
Behavioral and Physiological Differences in Female Rabbits at Different Stages of the Estrous Cycle

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
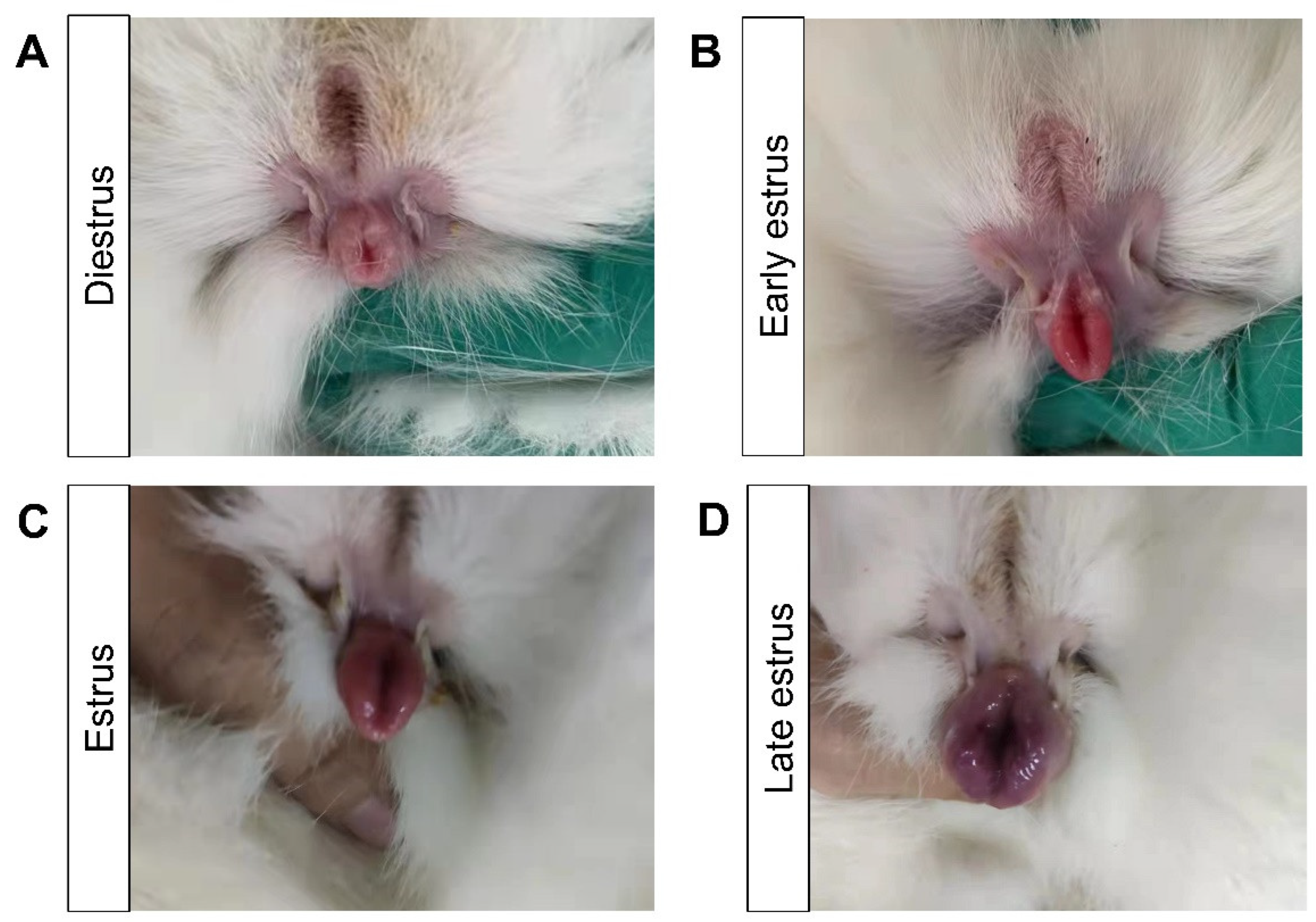
2.2. Estrus Identification
2.3. Behavior Observation and Recording
2.4. Plasma Collection and Reproductive Hormone Detection
2.5. Hematoxylin and Eosin (HE) Staining, and Follicle Counts
2.6. RNA-Seq Analysis
2.7. Real-Time Quantitative PCR (RT-qPCR)
2.8. Statistical Analysis
3. Results
3.1. Behavioral Differences in Female Rabbits at Different Stages of the Estrous Cycle
3.2. Hormonal Differences in Female Rabbits at Different Stages of the Estrous Cycle
3.3. Follicle Development Differences in Female Rabbits at Different Stages of the Estrous Cycle
3.4. Screening of Ovarian Transcription Patterns in Female Rabbits in Diestrus and Estrus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ola, S.I.; Oyegbade, M.O. Buck effect on rabbit oestrous: Vulva colour, vaginal lumen cells and ovarian follicle populations. World Rabbit Sci. 2012, 20, 71–79. [Google Scholar] [CrossRef]
- Hoffman, K.L.; González-Mariscal, G. Progesterone receptor activation signals behavioral transitions across the reproductive cycle of the female rabbit. Horm. Behav. 2006, 50, 154–168. [Google Scholar] [CrossRef]
- Gonzalez-Mariscal, G. Neuroendocrinology of maternal behavior in the rabbit. Horm. Behav. 2001, 40, 125–132. [Google Scholar] [CrossRef]
- Beyer, C.; Hoffman, K.L.; Gonzalez-Flores, O. Neuroendocrine regulation of estrous behavior in the rabbit: Similarities and differences with the rat. Horm. Behav. 2007, 52, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.U.; Wu, Y.B.; Ruan, G.C.; Wang, X.L. Environmental, Behavioral and Health Issues of Weaning and Finishing Pigs under Intensive Production System. In Proceedings of the International Livestock Environment Symposium, Beijing, China, 18–20 May 2005. [Google Scholar]
- Lopez, H.; Sartori, R.; Wiltbank, M.C. Reproductive hormones and follicular growth during development of one or multiple dominant follicles in cattle. Biol. Reprod. 2005, 72, 788–795. [Google Scholar] [CrossRef]
- Ferreira, A.C.A.; Cadenas, J.; Sá, N.A.R.; Correia, H.H.V.; Guerreiro, D.D.; Lobo, C.H.; Alves, B.G.; Maside, C.; Gastal, E.L.; Rodrigues, A.P.R.; et al. In vitro culture of isolated preantral and antral follicles of goats using human recombinant FSH: Concentration-dependent and stage-specific effect. Anim. Reprod. Sci. 2018, 196, 120–129. [Google Scholar] [CrossRef]
- Setty, S.L.; Mills, T.M. The effects of progesterone on follicular growth in the rabbit ovary. Biol. Reprod. 1987, 36, 1247–1252. [Google Scholar] [CrossRef] [PubMed]
- Cortvrindt, R.; Hu, Y.; Smitz, J. Recombinant luteinizing hormone as a survival and differentiation factor increases oocyte maturation in recombinant follicle stimulating hormone-supplemented mouse preantral follicle culture. Human Reprod. 1998, 13, 1292–1302. [Google Scholar] [CrossRef]
- Flaws, J.A.; Abbud, R.; Mann, R.J.; Nilson, J.H.; Hirshfield, A.N. Chronically elevated luteinizing hormone depletes primordial follicles in the mouse ovary. Biol. Reprod. 1997, 57, 1233–1237. [Google Scholar] [CrossRef] [PubMed]
- Hillier, S.G. Gonadotropic control of ovarian follicular growth and development. Mol. Cell. Endocrinol. 2001, 179, 39–46. [Google Scholar] [CrossRef]
- Paradis, F.; Novak, S.; Murdoch, G.K.; Dyck, M.K.; Dixon, W.T.; Foxcroft, G.R. Temporal regulation of BMP2, BMP6, BMP15, GDF9, BMPR1A, BMPR1B, BMPR2 and TGFBR1 mRNA expression in the oocyte, granulosa and theca cells of developing preovulatory follicles in the pig. Reproduction 2009, 138, 115–129. [Google Scholar] [CrossRef]
- Cauet, G.; Balbuena, D.; Achstetter, T.; Dumas, B. CYP11A1 stimulates the hydroxylase activity of CYP11B1 in mitochondria of recombinant yeast in vivo and in vitro. Eur. J. Biochem. 2001, 268, 4054–4062. [Google Scholar] [CrossRef]
- Luo, W.X.; Wiltbank, M.C. Distinct regulation by steroids of messenger RNAs for FSHR and CYP19A1 in bovine granulosa cells. Biol. Reprod. 2006, 75, 217–225. [Google Scholar] [CrossRef]
- Chaube, R.; Rawat, A.; Inbaraj, R.M.; Joy, K.P. Cloning and characterization of estrogen hydroxylase (cyp1a1 and cyp1b1) genes in the stinging catfish Heteropneustes fossilis and induction of mRNA expression during final oocyte maturation. Comp. Biochem. Physiol. A—Mol. Integr. Physiol. 2021, 253, 14. [Google Scholar] [CrossRef] [PubMed]
- Shen-Jin, L.U.; Yan, Y. Activity rhythm of mature female small-tailed Han sheep in household condition in spring and autumn season. Chin. J. Vet. Sci. 2012, 32, 1324–1328. [Google Scholar]
- Aguilar-Roblero, R.; González-Mariscal, G. Behavioral, neuroendocrine and physiological indicators of the circadian biology of male and female rabbits. Eur. J. Neuroscip. 2020, 51, 429–453. [Google Scholar] [CrossRef]
- Součková, M.; Přibylová, L.; Jurčová, L.; Chaloupková, H. Behavioural reactions of rabbits during AAI sessions. Appl. Anim. Behav. Sci. 2023, 262, 105908. [Google Scholar] [CrossRef]
- Zhuandan, Y. The observation on the behavior of wool rabbit. J. Shanghai Jiaotong Univ. 1986, 4, 253–262. [Google Scholar]
- Li, F.K.; Yang, Y.; Jenna, K.; Xia, C.H.; Lv, S.J.; Wei, W.H. Effect of heat stress on the behavioral and physiological patterns of Small-tail Han sheep housed indoors. Trop. Anim. Health Prod. 2018, 50, 1893–1901. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.R. The Behavior and Social Preferences of New Zealand White Rabbits; University of California: Davis, CA, USA, 2003. [Google Scholar]
- Beltrán, J.F.; Delibes, M. Environmental Determinants of Circadian Activity of Free-Ranging Iberian Lynxes. J. Mammal. 1994, 75, 382–393. [Google Scholar] [CrossRef]
- Arts, K.J.; Hudson, M.K.; Sharp, N.W.; Edelman, A.J. Eastern Spotted Skunks Alter Nightly Activity and Movement in Response to Environmental Conditions. Am. Midl. Nat. 2022, 188, 33–55. [Google Scholar] [CrossRef]
- Peng, X.U.; Yankuo, L.I.; Tao, M.; Yan, H. Daytime Allocation and Behavior Rhythm of Sichuan Snub-nosed Monkey in the Qingmuchuan Nature Reserve. Jiangxi Sci. 2015, 33, 324–329. [Google Scholar]
- Forbes, J.M. The Effects of Sex Hormones, Pregnancy and Lactation on Digestion, Metabolism and Voluntary Food Intake. In Control of Digestion and Metabolism in Ruminants; Reston Pub Co.: Reston, VA, USA, 1986; pp. 420–435. [Google Scholar]
- Ingvartsen, K.L.; Andersen, J.B. Integration of metabolism and intake regulation: A review focusing on periparturient animals. J. Dairy Sci. 2000, 83, 1573–1597. [Google Scholar] [CrossRef] [PubMed]
- Dybkjaer, L.; Jacobsen, A.P.; Togersen, F.A.; Poulsen, H.D. Eating and drinking activity of newly weaned piglets: Effects of individual characteristics, social mixing, and addition of extra zinc to the feed. J. Anim. Sci. 2006, 84, 702–711. [Google Scholar] [CrossRef]
- Kumar, A.; Mehta, J.S.; Ruhil, S.; Purohit, G.N. Effects of different estrous synchronization protocols on estrus and subsequent fertility in cycling cows. Rumin. Sci. 2019, 7, 83–86. [Google Scholar]
- Li, F.; Yang, Y.; Wang, H.; Pan, Z.; Wang, Z.; Lv, S. Behavioral and physiological difference of small-tailed Han sheep at different estrus stage after delivery. J. Vet. Behav. 2023, 60, 44–50. [Google Scholar] [CrossRef]
- Mora, S.; Dussaubat, N.; Díaz-Véliz, G. Effects of the estrous cycle and ovarian hormones on behavioral indices of anxiety in female rats. Psychoneuroendocrinology 1996, 21, 609–620. [Google Scholar] [CrossRef]
- Frye, C.A.; Petralia, S.M.; Rhodes, M.E. Estrous cycle and sex differences in performance on anxiety tasks coincide with increases in hippocampal progesterone and 3α,5α-THP. Pharmacol. Biochem. Behav. 2000, 67, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Mihm, A.; Bleach, E.C.L. Endocrine regulation of ovarian antral follicle development in cattle. Anim. Reprod. Sci. 2003, 78, 217–237. [Google Scholar] [CrossRef]
- Arat, O.; Deveci, D.; Ozkan, Z.S.; Tuncer Can, S. What is the effect of the early follicular phase FSH/LH ratio on the number of mature oocytes and embryo development? Turk. J. Med. Sci. 2020, 50, 420–425. [Google Scholar] [CrossRef]
- Eski, F.; Kurt, S.; Demir, P.A. Effect of different estrus synchronization protocols on estrus and pregnancy rates, oxidative stress and some biochemical parameters in Hair goats. Small Ruminant Res. 2021, 198, 6. [Google Scholar] [CrossRef]
- Widyastuti, R.; Boediono, A.; Syamsunarno, M.; Ghozali, M.; Sudiman, J. Effect of short-term gavage of ethanolic extract of cogon grass (Imperata cylindrica L) root on the ovarian activity and estrus behavior of female mice. Asian Pac. J. Reprod. 2020, 9, 70–76. [Google Scholar]
- Edson, M.A.; Nalam, R.L.; Clementi, C.; Franco, H.L.; DeMayo, F.J.; Lyons, K.M.; Pangas, S.A.; Matzuk, M.M. Granulosa Cell-Expressed BMPR1A and BMPR1B Have Unique Functions in Regulating Fertility but Act Redundantly to Suppress Ovarian Tumor Development. Mol. Endocrinol. 2010, 24, 1251–1266. [Google Scholar] [CrossRef] [PubMed]
- Shimasaki, S.; Moore, R.K.; Otsuka, F.; Erickson, G.F. The bone morphogenetic protein system in mammalian reproduction. Endocr. Rev. 2004, 25, 72–101. [Google Scholar] [CrossRef] [PubMed]
- Dongyong, Y.; Xiao, Y.; Fangfang, D.; Yanqing, W.; Yi, Y.; Min, H.; Yanxiang, C. The Role of Bone Morphogenetic Protein 4 in Ovarian Function and Diseases. Reprod. Sci. 2021, 28, 3316–3330. [Google Scholar] [CrossRef]
- Burghardt, R.C.; Anderson, E. Hormonal modulation of gap junctions in rat ovarian follicles. Cell Tissue Res. 1981, 214, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Takeuchi, H.; Yamaoka, M.; Nakanishi, T.; Tonai, S.; Nishimura, R.; Morita, T.; Nagano, M.; Kameda, S.; Genda, K.; et al. Lipopolysaccharide (LPS) suppresses follicle development marker expression and enhances cytokine expressions, which results in fail to granulosa cell proliferation in developing follicle in cows. Reprod. Biol. 2023, 23, 100710. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior | Specific Description |
|---|---|
| Relaxing [16] | Lying on the bottom of the cage, limbs hidden under the body; or lying on the bottom, limbs spread out |
| Locomotion [17] | Moving the body, including running and jumping |
| Grooming [18] | Licking the fur and limbs and cleaning the face with the forelimbs |
| Foraging [16] | Bending the head close to the trough, and making a chewing motion after raising the head |
| Drinking [19] | Raising their head after approaching the water bottle and swallowing it |
| Standing [20] | Sitting and staying still without any purpose |
| Observing [16] | Standing on all fours, keeping the body still, and raising the head to look out of the cage or the next cage |
| Biting [21] | Gnawing hard objects, such as huts, sinks, and troughs |
| Gene | Primer Sequence (5′ → 3′) | Accession Number | Tm (°C) | Product Size (bp) |
|---|---|---|---|---|
| CYP1B1 | F: GCTGGGAACTGACTCCACTC | XM_002709757.4 | 60 | 143 |
| R: CAAGAACGCTTGGCTAGGGA | ||||
| CYP19A1 | F: CCTGTCGTGGACTTGGTCAT | NM_001170921 | 60 | 279 |
| R: CACCTGGAATCGTCTCAGCA | ||||
| BMPR1B | F: GCAGTGGATGTGCCTTGTAT | XM_017347401.2 | 58 | 181 |
| R: CTCTTTCTTGGTGCCCACAT | ||||
| HOXA9 | F: CCCCATCGACCCCAATAACC | NM_001171403.1 | 60 | 240 |
| R: GCCCGGTCCTTGTTGATCTT | ||||
| IGF1R | F: GAGGAAGCGGAGAGATGTCG | XM_051822976.1 | 60 | 251 |
| R: CAAAGTTGGAGGCACTGCAC | ||||
| ESR1 | F: TGCTACGAAGTGGGAATGATGA | XM_051854226.1 | 60 | 273 |
| R: GGGTCTGGTAGGGTCGTACT | ||||
| GAPDH | F: CACCAGGGCTGCTTTTAACTCT | NM_001082253.1 | 60 | 145 |
| R: CTTCCCGTTCTCAGCCTTGACC |
| Behavior | Estrus Stage | |||
|---|---|---|---|---|
| Diestrus | Early Estrus | Estrus | Late Estrus | |
| Relaxing | 845.50 ± 84.07 | 831.68 ± 130.06 | 819.90 ± 91.15 | 829.60 ± 119.96 |
| Grooming | 192.06 ± 47.91 Bb | 219.00 ± 54.65 ABb | 291.30 ± 13.60 Aa | 249.60 ± 39.65 ABab |
| Foraging | 171.42 ± 45.18 Aa | 133.06 ± 36.56 ABab | 98.86 ± 25.54 Bb | 105.40 ± 31.83 ABb |
| Drinking | 31.44 ± 13.64 Aa | 25.46 ± 8.61 ABab | 17.48 ± 6.81 ABb | 13.34 ± 5.74 Bb |
| Locomotion | 53.66 ± 38.27 | 47.10 ± 37.32 | 57.22 ± 27.03 | 49.74 ± 29.99 |
| Standing | 23.10 ± 14.56 | 11.18 ± 5.01 | 15.50 ± 6.88 | 19.26 ± 17.03 |
| Observing | 20.20 ± 17.37 | 22.80 ± 14.28 | 26.08 ± 10.42 | 20.86 ± 10.43 |
| Biting | 10.18 ± 3.32 | 8.86 ± 3.48 | 16.05 ± 12.07 | 9.17 ± 1.46 |
| Behavior | Estrus Stage | |||
|---|---|---|---|---|
| Diestrus | Early Estrus | Estrus | Late Estrus | |
| Relaxing | 99.00 ± 21.01 b | 103.40 ± 32.97 b | 139.80 ± 52.44 a | 114.80 ± 45.04 ab |
| Grooming | 97.80 ± 15.55 Bb | 124.00 ± 39.51 ABb | 177.60 ± 23.97 Aa | 132.60 ± 45.08 ABab |
| Foraging | 63.40 ± 12.50 | 56.80 ± 22.81 | 57.60 ± 7.57 | 55.20 ± 9.39 |
| Drinking | 40.40 ± 11.41 ab | 42.40 ± 10.92 a | 29.40 ± 11.55 ab | 26.80 ± 7.66 b |
| Locomotion | 37.00 ± 10.56 b | 44.50 ± 21.06 b | 74.75 ± 18.79 a | 36.75 ± 12.31 b |
| Standing | 28.25 ± 10.01 | 43.33 ± 19.22 | 28.75 ± 7.85 | 29.25 ± 11.53 |
| Observing | 19.75 ± 11.76 | 20.60 ± 7.09 | 30.80 ± 14.72 | 17.80 ± 6.72 |
| Biting | 10.00 ± 3.61 Bc | 18.33 ± 1.53 Bb | 34.00 ± 5.66 Aa | 23.50 ± 0.71 Ab |
| Item | Estrus Stage | |||
|---|---|---|---|---|
| Diestrus | Early Estrus | Estrus | Late Estrus | |
| FSH, mIU/mL | 9.16 ± 0.52 ab | 9.48 ± 0.48 a | 9.06 ± 0.83 ab | 8.75 ± 0.52 b |
| LH, ng/mL | 50.37 ± 1.62 b | 52.33 ± 1.41 a | 52.83 ± 0.72 a | 48.97 ± 2.27 b |
| P4, ng/mL | 8.67 ± 0.51 a | 8.36 ± 0.53 ab | 7.94 ± 0.57 b | 8.75 ± 0.66 a |
| E2, pg/mL | 537.44 ± 36.65 ab | 524.99 ± 33.51 b | 554.07 ± 53.20 ab | 565.95 ± 46.14 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Jin, R.; Yang, A.; Li, J.; Song, Y.; Zhao, B.; Chen, Y.; Wu, X. Behavioral and Physiological Differences in Female Rabbits at Different Stages of the Estrous Cycle. Animals 2023, 13, 3414. https://doi.org/10.3390/ani13213414
Chen X, Jin R, Yang A, Li J, Song Y, Zhao B, Chen Y, Wu X. Behavioral and Physiological Differences in Female Rabbits at Different Stages of the Estrous Cycle. Animals. 2023; 13(21):3414. https://doi.org/10.3390/ani13213414
Chicago/Turabian StyleChen, Xin, Rongshuai Jin, Anqi Yang, Jiacheng Li, Ying Song, Bohao Zhao, Yang Chen, and Xinsheng Wu. 2023. "Behavioral and Physiological Differences in Female Rabbits at Different Stages of the Estrous Cycle" Animals 13, no. 21: 3414. https://doi.org/10.3390/ani13213414
APA StyleChen, X., Jin, R., Yang, A., Li, J., Song, Y., Zhao, B., Chen, Y., & Wu, X. (2023). Behavioral and Physiological Differences in Female Rabbits at Different Stages of the Estrous Cycle. Animals, 13(21), 3414. https://doi.org/10.3390/ani13213414