter, G.; Morello, J.; Matteuci, S.D.; Solbrig, O. La Biodiversidad y el uso de la tierra. Biodiversidad y uso de la Tierra: Conceptos y Ejemplos de Latinoamérica; Editorial Universitaria de Buenos Aires: Buenos Aires, Argentina, 1999; pp. 17–27. [Google Scholar]
- Morello, J.; Matteucci, S.D.; Rodriguez, A.F.; Silva, M.E.; Mesopotámica, P.; Llana, P. Ecorregiones y Complejos Eco-Sistémicos de Argentina; Orientación Gráfica Editora: Buenos Aires, Argentina, 2012. [Google Scholar]
- Morello, J.; Buzai, G.D.; Baxendale, C.A.; Rodríguez, A.F.; Matteucci, S.D.; Godagnone, R.E.; Casas, R.R. Urbanization and the consumption of fertile land and other ecological changes: The case of Buenos Aires. Environ. Urban. 2000, 12, 119–131. [Google Scholar] [CrossRef]
- Cavia, R.; Cueto, G.R.; Suárez, O.V. Changes in rodent communities according to the landscape structure in an urban ecosystem. Landsc. Urban Plan. 2009, 90, 11–19. [Google Scholar] [CrossRef]
- Gobierno de la Ciudad de Buenos Aires. 2011. Available online: https://data.buenosaires.gob.ar/dataset/ (accessed on 10 December 2021).
- González-García. Métodos para contar aves terrestres. Man. De Técnicas Para El Estud. De Fauna 2011, 1, 86–123. [Google Scholar]
- de la Peña, M.R. Nidos y reproducción de las aves argentinas. Ediciones Biol. Ser. Nat. Conserv. Soc. 2013, 8, 590. [Google Scholar]
- Xeno-canto. Xeno-canto: Sharing Bird Sounds from around the World. 2022. Available online: https://xeno-canto.org/ (accessed on 16 December 2021).
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Urbana, IL, USA, 1949. [Google Scholar]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. vegan: Community Ecology Package. R package version 2.5-7. 2022. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 8 January 2023).
- Zuur, A.F.; Ieno, E.N.; Smith, G.M. Analysing Ecological Data; Springer: New York, NY, USA, 2007; Volume 680. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: https://www.r-project.org/ (accessed on 4 January 2023).
- ToolsDev. Sound Meter–Decibel Meter & Noise Meter. 2016. Available online: https://play.google.com/store/apps/details?id=app.tools.soundmeter.decibel.noisedetector&hl=en_US (accessed on 10 December 2021).
- Dagnino, J. Coeficiente de correlación lineal de Pearson. Chil Anest 2014, 43, 150–153. [Google Scholar]
- Hartig, F.; Hartig, M.F. Package ‘DHARMa’. R package. 2017. Available online: https://cran.r-project.org/web/packages/DHARMa/index.html (accessed on 4 January 2023).
- Magnusson, A.; Skaug, H.; Nielsen, A.; Berg, C.; Kristensen, K.; Maechler, M.; Brooks, M.M. Package ‘Glmmtmb’. R Package Version 0.2.0. 2017. Available online: https://cran.r-project.org/web/packages/glmmTMB/index.html (accessed on 4 January 2023).
- Kristosch, G.C.; Marcondes-Machado, L.O. Diet and feeding behavior of the Reddish-bellied Parakeet (Pyrrhura frontalis) in an Araucaria forest in southeastern Brazil. Ornitol. Neotrop. 2001, 12, 215–223. [Google Scholar]
- Styche, A. Distribution and Behavioural Ecology of the Sulphur-Crested Cockatoo (Cacatua galerita L.) in New Zealand. Ph.D. Thesis, Victoria University of Wellington, Wellington, New Zealand, 2000. [Google Scholar]
- Cortes, J. Probable Nesting of the Blue-Crowned Conure Aratinga acuticaudata in Gibraltar. Bird Rep. 2006, 59–61. [Google Scholar]
- Paton, D.C.; Tucker, J.R.; Paton, J.B.; Paton, P.A. Avian vectors of the seeds of the European Olive Olea europaea. South Aust. Ornithol. 1988, 30, 158–159. [Google Scholar]
- Fernandez-Juricic, E.; Jokimäki, J. A habitat island approach to conserving birds in urban landscapes: Case studies from southern and northern Europe. Biodivers. Conserv. 2001, 10, 2023–2043. [Google Scholar] [CrossRef]
- Souza, R.C.H. Variação Temporal e Ocupação do Periquito-de-Asabranca Brotogeris versicolurus na área Urbana de Manaus, AM. Unpublished Master’s Dissertation, Instituto Nacional de Pesquisas da Amazônia, Manaus, Brazil, 2017. [Google Scholar]
- Renton, K. Seasonal variation in occurrence of macaws along a rainforest river. J. Field Ornithol. 2002, 73, 15–19. [Google Scholar] [CrossRef]
- Ragusa-Netto, J.; Fecchio, A. Plant food resources and the diet of a parrot community in a gallery forest of the southern Pantanal (Brazil). Braz. J. Biol. 2006, 66, 1021–1032. [Google Scholar] [CrossRef]
- Fragata, M.d.M.; Baccaro, F.; Gonçalves, A.L.S.; Borges, S.H. Living in a tropical concrete jungle: Diversity and abundance variation in a parrot assemblage (Aves, Psittacidae) of a major Amazonian city. Urban Ecosyst. 2022, 25, 977–987. [Google Scholar] [CrossRef]
- Geary, M.; Brailsford, C.J.; Hough, L.I.; Baker, F.; Guerrero, S.; Leon, Y.M.; Collar, N.J.; Marsden, S.J. Street-level green spaces support a key urban population of the threatened Hispaniolan parakeet Psittacara chloropterus. Urban Ecosyst. 2021, 24, 1371–1378. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Size (cm) | Native Range | Habitat Preferences | Diet | Sources |
|---|---|---|---|---|---|
| Amazona aestiva | 37 | Brazil, Bolivia, Paraguay, and N Argentina. | Wooded habitats, gallery forest, and palm groves. | Fruits, seeds, and flowers. | [50] |
| Aratinga nenday | 30 | Bolivia, Brazil, and N Argentina. | Gallery forest, deciduous forest, and palm stands. | Seeds, fruits, nuts, berries, and flowers. | [51] |
| Brotogeris chiriri | 23 | Bolivia, Brazil, Paraguay, and N Argentina. | Open woodlands, savannas, subtropical forest, gallery forest, and towns. | Seeds, nectar, fruits, and blossoms. | [52] |
| Psittacara leucophthalmus | 35 | Colombia, Ecuador, Perú, Brazil, Venezuela, Guianas, Bolivia, Paraguay, N Uruguay, and N Argentina. | Forest edge and adjacent savanna, scrub forest, secondary growth, deciduous or gallery woodland, palm groves, flooded forest, opening in rainforest, also mangroves. | Fruit, blossoms, grass seeds, and even insects. | [53] |
| Pyrrhura frontalis | 28 | Brazil, Paraguay, NE Argentina, and N Uruguay. | Humid and dry forest, gallery woodlands, partly cleared and thinned areas, and even cities. | Nuts, fruits, seeds, and flowers. | [54] |
| (A) Non-Breeding Season | |||
| Predictors | Estimates | CI | p-Value |
| Intercept | 1.33 | 1.07–1.65 | 0.009 |
| Park size | 1.10 | 0.93–1.30 | 0.272 |
| Distriv | 0.67 | 0.54–0.82 | <0.001 |
| S_tree | 1.54 | 1.23–1.93 | <0.001 |
| Tree_comp2 | 1.42 | 1.12–1.80 | 0.004 |
| (B) Breeding Season | |||
| Predictors | Estimates | CI | p-Value |
| Intercept | 1.95 | 1.64–2.32 | <0.001 |
| Park size | 1.17 | 1.00–1.37 | 0.051 |
| Distriv | 0.78 | 0.65–0.93 | 0.005 |
| S_tree | 1.27 | 1.05–1.55 | 0.016 |
| Db | 0.83 | 0.70–0.98 | 0.026 |
| Species | Predictors | Odds Ratios | CI | p-Value |
|---|---|---|---|---|
| (a) Am. aestiva | Intercept | 0.02 | 0.00–0.44 | 0.013 |
| Park size | 13.92 | 1.24–156.66 | 0.033 | |
| (b) Ar. nenday | Intercept | 0.73 | 0.31–0.74 | 0.479 |
| Park size | 6.51 | 1.74–24.32 | 0.005 | |
| (c) B. chiriri | Intercept | 2.82 | 0.83–9.58 | 0.096 |
| Park size | 23.41 | 2.16–253.80 | 0.010 | |
| Tree_comp2 | 2.49 | 0.96–6.44 | 0.060 | |
| (d) Py. frontalis | Intercept | 0.92 | 0.42–1.98 | 0.828 |
| Distriv | 0.34 | 0.14–0.85 | 0.021 | |
| Db | 0.44 | 0.19–1.03 | 0.058 |
| Species | Predictors | Estimates | CI | p-Value |
|---|---|---|---|---|
| (a) Am. aestiva | Intercept | 0.17 | 0.06–0.52 | 0.002 |
| Park size | 4.18 | 1.35–12.94 | 0.013 | |
| (b) Ar. nenday | Intercept | 0.52 | 0.21–1.29 | 0.157 |
| Park size | 4.24 | 1.25–14.36 | 0.020 | |
| Isolation | 0.39 | 0.14–1.03 | 0.058 | |
| (c) B. chiriri | Intercept | 20.80 | 1.07–404.09 | 0.045 |
| Park size | 1160.58 | 7.44–181,029.01 | 0.006 | |
| (d) Ps. leucophthalmus | Intercept | 5.05 | 1.70–14.97 | 0.004 |
| S_tree | 3.75 | 1.11–12.70 | 0.033 | |
| (e) Py. frontalis | Intercept | 0.19 | 0.05–0.79 | 0.022 |
| Distriv | 0.06 | 0.01–0.41 | 0.004 | |
| H_tree | 4.72 | 1.15–19.44 | 0.031 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santiago, S.M.; Paes Cavalcante, N.; Leveau, L.M. What Drives the Alien Parrot Richness and Occurrence in Urban Green Spaces along the Annual Cycle in Buenos Aires City, Argentina? Animals 2023, 13, 3426. https://doi.org/10.3390/ani13213426
Santiago SM, Paes Cavalcante N, Leveau LM. What Drives the Alien Parrot Richness and Occurrence in Urban Green Spaces along the Annual Cycle in Buenos Aires City, Argentina? Animals. 2023; 13(21):3426. https://doi.org/10.3390/ani13213426
Chicago/Turabian StyleSantiago, Sebastián Martín, Nínive Paes Cavalcante, and Lucas Matías Leveau. 2023. "What Drives the Alien Parrot Richness and Occurrence in Urban Green Spaces along the Annual Cycle in Buenos Aires City, Argentina?" Animals 13, no. 21: 3426. https://doi.org/10.3390/ani13213426
APA StyleSantiago, S. M., Paes Cavalcante, N., & Leveau, L. M. (2023). What Drives the Alien Parrot Richness and Occurrence in Urban Green Spaces along the Annual Cycle in Buenos Aires City, Argentina? Animals, 13(21), 3426. https://doi.org/10.3390/ani13213426