

Effect of Hermetia illucens Fat, Compared with That of Soybean Oil and Palm Oil, on Hepatic Lipid Metabolism and Plasma Metabolome in Healthy Rats

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Sample Collection
2.3. Determination of AID of Total Fat
2.4. Determination of Total Lipid Concentration
2.5. Histological Evaluation of Liver Lipid Accumulation
2.6. Determination of TG and Cholesterol Concentrations in Plasma and Liver
2.7. Total RNA Isolation and qPCR Analysis
2.8. Targeted Plasma Metabolomics
2.9. Statistical Analysis
3. Results
3.1. Body Weight Development, Feed Intake, Organ Weights, and AID of Total Fat
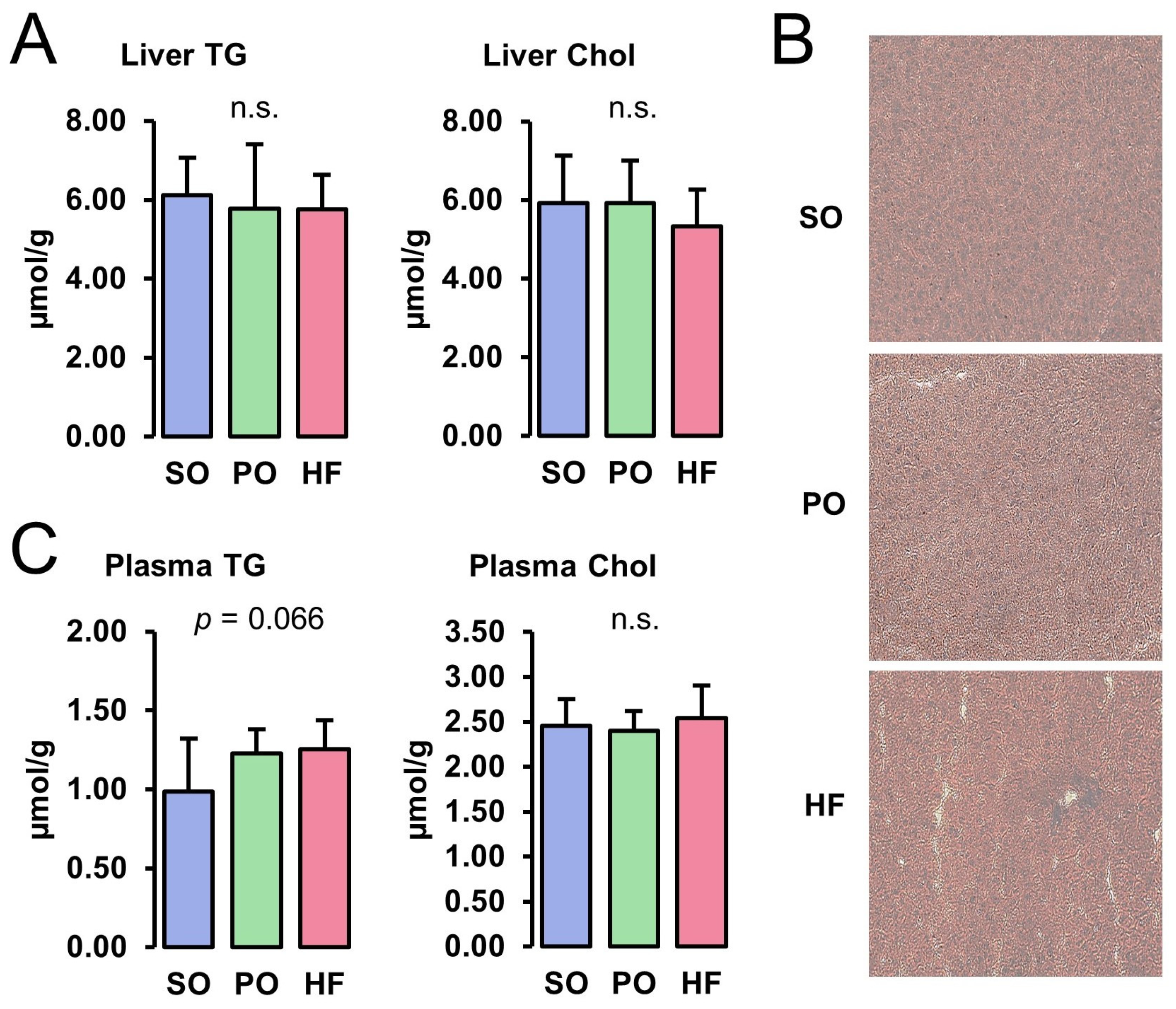
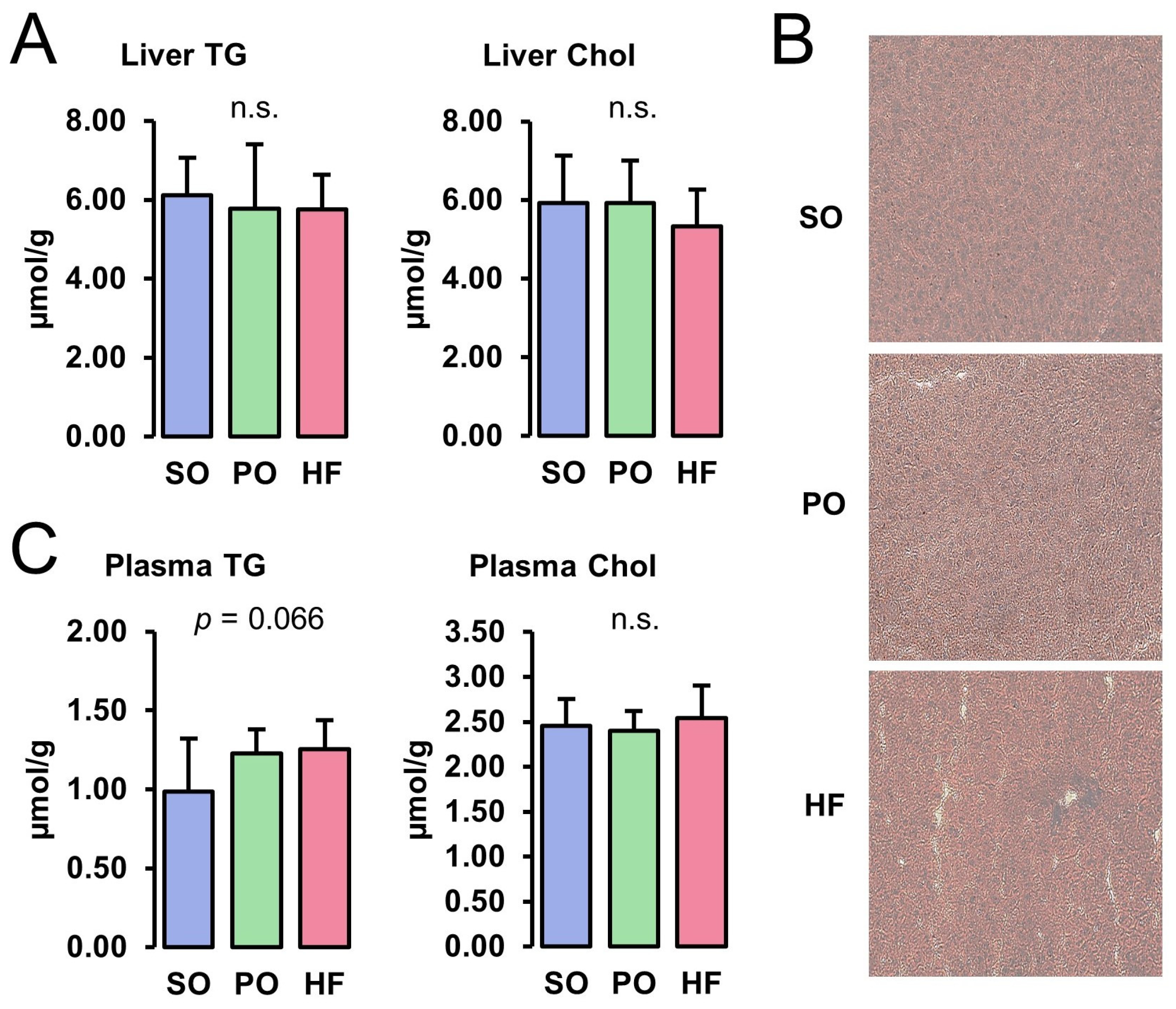
3.2. Lipid Concentrations in Liver and Plasma
3.3. Fatty Acid Concentrations in Liver Total Lipids
3.4. Fatty Acid Concentrations in Skeletal Muscle Total Lipids
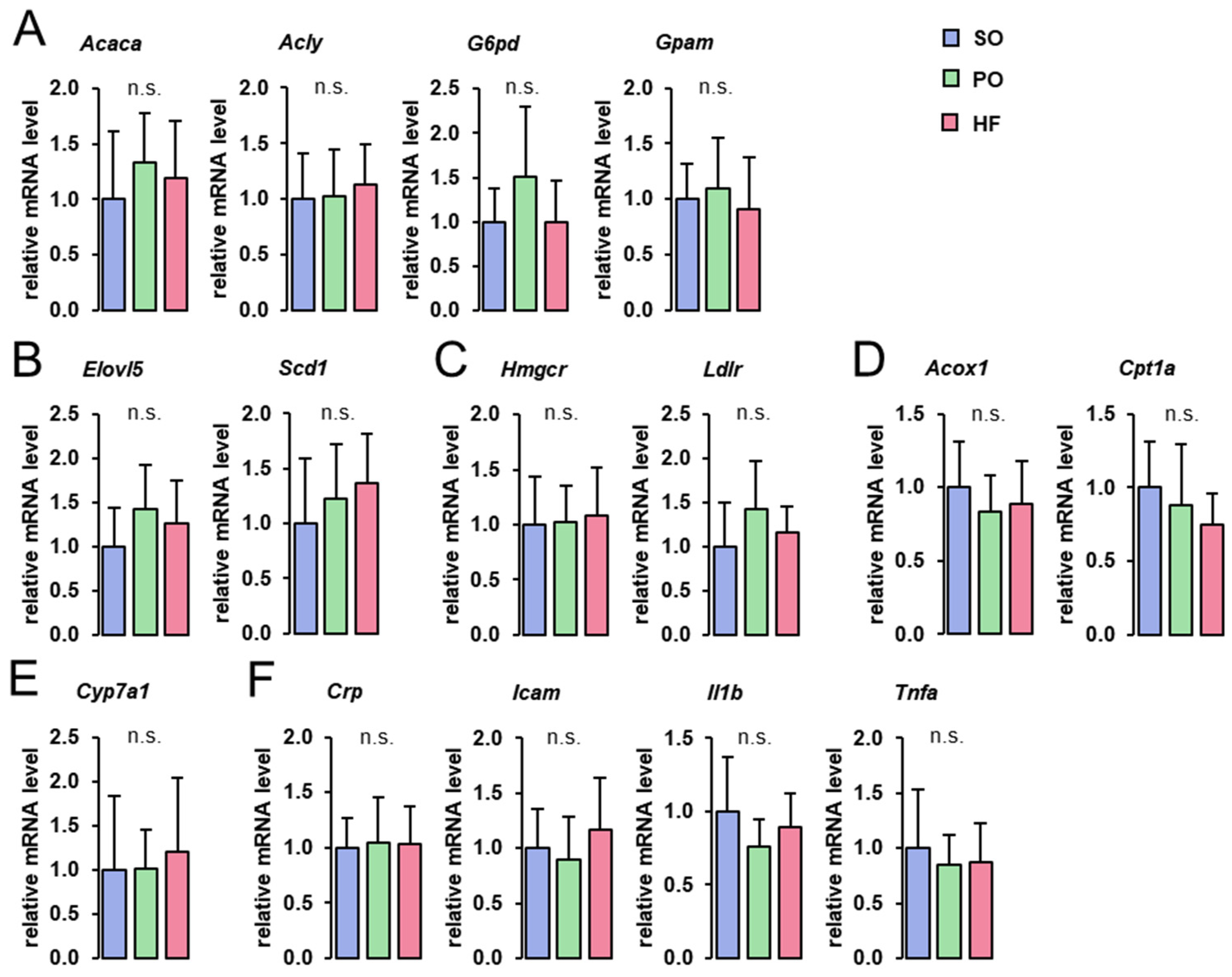
3.5. Hepatic Expression of Genes Involved in Lipid Metabolism and Inflammation
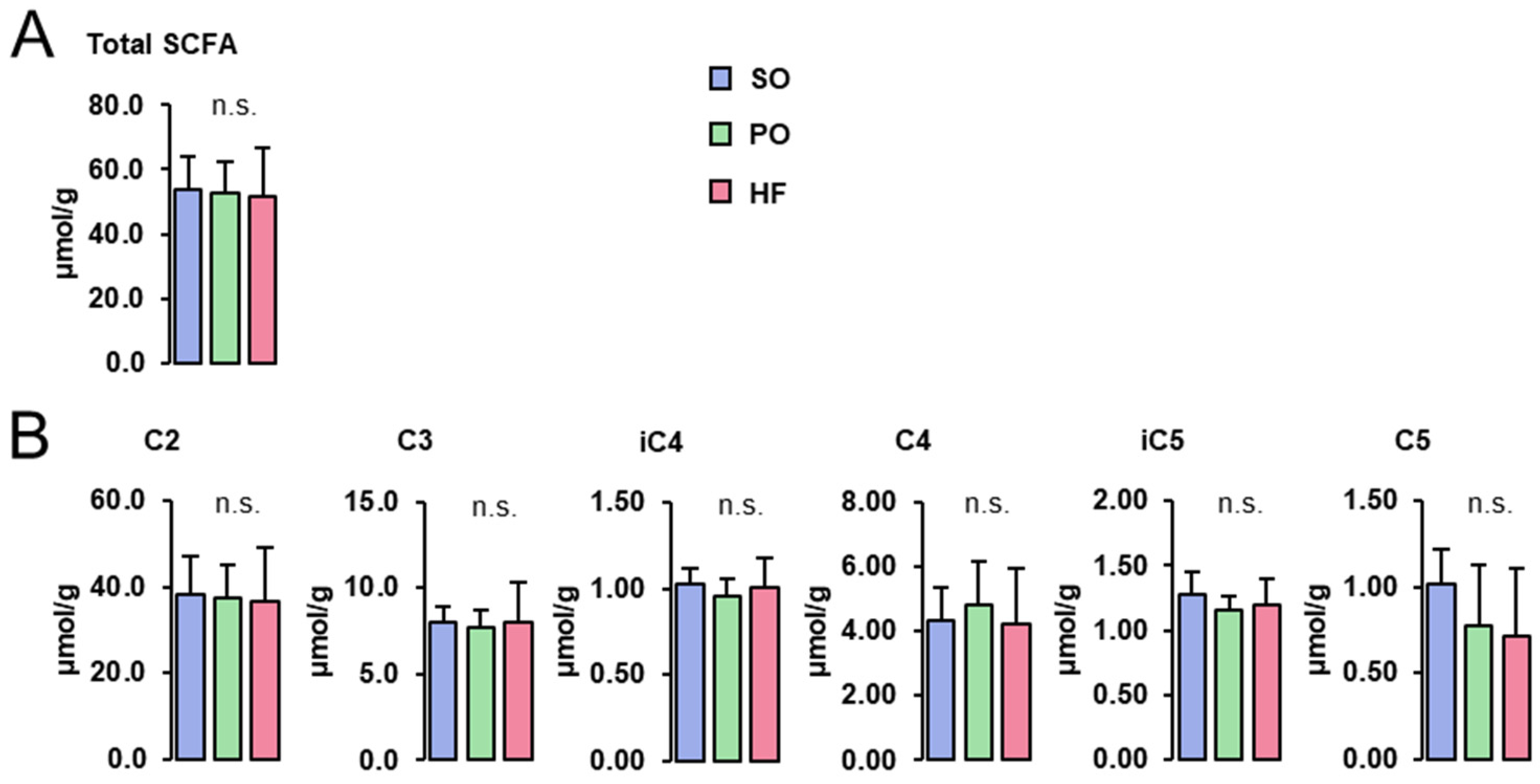
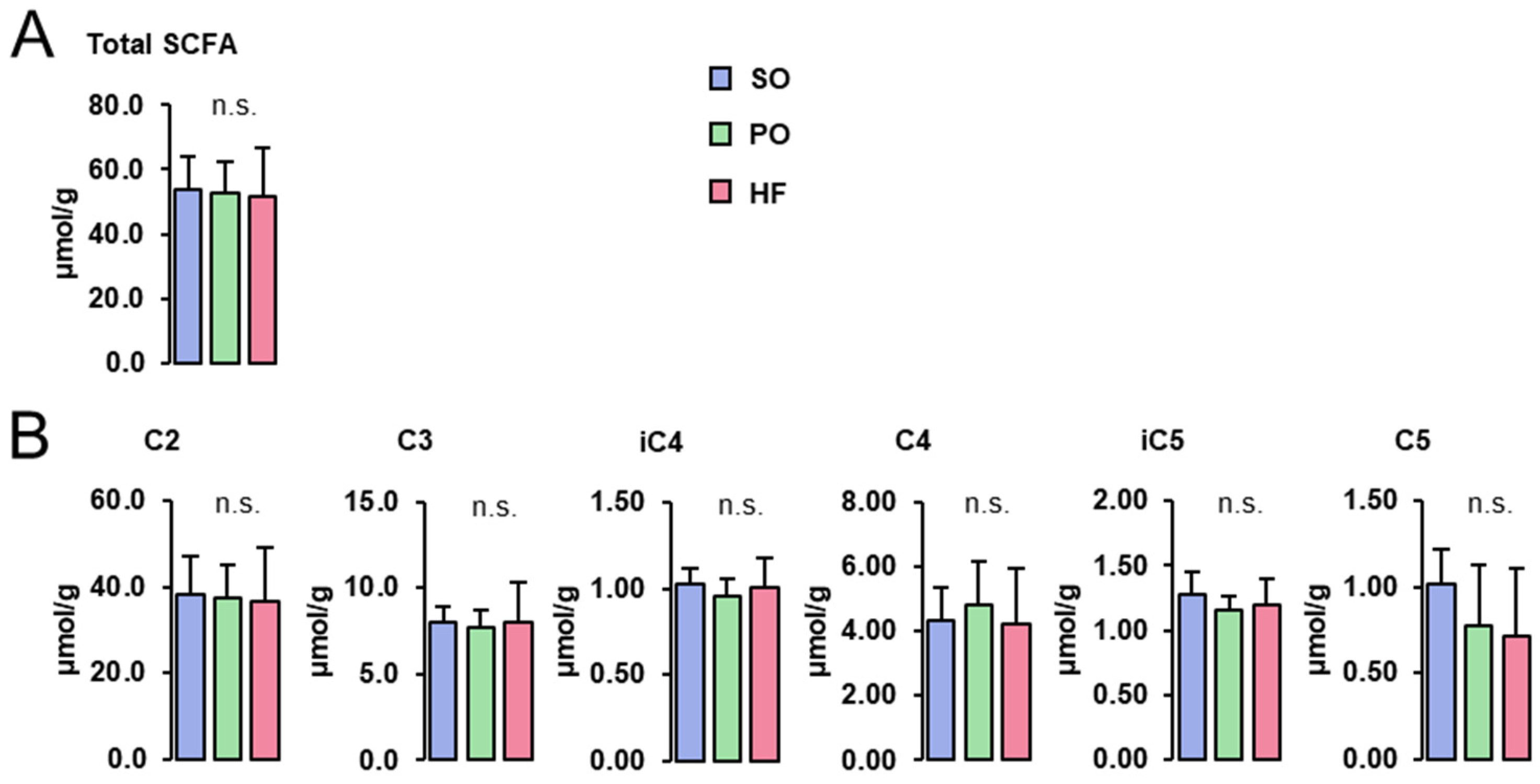
3.6. SCFA Concentrations in the Cecum Digesta
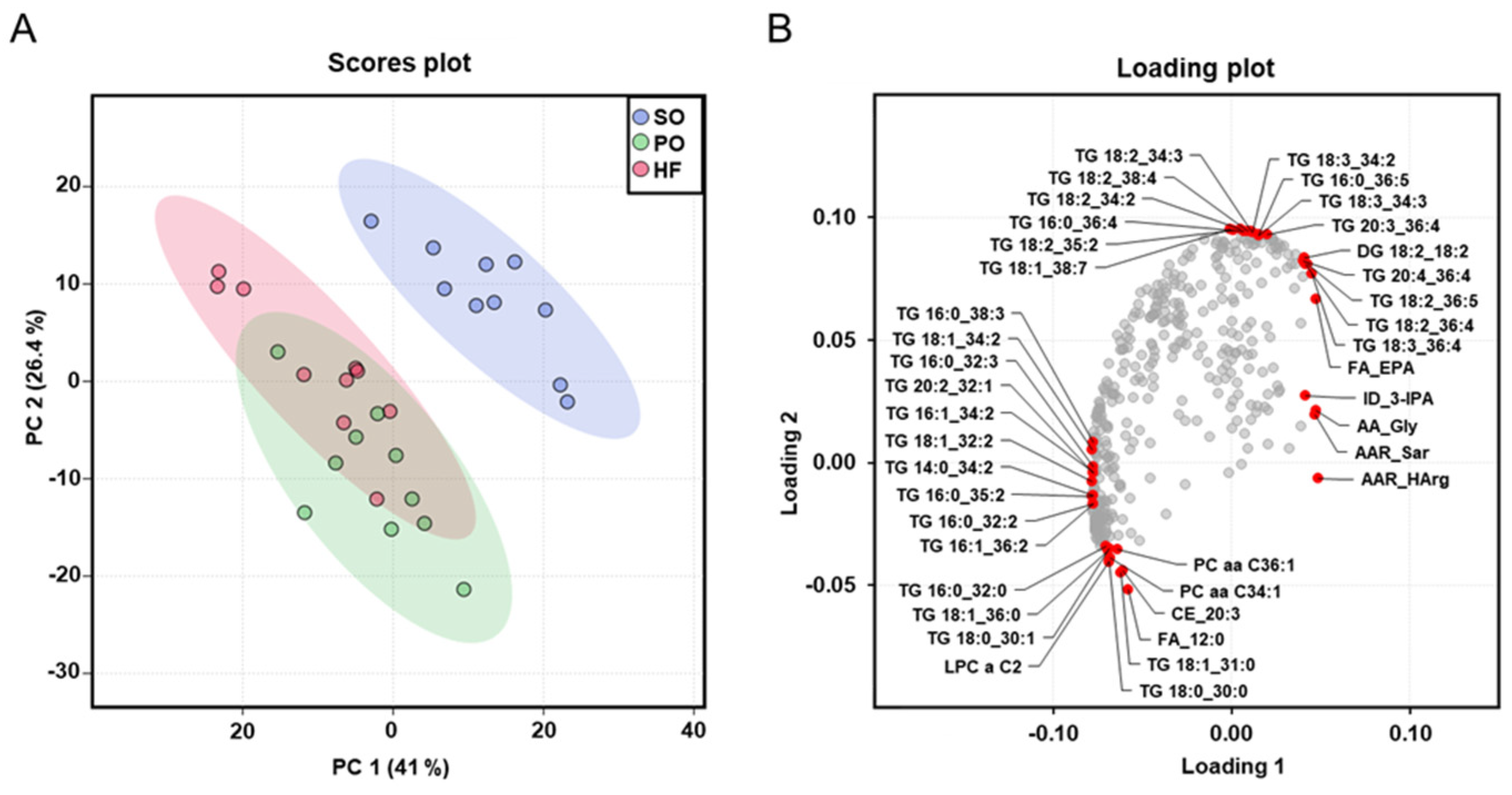
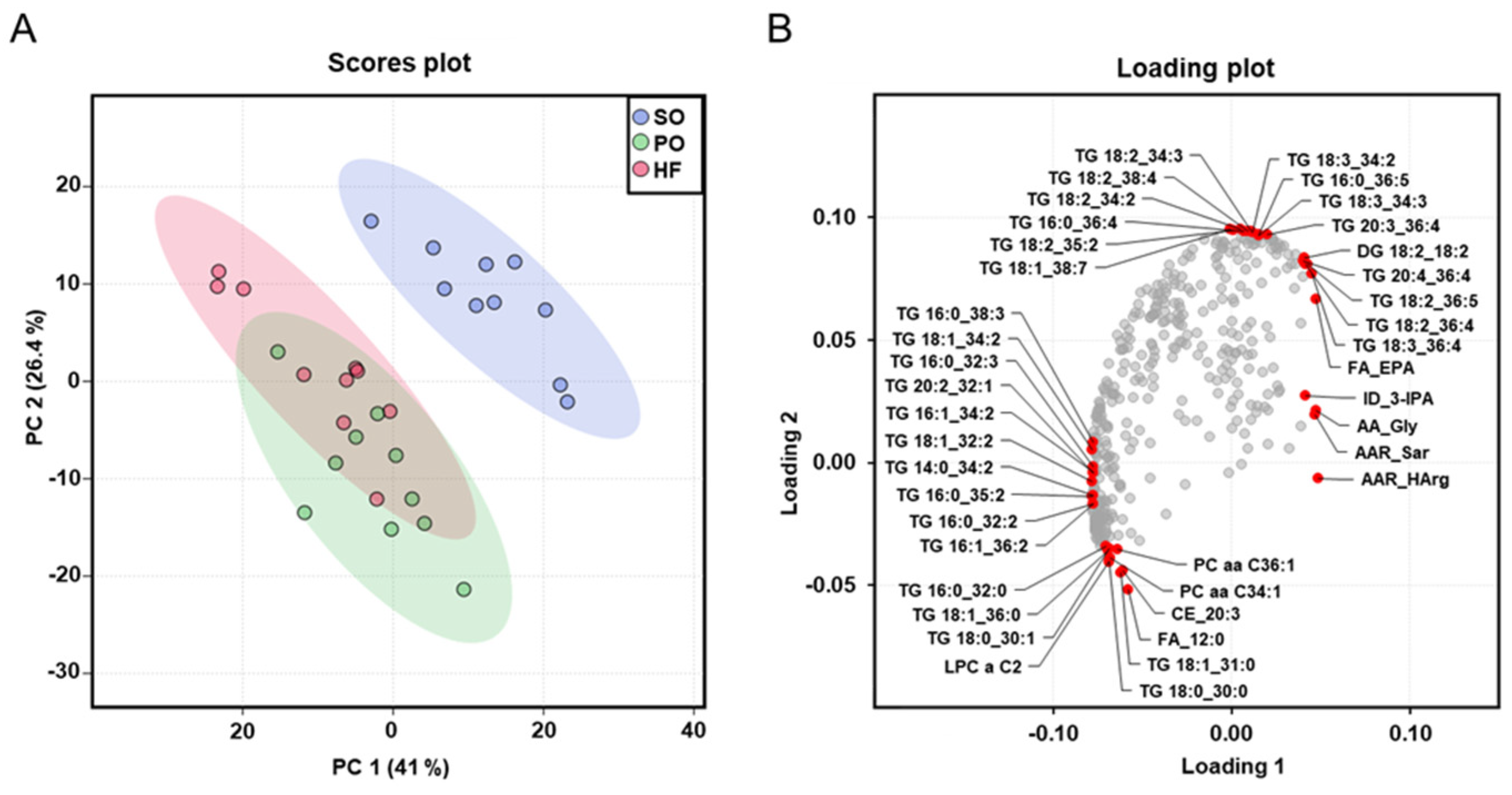
3.7. Identification of Altered Plasma Metabolites Using Targeted Metabolomics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mba, O.I.; Dumont, M.J.; Ngadi, M. Palm oil: Processing, characterization and utilization in the food industry–A review. Food Biosci. 2015, 10, 26–41. [Google Scholar] [CrossRef]
- Mancini, A.; Imperlini, E.; Nigro, E.; Montagnese, C.; Daniele, A.; Orrù, S.; Buono, P. Biological and nutritional properties of palm oil and palmitic acid: Effects on health. Molecules 2015, 20, 17339–17361. [Google Scholar] [CrossRef] [PubMed]
- Pinotti, L.; Giromini, C.; Ottoboni, M.; Tretola, M.; Marchis, D. Review: Insects and former foodstuffs for upgrading food waste biomasses/streams to feed ingredients for farm animals. Animal 2019, 13, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Pinotti, L.; Ottoboni, M. Substrate as insect feed for bio-mass production. J. Insects Food Feed. 2021, 7, 585–596. [Google Scholar] [CrossRef]
- van Huis, A.; Oonincx, D.G.A.B. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef]
- Oonincx, D.G.; de Boer, I.J. Environmental impact of the production of mealworms as a protein source for humans-a life cycle assessment. PLoS ONE 2012, 7, e51145. [Google Scholar] [CrossRef]
- Kim, Y.B.; Kim, D.H.; Jeong, S.B.; Lee, J.W.; Kim, T.H.; Lee, H.G.; Lee, K.W. Black soldier fly larvae oil as an alternative fat source in broiler nutrition. Poult. Sci. 2020, 99, 3133–3143. [Google Scholar] [CrossRef]
- Marschall, M.J.M.; Grundmann, S.M.; Gessner, D.K.; Wen, G.; Most, E.; Eder, K.; Ringseis, R. Feeding of Hermetia illucens larvae meal attenuates hepatic lipid synthesis and fatty liver development in obese Zucker rats. Nutrients. 2023, 15, 287. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Zähringer, J.; Beyerbach, J.; Werner, S.S.; Nagavci, B.; Heseker, H.; Koletzko, B.; Meerpohl, J.J.; International Union of Nutritional Sciences (IUNS) Task force on Dietary Fat Quality. A scoping review of current guidelines on dietary fat and fat quality. Ann. Nutr. Metab. 2021, 77, 65–82. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Health and Human Services; U.S. Department of Agriculture. The Dietary Guidelines for Americans, 2020–2025. Available online: https://www.dietaryguidelines.gov/ (accessed on 15 August 2023).
- Arnett, D.K.; Blumenthal, R.S.; Albert, M.A.; Buroker, A.B.; Goldberger, Z.D.; Hahn, E.J.; Himmelfarb, C.D.; Khera, A.; Lloyd-Jones, D.; McEvoy, J.W.; et al. 2019 ACC/AHA guideline on the primary prevention of cardiovascular disease: A report of the American College of Cardiology/American Heart Association Task Force on clinical practice guidelines. Circulation 2019, 140, e596–e646. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Laboratory Animals, 4th ed.; National Academy Press: Washington, DC, USA, 1995.
- Schäfer, L.; Grundmann, S.M.; Maheshwari, G.; Höring, M.; Liebisch, G.; Most, E.; Eder, K.; Ringseis, R. Effect of replacement of soybean oil by Hermetia illucens fat on performance, digestibility, cecal microbiome, liver transcriptome and liver and plasma lipidomes of broilers. J. Anim. Sci. Biotechnol. 2023, 14, 20. [Google Scholar] [CrossRef]
- Schlegel, G.; Ringseis, R.; Shibani, M.; Most, E.; Schuster, M.; Schwarz, F.J.; Eder, K. Influence of a rumen-protected conjugated linoleic acid mixture on carcass traits and meat quality in young Simmental heifers. J. Anim. Sci. 2012, 90, 1532–1540. [Google Scholar] [CrossRef]
- Krawielitzki, K.; Schadereit, R.; Borgmann, E.; Evers, B. Use of 51-Cr2O3 and TiO2 as markers for the determination of pas-sage rate and protein digestibility in rats. Arch. Tierernahr. 1987, 37, 1085–1099. [Google Scholar] [CrossRef]
- Brandt, M.; Allam, S.M. Analytik von TiO2 im Darminhalt und Kot nach Kjeldalaufschluß. Arch. Anim. Nutr. 1987, 37, 453–454. [Google Scholar]
- Zöllner, N.; Kirsch, K. Colorimetric Method for Determination of Total Lipids. JEM. 1962, 135, 545–550. [Google Scholar]
- Hara, A.; Radin, N.S. Lipid extraction of tissues with a low-toxicity solvent. Anal. Biochem. 1978, 90, 420–426. [Google Scholar] [CrossRef]
- Eder, K.; Kirchgessner, M. Dietary fat influences the effect of zinc deficiency on liver lipids and fatty acids in rats force-fed equal quantities of diet. J. Nutr. 1994, 124, 1917–1926. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RE-SEARCH0034. [Google Scholar] [CrossRef]
- Ringseis, R.; Grundmann, S.M.; Schuchardt, S.; Most, E.; Eder, K. Limited impact of pivalate-induced secondary carnitine deficiency on hepatic transcriptome and hepatic and plasma metabolome in nursery pigs. Metabolites 2021, 11, 573. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A web server for metabolomic data analysis and interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef]
- Seitz, H.K.; Moreira, B.; Neuman, M.G. Pathogenesis of alcoholic fatty liver. A narrative review. Life 2023, 13, 1662. [Google Scholar] [CrossRef] [PubMed]
- Ringseis, R.; Gessner, D.K.; Eder, K. The gut-liver axis in the control of energy metabolism and food intake in animals. Annu. Rev. Anim. Biosci. 2020, 8, 295–319. [Google Scholar] [CrossRef] [PubMed]
- Biasato, I.; Ferrocino, I.; Dabbou, S.; Evangelista, R.; Gai, F.; Gasco, L.; Cocolin, L.; Capucchio, M.T.; Schiavone, A. Black soldier fly and gut health in broiler chickens: Insights into the relationship between cecal microbiota and intestinal mucin composition. J. Anim. Sci. Biotechnol. 2020, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- Jian, S.; Zhang, L.; Ding, N.; Yang, K.; Xin, Z.; Hu, M.; Zhou, Z.; Zhao, Z.; Deng, B.; Deng, J. Effects of black soldier fly larvae as protein or fat sources on apparent nutrient digestibility, fecal microbiota, and metabolic profiles in beagle dogs. Front. Microbiol. 2022, 13, 1044986. [Google Scholar] [CrossRef]
- Schwingshackl, L.; Schlesinger, S. Coconut oil and cardiovascular disease risk. Curr. Atheroscler. Rep. 2023, 25, 231–236. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SO Diet | PO Diet | HF Diet | |
|---|---|---|---|
| Component, g/kg | |||
| Cornstarch | 525.6 | 525.6 | 525.6 |
| Casein | 200 | 200 | 200 |
| Sucrose | 100 | 100 | 100 |
| Mineral mix 1 | 35 | 35 | 35 |
| Vitamin mix 2 | 10 | 10 | 10 |
| Soybean oil | 70 | 0.7 | 0.7 |
| Palm oil | - | 68.6 | - |
| Hermetia illucens fat | - | - | 68.6 |
| Linseed oil | - | 0.7 | 0.7 |
| Cellulose | 50 | 50 | 50 |
| L-cysteine | 4.4 | 4.4 | 4.4 |
| TiO2 | 5 | 5 | 5 |
| Fatty acid composition of dietary total lipids, g/100 g total fatty acids | |||
| C10:0 | 0.01 | 0.03 | 1.1 |
| C12:0 | 0.39 | 0.66 | 54.8 |
| C14:0 | 0.22 | 1.19 | 8.78 |
| C16:0 | 10.6 | 39.9 | 11 |
| C16:1 n-7 | 0.11 | 0.21 | 2.64 |
| C18:0 | 4.81 | 4.5 | 1.49 |
| C18:1 n-9 | 23.1 | 39.9 | 7.65 |
| C18:2 n-6 | 51.7 | 10.4 | 9.6 |
| C18:3 n-6 | 0.2 | 0.49 | 0.29 |
| C18:3 n-3 | 5.66 | 0.91 | 1.35 |
| C20:0 | 1.21 | 0.59 | 0.36 |
| C20:1 n-9 | 0.25 | 0.15 | 0.04 |
| C20:4 n-6 | 0.26 | 0.43 | 0.14 |
| C22:0 | 0.71 | 0.36 | 0.36 |
| C22:1 n-9 | 0.57 | 0.1 | 0.23 |
| SFA/UFA ratio | 0.22 | 0.9 | 3.55 |
| n-6 PUFA/n-3 PUFA ratio | 9.22 | 12.44 | 7.43 |
| SO | PO | HF | p-Value | |
|---|---|---|---|---|
| Final body weight | 343 ± 34 | 351 ± 26 | 351 ± 24 | 0.769 |
| Daily body weight gain, g | 3.64 ± 0.83 | 3.93 ± 0.55 | 3.81 ± 0.44 | 0.589 |
| Daily feed intake, g | 20.6 ± 1.8 | 22.2 ± 1.5 | 21.8 ± 0.8 | 0.239 |
| Organ weights | ||||
| Liver, g | 12.7 ± 2.3 | 13.0 ± 1.4 | 14.0 ± 1.3 | 0.210 |
| Liver to body weight ratio, mg/g | 3.67 ± 0.37 | 3.70 ± 0.21 | 4.00 ± 0.44 | 0.080 |
| Kidney, g | 1.16 ± 0.14 | 1.15 ± 0.06 | 1.21 ± 0.11 | 0.457 |
| M. soleus, g | 0.12 ± 0.02 | 0.13 ± 0.01 | 0.13 ± 0.01 | 0.595 |
| M. gastrocnemius, g | 1.77 ± 0.23 | 1.85 ± 0.17 | 1.89 ± 0.14 | 0.392 |
| M. rectus femoris, g | 0.91 ± 0.14 | 1.01 ± 0.13 | 0.99 ± 0.13 | 0.199 |
| AID Total fat, % | 95.2 ± 3.1 a | 93.0 ± 2.4 ab | 89.3 ± 4.1 b | 0.004 |
| SO | PO | HF | p-Value | |
|---|---|---|---|---|
| Fatty acid, µmol/g | ||||
| C12:0 | 0.64 ± 0.17 b | 0.64 ± 0.15 b | 0.91 ± 0.18 a | 0.001 |
| C14:0 | 0.91 ± 0.32 b | 1.11 ± 0.18 b | 1.62 ± 0.24 a | <0.001 |
| C14:1 n-5 | 0.42 ± 0.21 | 0.37 ± 0.07 | n.d. | 0.880 |
| C16:0 | 27.4 ± 3.3 | 28.1 ± 2.4 | 29.7 ± 3.5 | 0.260 |
| C16:1 n-7 | 2.20 ± 0.95 c | 3.23 ± 0.37 b | 4.86 ± 1.48 a | 0.001 |
| C17:0 | 0.91 ± 0.20 a | 0.70 ± 0.22 b | 0.72 ± 0.11 ab | 0.027 |
| C18:0 | 19.0 ± 2.3 | 18.0 ± 2.0 | 17.3 ± 1.9 | 0.195 |
| C18:1 n-9 | 14.6 ± 2.0 b | 20.1 ± 1.9 a | 19.8 ± 4.7 a | <0.001 |
| C18:2 n-6 | 21.0 ± 3.3 a | 9.06 ± 1.19 c | 10.5 ± 0.9 b | <0.001 |
| C18:3 n-3 | n.d. b | 0.82 ± 0.17 a | 1.00 ± 0.13 a | 0.022 |
| C18:3 n-6 | 0.66 ± 0.15 a | 0.26 ± 0.05 c | 0.36 ± 0.06 b | <0.001 |
| C20:2 n-6 | 0.65 ± 0.23 a | 0.37 ± 0.09 b | 0.22 ± 0.07 c | <0.001 |
| C20:3 n-6 | 1.03 ± 0.76 | 1.13 ± 0.26 | 1.17 ± 0.20 | 0.825 |
| C20:4 n-6 | 18.5 ± 4.9 | 15.8 ± 3.9 | 15.9 ± 3.1 | 0.255 |
| C22:5 n-3 | 0.54 ± 0.20 a | 0.22 ± 0.08 b | 0.29 ± 0.08 b | <0.001 |
| C22:6 n-3 | 3.44 ± 1.30 | 2.76 ± 1.30 | 2.98 ± 1.22 | 0.483 |
| Total | 112 ± 11 | 103 ± 11 | 107 ± 12 | 0.200 |
| SO | PO | HF | p-Value | |
|---|---|---|---|---|
| Fatty acid, µmol/g | ||||
| C12:0 | 0.48 ± 0.16 b | 2.34 ± 1.53 a | 1.68 ± 1.52 a | <0.001 |
| C14:0 | 0.65 ± 0.32 | 1.12 ± 0.65 | 1.07 ± 0.83 | 0.247 |
| C14:1 n-5 | 0.19 ± 0.13 | 0.22 ± 0.03 | 0.22 ± 0.16 | 0.249 |
| C16:0 | 13.9 ± 4.7 | 13.4 ± 3.9 | 14.8 ± 7.4 | 0.855 |
| C16:1 n-7 | 1.28 ± 0.65 | 1.39 ± 0.82 | 1.75 ± 1.52 | 0.756 |
| C17:0 | 0.23 ± 0.11 | 0.15 ± 0.05 | 0.20 ± 0.05 | 0.139 |
| C18:0 | 6.01 ± 1.61 | 4.81 ± 0.80 | 5.37 ± 0.91 | 0.145 |
| C18:1 n-9 | 9.97 ± 5.53 | 8.42 ± 3.80 | 10.7 ± 7.2 | 0.690 |
| C18:2 n-6 | 16.7 ± 8.3 a | 8.58 ± 2.58 b | 9.23 ± 4.01 b | 0.016 |
| C18:3 n-3 | 0.83 ± 0.54 a | 0.23 ± 0.15 b | 0.33 ± 0.27 b | 0.008 |
| C20:4 n-6 | 4.43 ± 1.02 | 3.46 ± 0.54 | 4.39 ± 1.08 | 0.056 |
| C22:5 n-3 | 0.55 ± 0.13 a | 0.35 ± 0.07 b | 0.40 ± 0.14 ab | 0.004 |
| C22:6 n-3 | 1.76 ± 0.72 a | 1.05 ± 0.34 b | 1.56 ± 0.47 ab | 0.028 |
| Total | 56.9 ± 23.2 | 54.4 ± 14.6 | 51.6 ± 25.1 | 0.530 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ringseis, R.; Marschall, M.J.M.; Grundmann, S.M.; Schuchardt, S.; Most, E.; Gessner, D.K.; Wen, G.; Eder, K. Effect of Hermetia illucens Fat, Compared with That of Soybean Oil and Palm Oil, on Hepatic Lipid Metabolism and Plasma Metabolome in Healthy Rats. Animals 2023, 13, 3356. https://doi.org/10.3390/ani13213356
Ringseis R, Marschall MJM, Grundmann SM, Schuchardt S, Most E, Gessner DK, Wen G, Eder K. Effect of Hermetia illucens Fat, Compared with That of Soybean Oil and Palm Oil, on Hepatic Lipid Metabolism and Plasma Metabolome in Healthy Rats. Animals. 2023; 13(21):3356. https://doi.org/10.3390/ani13213356
Chicago/Turabian StyleRingseis, Robert, Magdalena J. M. Marschall, Sarah M. Grundmann, Sven Schuchardt, Erika Most, Denise K. Gessner, Gaiping Wen, and Klaus Eder. 2023. "Effect of Hermetia illucens Fat, Compared with That of Soybean Oil and Palm Oil, on Hepatic Lipid Metabolism and Plasma Metabolome in Healthy Rats" Animals 13, no. 21: 3356. https://doi.org/10.3390/ani13213356
APA StyleRingseis, R., Marschall, M. J. M., Grundmann, S. M., Schuchardt, S., Most, E., Gessner, D. K., Wen, G., & Eder, K. (2023). Effect of Hermetia illucens Fat, Compared with That of Soybean Oil and Palm Oil, on Hepatic Lipid Metabolism and Plasma Metabolome in Healthy Rats. Animals, 13(21), 3356. https://doi.org/10.3390/ani13213356