
The Protective Effects of Astaxanthin (AST) in the Liver of Weaned Piglets



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animals
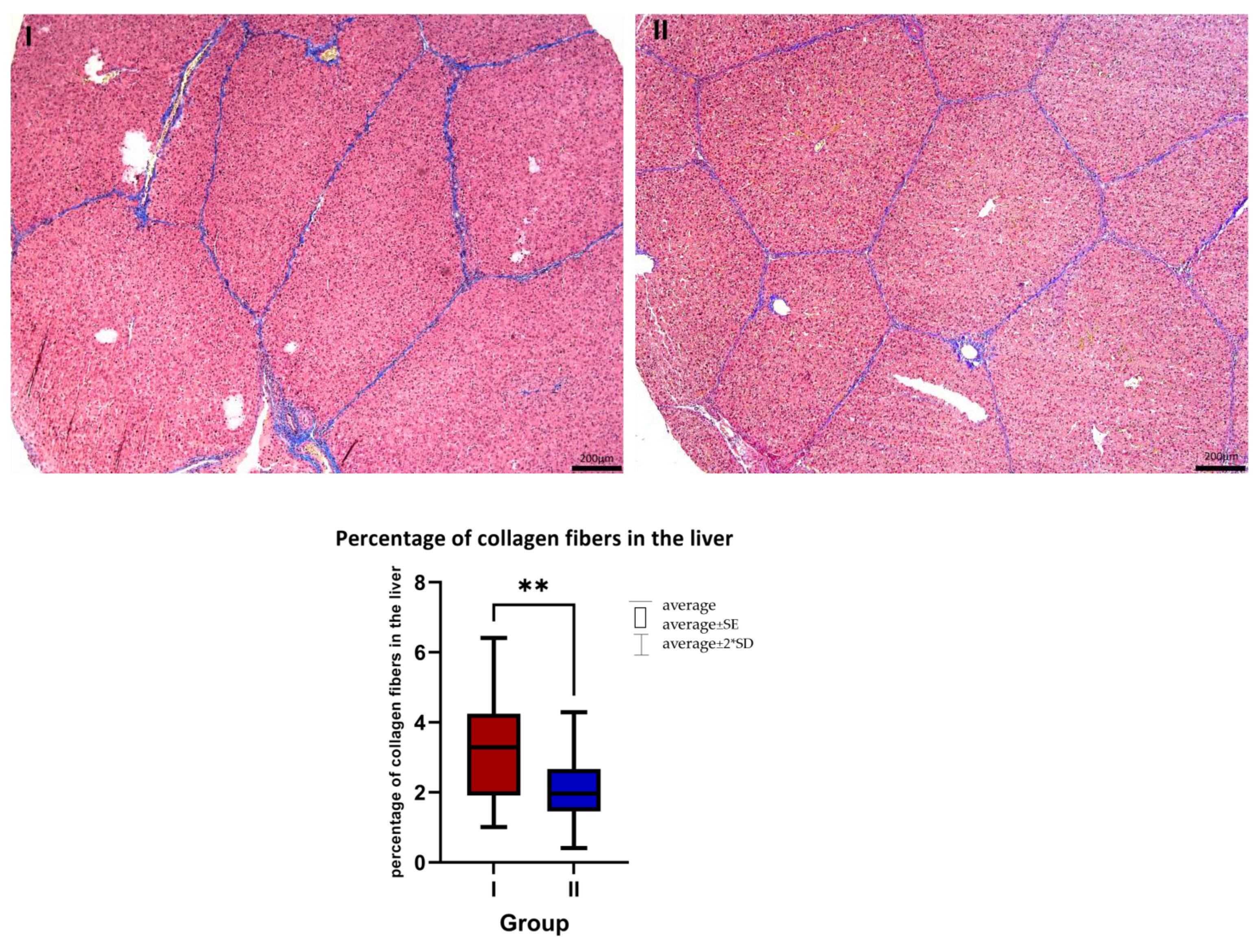
2.3. Histology
2.4. RNA Isolation and Quantitative PCR (qPCR)
3. Results
3.1. Histology
3.2. Gene Expression
4. Discussion
4.1. Histology
4.2. Gene Expression
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, J.-T.; Kotani, K. Astaxanthin as a Potential Protector of Liver Function: A Review. J. Clin. Med. Res. 2016, 8, 701–704. [Google Scholar] [CrossRef]
- Liu, H.; Liu, M.; Fu, X.; Zhang, Z.; Zhu, L.; Zheng, X.; Liu, J. Astaxanthin Prevents Alcoholic Fatty Liver Disease by Modulating Mouse Gut Microbiota. Nutrients 2018, 10, 1298. [Google Scholar] [CrossRef]
- Xuan, R.R.; Niu, T.T.; Chen, H.M. Astaxanthin Blocks Preeclampsia Progression by Suppressing Oxidative Stress and Inflammation. Mol. Med. Rep. 2016, 14, 2697–2704. [Google Scholar] [CrossRef]
- Goto, S.; Kogure, K.; Abe, K.; Kimata, Y.; Kitahama, K.; Yamashita, E.; Terada, H. Efficient Radical Trapping at the Surface and inside the Phospholipid Membrane Is Responsible for Highly Potent Antiperoxidative Activity of the Carotenoid Astaxanthin. Biochim. Biophys. Acta Biomembr. 2001, 1512, 251–258. [Google Scholar] [CrossRef] [PubMed]
- McNulty, H.P.; Byun, J.; Lockwood, S.F.; Jacob, R.F.; Mason, R.P. Differential Effects of Carotenoids on Lipid Peroxidation Due to Membrane Interactions: X-Ray Diffraction Analysis. Biochim. Biophys. Acta Biomembr. 2007, 1768, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free Radicals, Antioxidants in Disease and Health. Int. J. Biomed. Sci. 2008, 4, 89. [Google Scholar]
- Sanchez-Valle, V.; Chavez-Tapia, N.C.; Uribe, M.; Mendez-Sanchez, N. Role of Oxidative Stress and Molecular Changes in Liver Fibrosis: A Review. Curr. Med. Chem. 2012, 19, 4850–4860. [Google Scholar] [CrossRef] [PubMed]
- Kolios, G.; Valatas, V.; Kouroumalis, E. Role of Kupffer Cells in the Pathogenesis of Liver Disease. World J. Gastroenterol. 2006, 12, 7413–7420. [Google Scholar] [CrossRef]
- Friedman, S.L. Molecular Regulation of Hepatic Fibrosis, an Integrated Cellular Response to Tissue Injury. J. Biol. Chem. 2000, 275, 2247–2250. [Google Scholar] [CrossRef]
- Jaeschke, H. Reactive Oxygen and Mechanisms of Inflammatory Liver Injury: Present Concepts. J. Gastroenterol. Hepatol. 2011, 26, 173–179. [Google Scholar] [CrossRef]
- Lieber, C.S. Alcoholic Liver Disease: New Insights in Pathogenesis Lead to New Treatments. J. Hepatol. 2000, 32, 113–128. [Google Scholar] [CrossRef] [PubMed]
- Ciaraldi, T.P.; Boeder, S.C.; Mudaliar, S.R.; Giovannetti, E.R.; Henry, R.R.; Pettus, J.H. Astaxanthin, a Natural Antioxidant, Lowers Cholesterol and Markers of Cardiovascular Risk in Individuals with Prediabetes and Dyslipidaemia. Diabetes Obes. Metab. 2023, 25, 1985–1994. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.D.; Youn, Y.K.; Shin, W.G. Positive Effects of Astaxanthin on Lipid Profiles and Oxidative Stress in Overweight Subjects. Plant Foods Hum. Nutr. 2011, 66, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Bae, M.; Kim, B.; Park, Y.K.; Koo, S.I.; Lee, J.Y. Astaxanthin Prevents and Reverses the Activation of Mouse Primary Hepatic Stellate Cells. J. Nutr. Biochem. 2016, 29, 21–26. [Google Scholar] [CrossRef]
- Sila, A.; Kamoun, Z.; Ghlissi, Z.; Makni, M.; Nasri, M.; Sahnoun, Z.; Nedjar-Arroume, N.; Bougatef, A. Ability of Natural Astaxanthin from Shrimp By-Products to Attenuate Liver Oxidative Stress in Diabetic Rats. Pharmacol. Rep. 2015, 67, 310–316. [Google Scholar] [CrossRef]
- Ni, Y.; Nagashimada, M.; Zhuge, F.; Zhan, L.; Nagata, N.; Tsutsui, A.; Nakanuma, Y.; Kaneko, S.; Ota, T. Astaxanthin Prevents and Reverses Diet-Induced Insulin Resistance and Steatohepatitis in Mice: A Comparison with Vitamin E. Sci. Rep. 2015, 5, 17192. [Google Scholar] [CrossRef]
- Zhang, Z.; Guo, C.; Jiang, H.; Han, B.; Wang, X.; Li, S.; Lv, Y.; Lv, Z.; Zhu, Y. Inflammation Response after the Cessation of Chronic Arsenic Exposure and Post-Treatment of Natural Astaxanthin in Liver: Potential Role of Cytokine-Mediated Cell-Cell Interactions. Food Funct. 2020, 11, 9252–9262. [Google Scholar] [CrossRef]
- Jia, Y.; Wu, C.; Kim, J.; Kim, B.; Lee, S.-J. Astaxanthin Reduces Hepatic Lipid Accumulations in High-Fat-Fed C57BL/6J Mice via Activation of Peroxisome Proliferator-Activated Receptor (PPAR) Alpha and Inhibition of PPAR Gamma and Akt. J. Nutr. Biochem. 2016, 28, 9–18. [Google Scholar] [CrossRef]
- van Beers-Schreurs, H.M.; Vellenga, L.; Wensing, T.; Breukink, H.J. The Pathogenesis of the Post-Weaning Syndrome in Weaned Piglets: A Review. Vet. Q. 1992, 14, 29–34. [Google Scholar] [CrossRef]
- Seki, E.; Schnabl, B. Role of Innate Immunity and the Microbiota in Liver Fibrosis: Crosstalk between the Liver and Gut. J. Physiol. 2012, 590, 447–458. [Google Scholar] [CrossRef]
- Bailey, S.M.; Cunningham, C.C. Contribution of Mitochondria to Oxidative Stress Associated with Alcoholic Liver Disease. Free Radic. Biol. Med. 2002, 32, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Thayer, W.S.; Rubin, E. Antimycin Inhibition as a Probe of Mitochondrial Function in Isolated Rat Hepatocytes Effects of Chronic Ethanol Consumption. BBA Mol. Cell Res. 1982, 721, 328–335. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Zhu, W.; Guo, Q.; Luo, W.; Zhang, J.; Xu, W.; Xu, J. Weaning Induced Hepatic Oxidative Stress, Apoptosis, and Aminotransferases through MAPK Signaling Pathways in Piglets. Oxid. Med. Cell Longev. 2016, 2016, 4768541. [Google Scholar] [CrossRef] [PubMed]
- Szczepanik, K.; Furgał-Dierżuk, I.; Gala, Ł.; Świątkiewicz, M. Effects of Hermetia Illucens Larvae Meal and Astaxanthin as Feed Additives on Health and Production Indices in Weaned Pigs. Animals 2023, 13, 163. [Google Scholar] [CrossRef]
- Bergstrom, J.R.; Nelssen, J.L.; Tokach, M.D.; DeRouchey, J.M.; Goodband, R.D.; Dritz, S.S. An Evaluation of Astaxanthin as a Nutraceutical Growth Promoter in Starter Diets for Weanling Pigs. Kans. Agric. Exp. Stn. Res. Rep. 2007, 10, 83–87. [Google Scholar] [CrossRef]
- Miki, W. Biological Functions and Activities of Animal Carotenoids. Pure Appl. Chem. 1991, 63, 141–146. [Google Scholar] [CrossRef]
- Igielska-Kalwat, J.; Gościańska, J.; Nowak, I. Carotenoids as natural antioxidants. Postepy Hig. Med. Dosw. 2015, 69, 418–428. [Google Scholar] [CrossRef]
- Grela, E.R.; Skomiał, J. Zalecenia Żywieniowe i Wartość Pokarmowa Pasz Dla Świń. Normy Żywienia Świń.; Wyd. Instytut Fizjologii i Żywienia Zwierząt PAN: Jabłonna, Poland, 2020. [Google Scholar]
- Horwitz, W. Official Methods of Analyses, 17th ed.; AOAC: Gaithersburg, MD, USA, 2009. [Google Scholar]
- Hoffmann, L.; Schiemann, R. Von Der Kalorie Zum Joule: Neue Größenbeziehungen Bei Messungen Des Energieumsatzes Und Bei Der Berechnung von Kennzahlen Der Energetischen Futterbewertung. Archiv. Tierernaehrung 1980, 30, 733–742. [Google Scholar] [CrossRef]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The Biological Stress of Early Weaned Piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef]
- Pluske, J.R.; Kerton, D.J.; Cranwell, P.D.; Campbell, R.G.; Mullan, B.P.; King, R.H.; Power, G.N.; Pierzynowski, S.G.; Westrom, B.; Rippe, C.; et al. Age, Sex, and Weight at Weaning Influence Organ Weight and Gastrointestinal Development of Weanling Pigs. Aust. J. Agric. Res. 2003, 54, 515–527. [Google Scholar] [CrossRef]
- Kisseleva, T.; Brenner, D.A. Role of Hepatic Stellate Cells in Fibrogenesis and the Reversal of Fibrosis. J. Gastroen. Hepatol. 2007, 22, S73–S78. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.X.; Török, N.J. Liver Injury and the Activation of the Hepatic Myofibroblasts. Curr. Pathobiol. Rep. 2013, 1, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Chen, K.; Lu, J.; Cheng, P.; Xu, L.; Dai, W.; Wang, F.; He, L.; Zhang, Y.; Chengfen, W.; et al. Protective Effect of Astaxanthin on Liver Fibrosis through Modulation of TGF- β 1 Expression and Autophagy. Mediat. Inflamm. 2014, 2014, 954502. [Google Scholar] [CrossRef] [PubMed]
- Wynn, T.A. Common and Unique Mechanisms Regulate Fibrosis in Various Fibroproliferative Diseases. J. Clin. Investig. 2007, 117, 524–529. [Google Scholar] [CrossRef]
- Friedman, S.L. Evolving Challenges in Hepatic Fibrosis. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 425–436. [Google Scholar] [CrossRef]
- Uchiyama, A. Clinical Efficacy of Astaxanthin-Containing Haematococcus Pluvialis Extract for the Volunteers at Risk of Metabolic Syndrome. J. Clin. Biochem. Nutr. 2008, 43, 38–43. [Google Scholar]
- Enjoji, M.; Kohjima, M.; Nakamuta, M. Lipid Metabolism and the Liver. In The Liver in Systemic Diseases; Springer: Tokyo, Japan, 2016. [Google Scholar]
- Nakamuta, M.; Fujino, T.; Yada, R.; Yada, M.; Yasutake, K.; Yoshimoto, T.; Harada, N.; Higuchi, N.; Kato, M.; Kohjima, M.; et al. Impact of Cholesterol Metabolism and the LXRα-SREBP-1c Pathway on Nonalcoholic Fatty Liver Disease. Int. J. Mol. Med. 2009, 23, 603–608. [Google Scholar] [CrossRef][Green Version]
- Wang, M.; Xu, W.; Yu, J.; Liu, Y.; Ma, H.; Ji, C.; Zhang, C.; Xue, J.; Li, R.; Cui, H. Astaxanthin From Haematococcus Pluvialis Prevents High-Fat Diet-Induced Hepatic Steatosis and Oxidative Stress in Mice by Gut-Liver Axis Modulating Properties. Front. Nutr. 2022, 9, 840648. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, J.; Su, W.; Wu, J.; Wang, C.; Kong, X.; Gustafsson, J.Å.; Ding, J.; Ma, X.; Guan, Y. Liver X Receptor Activation Increases Hepatic Fatty Acid Desaturation by the Induction of SCD1 Expression through an LXRα-SREBP1c-Dependent Mechanism. J. Diabetes 2014, 6, 212–220. [Google Scholar] [CrossRef]
- Calkin, A.C.; Tontonoz, P. Liver X Receptor Signaling Pathways and Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1513–1518. [Google Scholar] [CrossRef]
- Jakobsson, T.; Treuter, E.; Gustafsson, J.Å.; Steffensen, K.R. Liver X Receptor Biology and Pharmacology: New Pathways, Challenges and Opportunities. Trends Pharmacol. Sci. 2012, 33, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Shang, P.; Qiangba, Y.; Xu, A.; Wang, Z.; Zhang, H. The Association of NR1H3 Gene with Lipid Deposition in the Pig. Lipids Health Dis. 2016, 15, 99. [Google Scholar] [CrossRef] [PubMed]
- Zelcer, N.; Tontonoz, P. Liver X Receptors as Integrators of Metabolic and Inflammatory Signaling. J. Clin. Investig. 2006, 116, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Beaven, S.W.; Wroblewski, K.; Wang, J.; Hong, C.; Bensinger, S.; Tsukamoto, H.; Tontonoz, P. Liver X Receptor Signaling Is a Determinant of Stellate Cell Activation and Susceptibility to Fibrotic Liver Disease. Gastroenterology 2011, 140, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Jiang, Q.; Xu, K. CREB Family: A Significant Role in Liver Fibrosis. Biochimie 2019, 163, 94–100. [Google Scholar] [CrossRef]
- Eng, F.J.; Friedman, S.L. Transcriptional Regulation in Hepatic Stellate Cells. Semin. Liver Dis. 2001, 21, 385–396. [Google Scholar] [CrossRef]
- Li, R.; Xin, T.; Li, D.; Wang, C.; Zhu, H.; Zhou, H. Therapeutic Effect of Sirtuin 3 on Ameliorating Nonalcoholic Fatty Liver Disease: The Role of the ERK-CREB Pathway and Bnip3-Mediated Mitophagy. Redox Biol. 2018, 18, 229–243. [Google Scholar] [CrossRef]
- Wang, P.; Deng, L.; Zhuang, C.; Cheng, C.; Xu, K. P-CREB-1 Promotes Hepatic Fibrosis through the Transactivation of Transforming Growth Factor-1 Expression in Rats. Int. J. Mol. Med. 2016, 38, 521–528. [Google Scholar] [CrossRef]
- Ferrando, A.A. The Role of NOTCH1 Signaling in T-ALL. Hematol./Educ. Program Am. Soc. Hematol. Am. Soc. Hematol. Educ. Program 2009, 2009, 353–361. [Google Scholar] [CrossRef]
- Takeshita, K.; Satoh, M.; Ii, M.; Silver, M.; Limbourg, F.P.; Mukai, Y.; Rikitake, Y.; Radtke, F.; Gridley, T.; Losordo, D.W.; et al. Critical Role of Endothelial Notch1 Signaling in Postnatal Angiogenesis. Circ. Res. 2007, 100, 70–78. [Google Scholar] [CrossRef]
- Yamaguchi, K.; Hayashi, M.; Uchida, Y.; Cheng, X.W.; Nakayama, T.; Matsushita, T.; Murohara, T.; Takeshita, K. Notch1 Haploinsufficiency in Mice Accelerates Adipogenesis. Sci. Rep. 2021, 11, 16761. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tang, Y.; Cai, D. IKKβ/NF-ΚB Disrupts Adult Hypothalamic Neural Stem Cells to Mediate a Neurodegenerative Mechanism of Dietary Obesity and Pre-Diabetes. Nat. Cell Biol. 2012, 14, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Du, G.; Xu, Y.; Li, X.; Fan, W.; Chen, J.; Liu, C.; Chen, G.; Liu, C.; Zern, M.A.; et al. Inhibition of Notch Signaling Pathway Prevents Cholestatic Liver Fibrosis by Decreasing the Differentiation of Hepatic Progenitor Cells into Cholangiocytes. Lab. Investig. 2016, 96, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Mensenkamp, A.R.; Jong, M.C.; Van Goor, H.; Van Luyn, M.J.A.; Bloks, V.; Havinga, R.; Voshol, P.J.; Hofker, M.H.; Van Dijk, K.W.; Havekes, L.M.; et al. Apolipoprotein E Participates in the Regulation of Very Low Density Lipoprotein-Triglyceride Secretion by the Liver. J. Biol. Chem. 1999, 274, 35711–35718. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.J.; Lin, C.H.; Ren, E.D.; Su, Y.; Zhu, W.Y. Effects of Early Intervention with Maternal Fecal Bacteria and Antibiotics on Liver Metabolome and Transcription in Neonatal Pigs. Front. Physiol. 2019, 10, 171. [Google Scholar] [CrossRef]
- Sodhi, S.S.; Ghosh, M.; Song, K.D.; Sharma, N.; Kim, J.H.; Kim, N.E.; Lee, S.J.; Kang, C.W.; Oh, S.J.; Jeong, D.K. An Approach to Identify SNPs in the Gene Encoding Acetyl-CoA Acetyltransferase-2 (ACAT-2) and Their Proposed Role in Metabolic Processes in Pig. PLoS ONE 2014, 9, e102432. [Google Scholar] [CrossRef]
- Fujii-Kuriyama, Y.; Imataka, H.; Sogawa, K.; Yasumoto, K.; Kikuchi, Y. Regulation of CYP1A1 Expression. FASEB J. 1992, 6, 706–710. [Google Scholar] [CrossRef]
- Hirschey, M.D.; Shimazu, T.; Goetzman, E.; Jing, E.; Schwer, B.; Lombard, D.B.; Grueter, C.A.; Harris, C.; Biddinger, S.; Ilkayeva, O.R.; et al. SIRT3 Regulates Mitochondrial Fatty-Acid Oxidation by Reversible Enzyme Deacetylation. Nature 2010, 464, 121–125. [Google Scholar] [CrossRef]
- Li, Y.; Tian, H.; Luo, H.; Fu, J.; Jiao, Y.; Li, Y. Prognostic Significance and Related Mechanisms of Hexokinase 1 in Ovarian Cancer. Onco Targets Ther. 2020, 13, 11583–11594. [Google Scholar] [CrossRef]
- Yang, Y.; Seo, J.M.; Nguyen, A.; Pham, T.X.; Park, H.J.; Park, Y.; Kim, B.; Bruno, R.S.; Lee, J. Astaxanthin-Rich Extract from the Green Alga Haematococcus Pluvialis Lowers Plasma Lipid Concentrations and Enhances Antioxidant Defense in Apolipoprotein E Knockout Mice. J. Nutr. 2011, 141, 1611–1617. [Google Scholar] [CrossRef]
- Kalinowski, C.T.; Betancor, M.B.; Torrecillas, S.; Sprague, M.; Larroquet, L.; Véron, V.; Panserat, S.; Izquierdo, M.S.; Kaushik, S.J.; Fontagné-Dicharry, S. More Than an Antioxidant: Role of Dietary Astaxanthin on Lipid and Glucose Metabolism in the Liver of Rainbow Trout (Oncorhynchus mykiss). Antioxidants 2023, 12, 136. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.; Shen, X.D.; Gao, F.; Zhao, A.; Freitas, M.C.; Lassman, C.; Luster, A.D.; Busuttil, R.W.; Kupiec-Weglinski, J.W. CXCL10 Regulates Liver Innate Immune Response against Ischemia and Reperfusion Injury. Hepatology 2008, 47, 207–214. [Google Scholar] [CrossRef]
- Hintermann, E.; Bayer, M.; Pfeilschifter, J.M.; Luster, A.D.; Christen, U. CXCL10 Promotes Liver Fibrosis by Prevention of NK Cell Mediated Hepatic Stellate Cell Inactivation. J. Autoimmun. 2010, 35, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Zeremski, M.; Dimova, R.; Astemborski, J.; Thomas, D.L.; Talal, A.H. CXCL9 and CXCL10 Chemokines as Predictors of Liver Fibrosis in a Cohort of Primarily African-American Injection Drug Users with Chronic Hepatitis C. J. Infect. Dis. 2011, 204, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Nieto, N. Ethanol and Fish Oil Induce NFκB Transactivation of the Collagen A2(I) Promoter through Lipid Peroxidation-Driven Activation of the PKC-PI3K-Akt Pathway. Hepatology 2007, 45, 1433–1445. [Google Scholar] [CrossRef]
- Dansou, D.M.; Wang, H.; Nugroho, R.D.; He, W.; Zhao, Q.; Zhang, J. Assessment of Response to Moderate and High Dose Supplementation of Astaxanthin in Laying Hens. Animals 2021, 11, 1138. [Google Scholar] [CrossRef]
- Monmeesil, P.; Fungfuang, W.; Tulayakul, P.; Pongchairerk, U. The Effects of Astaxanthin on Liver Histopathology and Expression of Superoxide Dismutase in Rat Aflatoxicosis. J. Vet. Med. Sci. 2019, 81, 1162–1172. [Google Scholar] [CrossRef]
- Zhu, X.; Hao, R.; Zhang, J.; Tian, C.; Hong, Y.; Zhu, C.; Li, G. Dietary Astaxanthin Improves the Antioxidant Capacity, Immunity and Disease Resistance of Coral Trout (Plectropomus leopardus). Fish Shellfish. Immunol. 2022, 122, 38–47. [Google Scholar] [CrossRef]
- Islam, M.A.; Al Mamun, M.A.; Faruk, M.; Ul Islam, M.T.; Rahman, M.M.; Alam, M.N.; Towheedur Rahman, A.F.M.; Reza, H.M.; Alam, M.A. Astaxanthin Ameliorates Hepatic Damage and Oxidative Stress in Carbon Tetrachloride-Administered Rats. Pharmacogn. Res. 2017, 9, S84–S91. [Google Scholar] [CrossRef]


{kind=link}
| I | II | |
|---|---|---|
| Items | Control | AST |
| Soybean pressed cake | 19 | 19 |
| Wheat | 42.17 | 42.17 |
| Corn | 20 | 20 |
| Rapeseed oil | 0.8 | 0.8 |
| Skimmed milk powder | 10 | 10 |
| Dried whey | 5 | 5 |
| 1-Ca phosphate | 0.4 | 0.4 |
| Feed chalk | 1.15 | 1.15 |
| Salt | 0.13 | 0.13 |
| Lysine | 0.4 | 0.4 |
| Methionine | 0.23 | 0.23 |
| Threonine | 0.16 | 0.16 |
| Tryptophan | 0.06 | 0.06 |
| Vitamin-mineral premix * | 0.5 | 0.5 |
| Astaxanthin (0.025 g/kg) | - | + |
| Content in 1 kg: | ||
| Dry matter, g | 901 | 901 |
| Crude protein, g | 183 | 183 |
| Crude fat, g | 40 | 40 |
| Crude fiber, g | 27 | 27 |
| Crude ash, g | 56 | 56 |
| Metabolizable energy, MJ ** | 13.8 | 13.8 |
| Lysine, g | 13.7 | 13.7 |
| Methionine + Cystine, g | 8.2 | 8.2 |
| Threonine, g | 8.8 | 8.8 |
| Tryptophan, g | 2.7 | 2.7 |
| Calcium, g | 7.6 | 7.6 |
| Phosphorus digestible, g | 3.1 | 3.1 |
| Gene | Primers Sequence | Amplicon Length, bp | |
|---|---|---|---|
| RPS29 | ribosomal protein S29 | Ss03391548_g1 | 71 |
| HK1 | hexokinase 1 | Ss04323446_m1 | 59 |
| NOTCH1 | notch receptor 1 | Ss03377164_u | 71 |
| CYP7A1 | Cytochrome P450 family 7 subfamily A member 1 | Ss03378689_u1 | 119 |
| NR1H3 | Nuclear receptor subfamily 1 group H member 3 | Ss03389237_g1 | 101 |
| CREB1 | cAMP responsive element binding protein 1 | Ss03386122_u1 | 98 |
| SREBF2 | Sterol regulatory element binding transcription factor 2 | Ss03376492_u1 | 67 |
| SIRT3 | Sirtuin 3 | Ss03386766_u1 | 80 |
| DHCR24 | 24-dehydrocholesterol reductase | Ss04323966_m1 | 104 |
| APOE | Apolipoprotein E | Ss03394681_m1 | 63 |
| ACAA2 | Acetyl-CoA acyltransferase 2 | Ss04245775_m1 | 71 |
| CXCL10 | C–X–C motif chemokine ligand 10 | Ss03391845_g1 | 146 |
| CYP1A1 | Cytochrome P450 family 1 subfamily A member 1 | Ss03394917_g1 | 77 |
| COL1A2 | Collagen type I alpha 2 chain | Ss03375009_u1 | 76 |
| SOD1 | Superoxide dismutase 1 | Ss03373476_u1 | 77 |
| CAT | Catalase | Ss04323025_m1 | 56 |
| Gene | Mean RQ ± SE | p-Value | |
|---|---|---|---|
| Group | |||
| I | II | ||
| CYP7A1 | 0.403 ± 0.07 | 0.927 ± 0.14 | 0.001 |
| SREBP2 | 0.881 ± 0.11 | 0.786 ± 0.14 | 0.232 |
| SIRT3 | 0.210 ± 0.05 | 0.395 ± 0.11 | 0.279 |
| APOE | 0.439 ± 0.11 | 0.564 ± 0.22 | 0.867 |
| CREB | 1.701 ± 0.32 | 0.807 ± 0.07 | 0.040 |
| NOTCH1 | 0.433 ± 0.07 | 0.193 ± 0.03 | 0.004 |
| NR1H3 | 0.459 ± 0.08 | 0.134 ± 0.05 | 0.002 |
| DHCR24 | 0.268 ± 0.04 | 0.185 ± 0.04 | 0.121 |
| ACCA2 | 0.196 ± 0.24 | 0.127 ± 0.13 | 0.878 |
| HK1 | 1.271 ± 0.24 | 1.183 ± 0.14 | 0.959 |
| CXCL10 | 0.205 ± 0.04 | 0.158 ± 0.04 | 0.152 |
| CYP1A1 | 3.134 ± 1.28 | 6.112 ± 4.11 | 0.645 |
| COL2A1 | 3.147 ± 0.50 | 3.053 ± 0.52 | 0.954 |
| SOD1 | 1.326 ± 0.18 | 1.638 ± 0.23 | 0.382 |
| CAT | 2.381 ± 0.51 | 2.323 ± 0.34 | 0.878 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szczepanik, K.; Oczkowicz, M.; Dobrowolski, P.; Świątkiewicz, M. The Protective Effects of Astaxanthin (AST) in the Liver of Weaned Piglets. Animals 2023, 13, 3268. https://doi.org/10.3390/ani13203268
Szczepanik K, Oczkowicz M, Dobrowolski P, Świątkiewicz M. The Protective Effects of Astaxanthin (AST) in the Liver of Weaned Piglets. Animals. 2023; 13(20):3268. https://doi.org/10.3390/ani13203268
Chicago/Turabian StyleSzczepanik, Kinga, Maria Oczkowicz, Piotr Dobrowolski, and Małgorzata Świątkiewicz. 2023. "The Protective Effects of Astaxanthin (AST) in the Liver of Weaned Piglets" Animals 13, no. 20: 3268. https://doi.org/10.3390/ani13203268
APA StyleSzczepanik, K., Oczkowicz, M., Dobrowolski, P., & Świątkiewicz, M. (2023). The Protective Effects of Astaxanthin (AST) in the Liver of Weaned Piglets. Animals, 13(20), 3268. https://doi.org/10.3390/ani13203268