
Temperature Elevation during Semen Delivery Deteriorates Boar Sperm Quality by Promoting Apoptosis

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Semen Handling and Experiment Design
2.2. Assessment of Sperm Quality and Functionality
2.3. Determination of Intracellular ATP, ADP, and AMP Content
2.4. Western Blotting
2.5. Statistical Analysis
3. Results
3.1. Effect of Temperature Changes during Delivery on Sperm Quality and Functionality
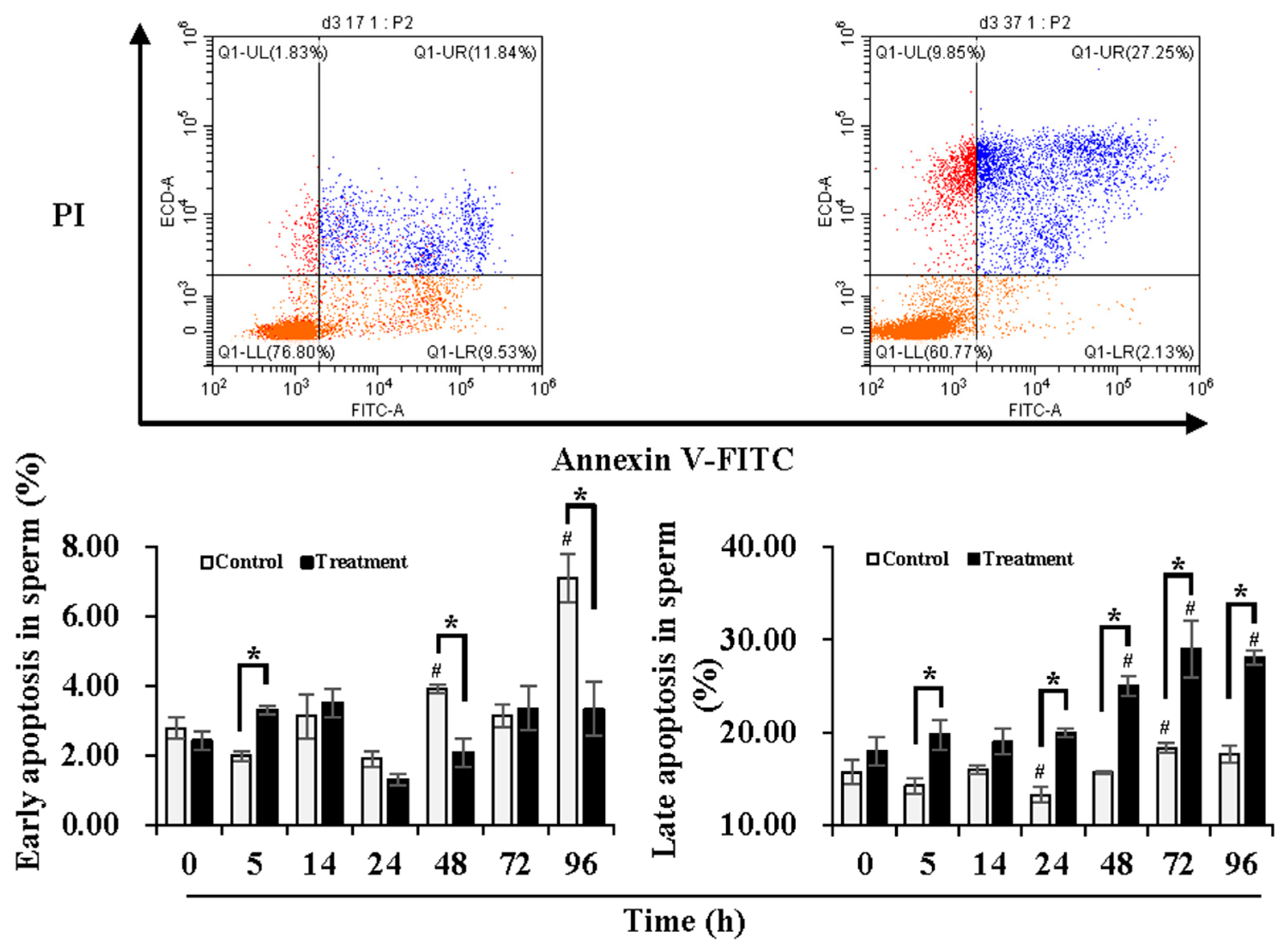
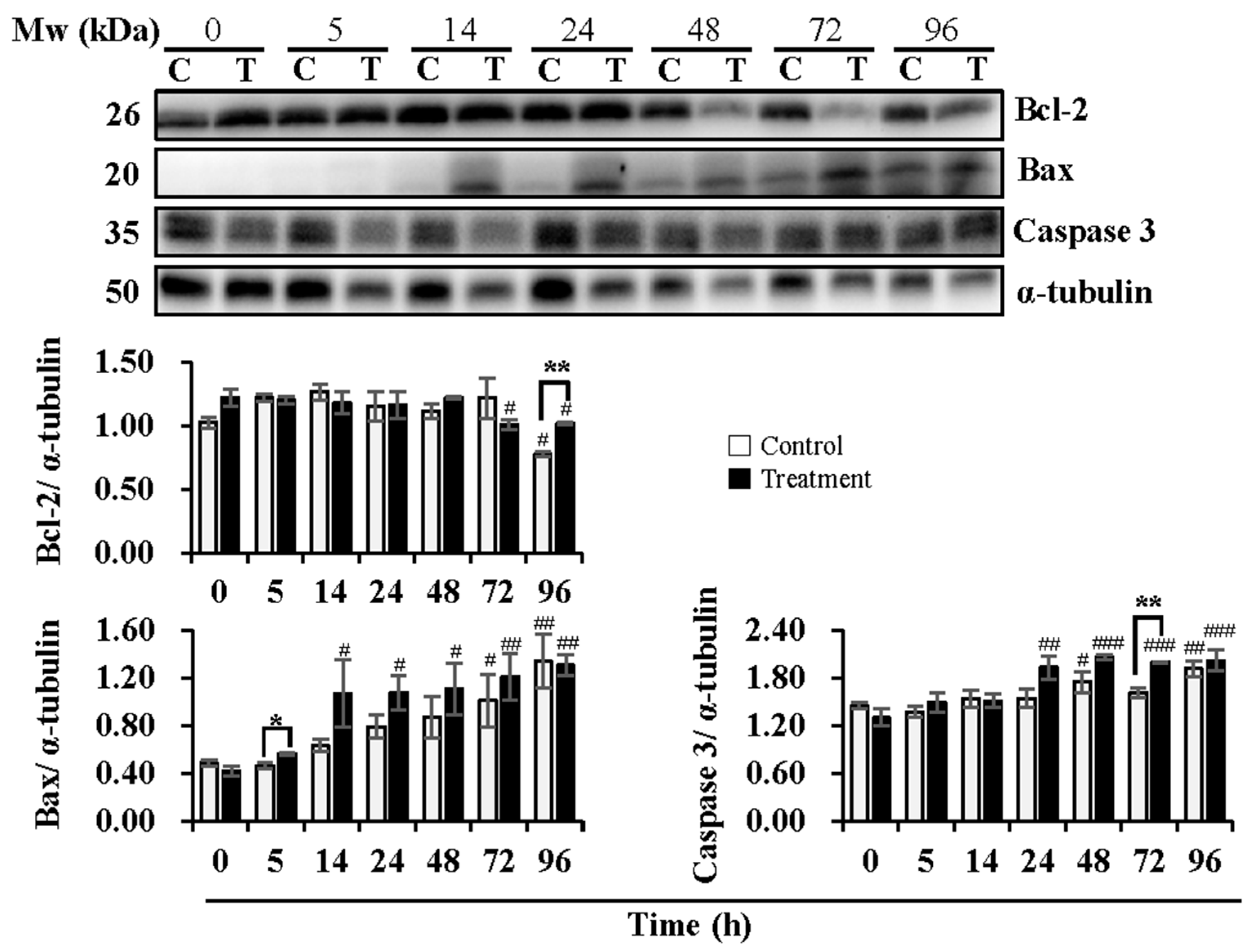
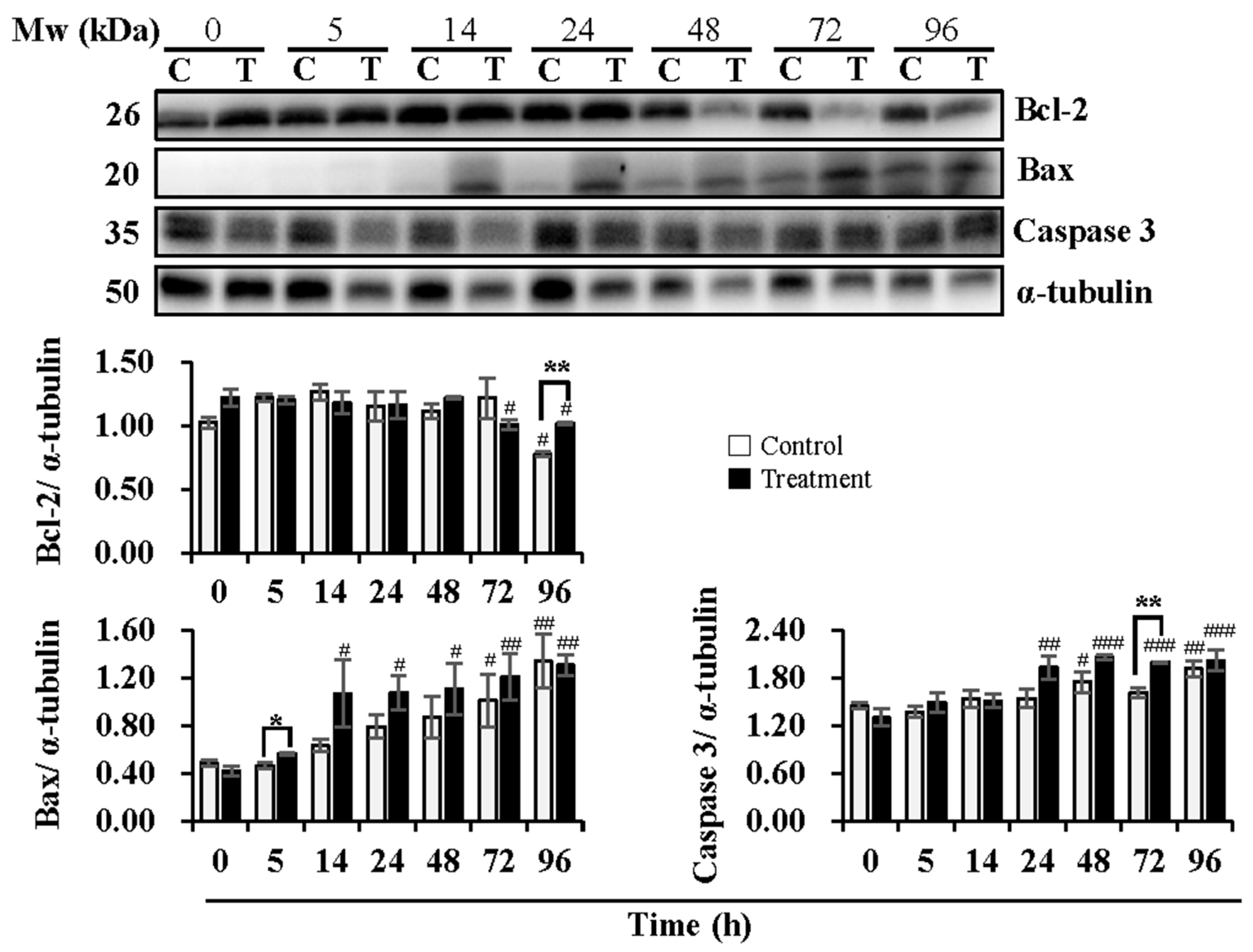
3.2. Effect of Temperature Changes during Delivery and Subsequent Storage (at 17 °C) on Sperm Apoptotic Levels
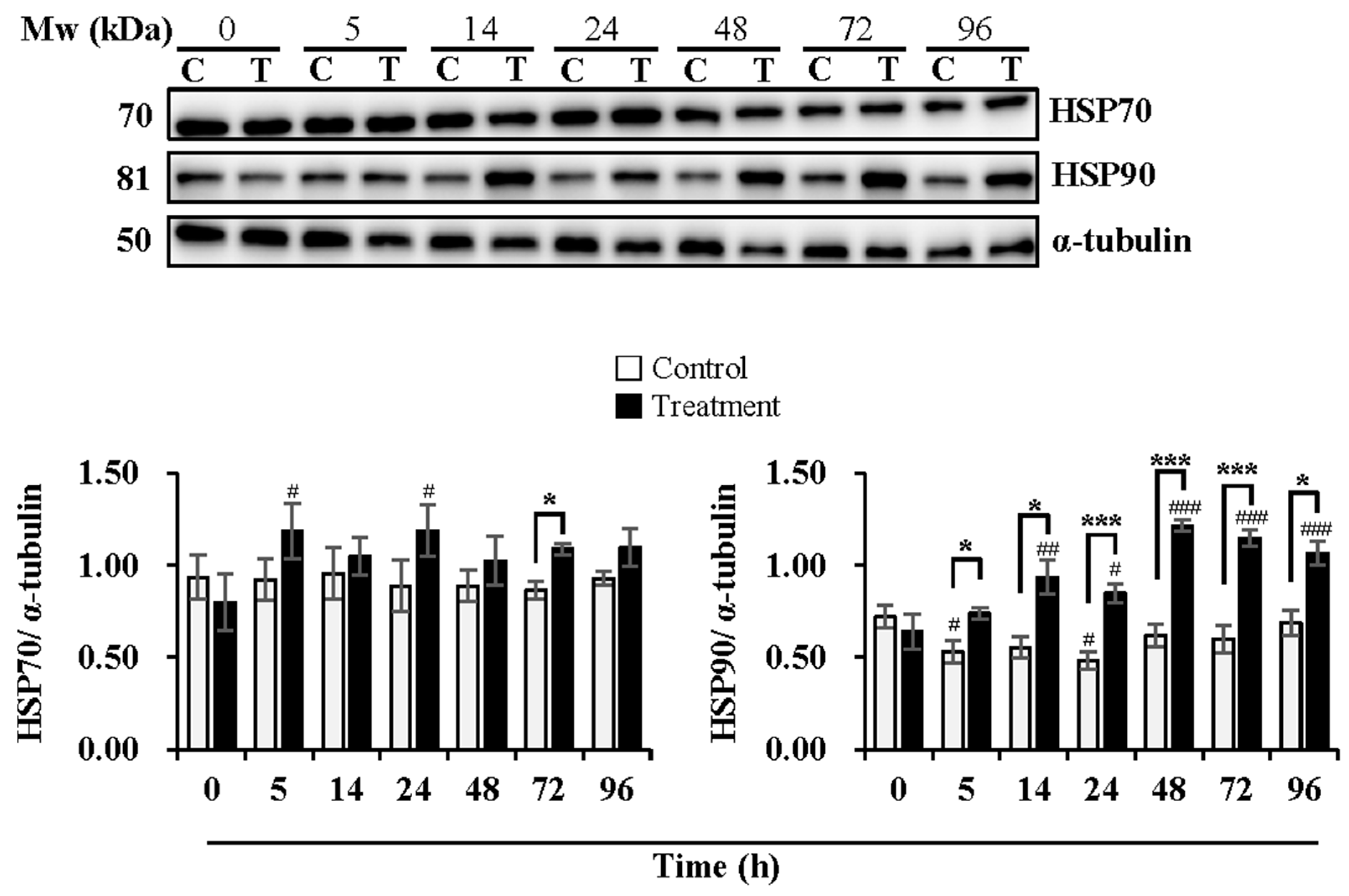
3.3. Effect of Temperature Changes during Delivery and Subsequent Storage (at 17 °C) on Activation of Sperm AMPK and Expression Levels of HSP70 and HSP90
4. Discussion
4.1. Effect of Temperature Changes during Delivery on Sperm Quality and Functionality
4.2. Effect of Temperature Changes during Delivery and Subsequent Storage (at 17 °C) on Sperm Apoptotic Levels
4.3. Effect of Temperature Changes during Delivery and Subsequent Storage (at 17 °C) on Activation of Sperm AMPK and Expression Levels of HSP70 and HSP90
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lopez Rodriguez, A.; Van Soom, A.; Arsenakis, I.; Maes, D. Boar management and semen handling factors affect the quality of boar extended semen. Porc. Health Manag. 2017, 3, 15. [Google Scholar] [CrossRef]
- Collier, R.J.; Renquist, B.J.; Xiao, Y. A 100-Year Review: Stress physiology including heat stress. J. Dairy Sci. 2017, 100, 10367–10380. [Google Scholar] [CrossRef]
- Gonzalez-Castro, R.; Porflidt, C.; Patton, T.; Goins, D.; Herickhoff, L. Effect of season, genetic line and temperature during transport on sperm motility of commercial insemination doses of pooled boar semen: A retrospective study. Reprod. Domest. Anim. 2022, 57, 1363–1374. [Google Scholar] [CrossRef]
- Paschoal, A.F.; Luther, A.M.; Jakop, U.; Schulze, M.; Bortolozzo, F.P.; Waberski, D. Factors influencing the response of spermatozoa to agitation stress: Implications for transport of extended boar semen. Theriogenology 2021, 175, 54–60. [Google Scholar] [CrossRef]
- Iglesias-Carrasco, M.; Harrison, L.; Jennions, M.D.; Head, M.L. Combined effects of rearing and testing temperatures on sperm traits. J. Evol. Biol. 2020, 33, 1715–1724. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.; Li, Z.; Li, C. Mitochondrial OXPHOS is involved in the protective effects of L-arginine against heat-induced low sperm motility of boar. J. Therm. Biol. 2019, 84, 236–244. [Google Scholar] [CrossRef]
- Peña, S.T.; Stone, F.; Gummow, B.; Parker, A.J.; Paris, D.B.B.P. Susceptibility of boar spermatozoa to heat stress using in vivo and in vitro experimental models. Trop. Anim. Health Prod. 2021, 53, 97. [Google Scholar] [CrossRef]
- Kim, B.; Park, K.; Rhee, K. Heat stress response of male germ cells. Cell Mol. Life Sci. 2013, 70, 2623–2636. [Google Scholar] [CrossRef]
- Santiago, J.; Santos, M.A.S.; Fardilha, M.; Silva, J.V. Stress response pathways in the male germ cells and gametes. Mol. Hum. Reprod. 2020, 26, 1–13. [Google Scholar] [CrossRef]
- de Oliveira, M.R.; Nabavi, S.F.; Manayi, A.; Daglia, M.; Hajheydari, Z.; Nabavi, S.M. Resveratrol and the mitochondria: From triggering the intrinsic apoptotic pathway to inducing mitochondrial biogenesis, a mechanistic view. Biochim. Biophys. Acta 2016, 1860, 727–745. [Google Scholar] [CrossRef]
- Yang, X.; Huang, N. Berberine induces selective apoptosis through the AMPK-mediated mitochondrial/caspase pathway in hepatocellular carcinoma. Mol. Med. Rep. 2013, 8, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Hurtado de Llera, A.; Martin-Hidalgo, D.; Rodriguez-Gil, J.E.; Gil, M.C.; Garcia-Marin, L.J.; Bragado, M.J. AMP-activated kinase, AMPK, is involved in the maintenance of plasma membrane organization in boar spermatozoa. Biochim. Biophys. Acta 2013, 1828, 2143–2151. [Google Scholar] [CrossRef] [PubMed]
- Martin-Hidalgo, D.; Hurtado de Llera, A.; Calle-Guisado, V.; Gonzalez-Fernandez, L.; Garcia-Marin, L.; Bragado, M.J. AMPK Function in Mammalian Spermatozoa. Int. J. Mol. Sci. 2018, 19, 3293. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.Y.; Zhou, X.D.; Yang, J.; Chen, L.X.; Ran, D.H. Inhibition of autophagy enhances heat-induced apoptosis in human non-small cell lung cancer cells through ER stress pathways. Arch. Biochem. Biophys. 2016, 607, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Shen, X.; Shen, F.; Zhong, W.; Wu, H.; Liu, S.; Lai, J. TAK1 activates AMPK-dependent cell death pathway in hydrogen peroxide-treated cardiomyocytes, inhibited by heat shock protein-70. Mol. Cell. Biochem. 2013, 377, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yu, Q.; Chen, J.; Deng, B.; Qian, L.; Le, Y. PP2A mediated AMPK inhibition promotes HSP70 expression in heat shock response. PLoS ONE 2010, 5, e13096. [Google Scholar] [CrossRef] [PubMed]
- Boudeau, J.; Deak, M.; Lawlor, M.A.; Morrice, N.A.; Alessi, D.R. Heat-shock protein 90 and Cdc37 interact with LKB1 and regulate its stability. Biochem. J. 2003, 370, 849–857. [Google Scholar] [CrossRef]
- Purandhar, K.; Jena, P.K.; Prajapati, B.; Rajput, P.; Seshadri, S. Understanding the role of heat shock protein isoforms in male fertility, aging and apoptosis. World J. Mens. Health 2014, 32, 123–132. [Google Scholar] [CrossRef]
- Zannoni, A.; Bernardini, C.; Zaniboni, A.; Ferlizza, E.; Ventrella, D.; Bacci, M.L.; Forni, M. Relative abundance of heat shock proteins and clusterin transcripts in spermatozoa collected from boar routinely utilised in an artificial insemination centre: Preliminary results. Vet. Res. Commun. 2017, 41, 233–239. [Google Scholar] [CrossRef]
- Jego, G.; Hazoumé, A.; Seigneuric, R.; Garrido, C. Targeting heat shock proteins in cancer. Cancer Lett. 2013, 332, 275–285. [Google Scholar] [CrossRef]
- Li, J.; Zhao, W.; Zhu, J.; Ju, H.; Liang, M.; Wang, S.; Chen, S.; Ferreira-Dias, G.; Liu, Z. Antioxidants and Oxidants in Boar Spermatozoa and Their Surrounding Environment Are Associated with AMPK Activation during Liquid Storage. Vet. Sci. 2023, 10, 214. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, H.D.; Welch, G.R. Determination of intracellular reactive oxygen species and high mitochondrial membrane potential in Percoll-treated viable boar sperm using fluorescence-activated flow cytometry. J. Anim. Sci. 2006, 84, 2089–2100. [Google Scholar] [CrossRef] [PubMed]
- Pena, F.J.; Johannisson, A.; Wallgren, M.; Rodriguez-Martinez, H. Assessment of fresh and frozen-thawed boar semen using an Annexin-V assay: A new method of evaluating sperm membrane integrity. Theriogenology 2003, 60, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.T.; Wallner, U.; Schmicke, M.; Waberski, D.; Henning, H. Energy metabolic state in hypothermically stored boar spermatozoa using a revised protocol for efficient ATP extraction. Biol. Open 2016, 5, 1743–1751. [Google Scholar] [CrossRef] [PubMed]
- Mellagi, A.P.G.; Will, K.J.; Quirino, M.; Bustamante-Filho, I.C.; Ulguim, R.D.R.; Bortolozzo, F.P. Update on artificial insemination: Semen, techniques, and sow fertility. Mol. Reprod. Dev. 2022, 90, 601–611. [Google Scholar] [CrossRef]
- Schulze, M.; Bortfeldt, R.; Schäfer, J.; Jung, M.; Fuchs-Kittowski, F. Effect of vibration emissions during shipping of artificial insemination doses on boar semen quality. Anim. Reprod. Sci. 2018, 192, 328–334. [Google Scholar] [CrossRef]
- Morrell, J.M. Heat stress and bull fertility. Theriogenology 2020, 153, 62–67. [Google Scholar] [CrossRef]
- Rizzoto, G.; Boe-Hansen, G.; Klein, C.; Thundathil, J.C.; Kastelic, J.P. Acute mild heat stress alters gene expression in testes and reduces sperm quality in mice. Theriogenology 2020, 158, 375–381. [Google Scholar] [CrossRef]
- Lyrio, L.L.; Lazaro, M.A.; Sonegheti, R.; Moulin, L.; Coslop, L.; Sarto, C.G.; Loureiro, B.; Favoreto, M.G. Effects of heat stress on sperm quality of French Bulldogs. Theriogenology 2023, 199, 131–137. [Google Scholar] [CrossRef]
- Song, Y.; Zhao, X.; Aihemaiti, A.; Haire, A.; Gao, Y.; Niu, C.; Yang, P.; Liu, G.; Jia, G.; Wusiman, A. The Mechanism of Heat Stress Resistance During Spermatogenesis in Turpan Black Sheep. Front. Vet. Sci. 2022, 9, 846981. [Google Scholar] [CrossRef]
- Einarsson, S.; Brandt, Y.; Lundeheim, N.; Madej, A. Stress and its influence on reproduction in pigs: A review. Acta Vet. Scand. 2008, 50, 48. [Google Scholar] [CrossRef] [PubMed]
- Cerolini, S.; Maldjian, A.; Surai, P.; Noble, R. Viability, susceptibility to peroxidation and fatty acid composition of boar semen during liquid storage. Anim. Reprod. Sci. 2000, 58, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Casas, I.; Althouse, G.C. The protective effect of a 17 °C holding time on boar sperm plasma membrane fluidity after exposure to 5 °C. Cryobiology 2013, 66, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Canvin, A.T.; Buhr, M.M. Effect of temperature on the fluidity of boar sperm membranes. J. Reprod. Fertil. 1989, 85, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.X.; Yang, Z.M. Evaluation on sperm quality of freshly ejaculated boar semen during in vitro storage under different temperatures. Theriogenology 2000, 53, 1477–1488. [Google Scholar] [CrossRef]
- Henning, H.; Nguyen, Q.T.; Wallner, U.; Waberski, D. Temperature limits for storage of extended boar semen from the perspective of the sperm’s energy status. Front. Vet. Sci. 2022, 9, 953021. [Google Scholar] [CrossRef]
- da Silva, D.F.; Rodrigues, T.A.; da Silveira, J.C.; Gonella-Diaza, A.M.; Binelli, M.; Lopes, J.V.; Moura, M.T.; Feitosa, W.B.; Paula-Lopes, F.F. Cellular responses and microRNA profiling in bovine spermatozoa under heat shock. Reproduction 2022, 164, 155–168. [Google Scholar] [CrossRef]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- Durairajanayagam, D.; Agarwal, A.; Ong, C. Causes, effects and molecular mechanisms of testicular heat stress. Reprod. Biomed. Online 2015, 30, 14–27. [Google Scholar] [CrossRef]
- Aitken, R.J.; Jones, K.T.; Robertson, S.A. Reactive oxygen species and sperm function--in sickness and in health. J. Androl. 2012, 33, 1096–1106. [Google Scholar] [CrossRef]
- Moazamian, R.; Polhemus, A.; Connaughton, H.; Fraser, B.; Whiting, S.; Gharagozloo, P.; Aitken, R.J. Oxidative stress and human spermatozoa: Diagnostic and functional significance of aldehydes generated as a result of lipid peroxidation. Mol. Hum. Reprod. 2015, 21, 502–515. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Xi, H.; Zhang, Z.; Liang, Y.; Li, Q.; He, J. Germ cell apoptosis and expression of Bcl-2 and Bax in porcine testis under normal and heat stress conditions. Acta Histochem. 2017, 119, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Parrish, J.J.; Willenburg, K.L.; Gibbs, K.M.; Yagoda, K.B.; Krautkramer, M.M.; Loether, T.M.; Melo, F.C.S.A. Scrotal insulation and sperm production in the boar. Mol. Reprod. Dev. 2017, 84, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Capela, L.; Leites, I.; Romão, R.; Lopes-da-Costa, L.; Pereira, R.M.L.N. Impact of Heat Stress on Bovine Sperm Quality and Competence. Animals 2022, 12, 975. [Google Scholar] [CrossRef]
- Vera, Y.; Rodriguez, S.; Castanares, M.; Lue, Y.; Atienza, V.; Wang, C.; Swerdloff, R.S.; Sinha Hikim, A.P. Functional role of caspases in heat-induced testicular germ cell apoptosis. Biol. Reprod. 2005, 72, 516–522. [Google Scholar] [CrossRef]
- Matsuki, S.; Iuchi, Y.; Ikeda, Y.; Sasagawa, I.; Tomita, Y.; Fujii, J. Suppression of cytochrome c release and apoptosis in testes with heat stress by minocycline. Biochem. Biophys. Res. Commun. 2003, 312, 843–849. [Google Scholar] [CrossRef]
- Wang, S.; Song, P.; Zou, M.H. AMP-activated protein kinase, stress responses and cardiovascular diseases. Clin. Sci. 2012, 122, 555–573. [Google Scholar] [CrossRef]
- Chen, M.B.; Wu, X.Y.; Gu, J.H.; Guo, Q.T.; Shen, W.X.; Lu, P.H. Activation of AMP-activated protein kinase contributes to doxorubicin-induced cell death and apoptosis in cultured myocardial H9c2 cells. Cell Biochem. Biophys. 2011, 60, 311–322. [Google Scholar] [CrossRef]
- Chen, L.; Xu, B.; Liu, L.; Luo, Y.; Yin, J.; Zhou, H.; Chen, W.; Shen, T.; Han, X.; Huang, S. Hydrogen peroxide inhibits mTOR signaling by activation of AMPKalpha leading to apoptosis of neuronal cells. Lab. Investig. 2010, 90, 762–773. [Google Scholar] [CrossRef]
- Hurtado de Llera, A.; Martin-Hidalgo, D.; Gil, M.C.; Garcia-Marin, L.J.; Bragado, M.J. AMP-activated kinase AMPK is expressed in boar spermatozoa and regulates motility. PLoS ONE 2012, 7, e38840. [Google Scholar] [CrossRef]
- Min, L.; Cheng, J.B.; Shi, B.L.; Yang, H.J.; Zheng, N.; Wang, J.Q. Effects of heat stress on serum insulin, adipokines, AMP-activated protein kinase, and heat shock signal molecules in dairy cows. J. Zhejiang Univ. Sci. B 2015, 16, 541–548. [Google Scholar] [CrossRef]
- Adesina, P.A.; Saeki, I.; Yamamoto, Y.; Suzuki, T. N-butyrate increases heat shock protein 70 through heat shock factor 1 and AMP-activated protein kinase pathways in human intestinal Caco-2 cells. Arch. Biochem. Biophys. 2023, 736, 109525. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Lam, K.K.; Peng, Y.J.; Lee, Y.M.; Yang, C.Y.; Tsai, Y.J.; Yen, M.H.; Cheng, P.Y. Heat shock protein 70 and AMP-activated protein kinase contribute to 17-DMAG-dependent protection against heat stroke. J. Cell. Mol. Med. 2016, 20, 1889–1897. [Google Scholar] [CrossRef] [PubMed]
- Ferlin, A.; Speltra, E.; Patassini, C.; Pati, M.A.; Garolla, A.; Caretta, N.; Foresta, C. Heat shock protein and heat shock factor expression in sperm: Relation to oligozoospermia and varicocele. J. Urol. 2010, 183, 1248–1252. [Google Scholar] [CrossRef] [PubMed]
- Saibil, H. Chaperone machines for protein folding, unfolding and disaggregation. Nat. Rev. Mol. Cell Biol. 2013, 14, 630–642. [Google Scholar] [CrossRef] [PubMed]
- Lymbery, R.A.; Evans, J.P.; Kennington, W.J. Post-ejaculation thermal stress causes changes to the RNA profile of sperm in an external fertilizer. Proc. Biol. Sci. 2020, 287, 20202147. [Google Scholar] [CrossRef] [PubMed]
- Parmar, K.H.; Singh, V.; Savsani, H.H.; Kavani, F.S.; Rajoriya, J.S.; Lone, S.A. Seasonal variation in heat shock proteins (hsp70 and hsp90) and their association with frozen semen quality and fertility in buffaloes. Cryo Lett. 2021, 42, 261–266. [Google Scholar]
- Ikwegbue, P.C.; Masamba, P.; Oyinloye, B.E.; Kappo, A.P. Roles of Heat Shock Proteins in Apoptosis, Oxidative Stress, Human Inflammatory Diseases, and Cancer. Pharmaceuticals 2017, 11, 2. [Google Scholar] [CrossRef]
- Spinaci, M.; Volpe, S.; Bernardini, C.; De Ambrogi, M.; Tamanini, C.; Seren, E.; Galeati, G. Immunolocalization of heat shock protein 70 (Hsp 70) in boar spermatozoa and its role during fertilization. Mol. Reprod. Dev. 2005, 72, 534–541. [Google Scholar] [CrossRef]
- Varghese, T.; Divyashree, B.C.; Roy, S.C.; Roy, K.S. Loss of heat shock protein 70 from apical region of buffalo (Bubalus bubalis) sperm head after freezing and thawing. Theriogenology 2016, 85, 828–834. [Google Scholar] [CrossRef]
- Volpe, S.; Galeati, G.; Bernardini, C.; Tamanini, C.; Mari, G.; Zambelli, D.; Seren, E.; Spinaci, M. Comparative immunolocalization of heat shock proteins (Hsp)-60, -70, -90 in boar, stallion, dog and cat spermatozoa. Reprod. Domest. Anim. 2008, 43, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Calle-Guisado, V.; Bragado, M.J.; García-Marín, L.J.; González-Fernández, L. HSP90 maintains boar spermatozoa motility and mitochondrial membrane potential during heat stress. Anim. Reprod. Sci. 2017, 187, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009, 14, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Garrido, C.; Brunet, M.; Didelot, C.; Zermati, Y.; Schmitt, E.; Kroemer, G. Heat shock proteins 27 and 70: Anti-apoptotic proteins with tumorigenic properties. Cell Cycle 2006, 5, 2592–2601. [Google Scholar] [CrossRef] [PubMed]
- Gallardo-Soler, A.; Macías-García, B.; García-Marín, L.J.; Bragado, M.J.; González-Fernández, L. Effect of boar semen supplementation with recombinant heat shock proteins during summer. Anim. Reprod. Sci. 2019, 211, 106227. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Motility, % | Progressive Motility, % | ||||
|---|---|---|---|---|---|
| Control | Treatment | Control | Treatment | ||
| During delivery | 0 h | 80.00 ± 5.64 a | 74.33 ± 8.09 a | 45.00 ± 5.86 a | 42.00 ± 5.51 a |
| 5 h | 81.33 ± 8.01 a | 77.33 ± 6.96 a | 45.33 ± 7.54 a | 41.67 ± 5.36 a | |
| 14 h | 86.33 ± 3.48 a | 80.33 ± 9.70 a | 50.33 ± 2.03 a | 53.33 ± 6.69 a | |
| 24 h | 85.33 ± 5.90 ax | 5.00 ± 3.22 by | 49.00 ± 3.61 ax | 0.67 ± 0.67 by | |
| Subsequent storage at 17 °C | 48 h | 85.00 ± 7.21 ax | 3.33 ± 1.76 by | 47.67 ± 7.84 ax | 0.33 ± 0.33 by |
| 72 h | 46.33 ± 5.04 bx | 2.33 ± 1.16 by | 26.00 ± 4.04 bx | 0.33 ± 0.33 by | |
| 96 h | 9.00 ± 3.22 c | 2.00 ± 1.00 b | 2.67 ± 1.45 c | 0.00 ± 0.00 b | |
| Sperm Parameters | Delivery Time | ||||
|---|---|---|---|---|---|
| 0 h | 5 h | 14 h | 24 h | ||
| Viability, % | Control | 72.17 ± 0.48 a | 64.08 ± 1.03 a | 74.28 ± 0.98 bx | 62.49 ± 1.43 bx |
| Treatment | 69.44 ± 1.45 a | 62.26 ± 3.17 b | 58.06 ± 2.09 by | 48.29 ± 1.19 by | |
| Percentage of viable sperm with damaged acrosome, % | Control | 0.07 ± 0.02 a | 0.15 ± 0.04 ab | 0.19 ± 0.04 bx | 0.16 ± 0.03 ab |
| Treatment | 0.11 ± 0.02 a | 0.12 ± 0.03 a | 0.35 ± 0.03 by | 0.28 ± 0.07 b | |
| Intracellular ROS production, 106 fluorescence units/106 viable sperm | Control | 5.97 ± 0.47 ax | 3.96 ± 0.22 b | 2.54 ± 0.09 cx | 2.88 ± 0.32 bc |
| Treatment | 4.42 ± 0.39 y | 3.41 ± 0.10 | 4.02 ± 0.56 y | 3.42 ± 0.45 | |
| Mitochondrial membrane potential of viable sperm, % | Control | 53.10 ± 4.90 a | 60.02 ± 5.17 ab | 63.92 ± 2.52 ab | 68.44 ± 2.81 b |
| Treatment | 55.35 ± 2.27 a | 52.79 ± 8.37 ab | 68.91 ± 2.24 ab | 74.98 ± 0.82 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Zhao, W.; Zhu, J.; Wang, S.; Ju, H.; Chen, S.; Basioura, A.; Ferreira-Dias, G.; Liu, Z. Temperature Elevation during Semen Delivery Deteriorates Boar Sperm Quality by Promoting Apoptosis. Animals 2023, 13, 3203. https://doi.org/10.3390/ani13203203
Li J, Zhao W, Zhu J, Wang S, Ju H, Chen S, Basioura A, Ferreira-Dias G, Liu Z. Temperature Elevation during Semen Delivery Deteriorates Boar Sperm Quality by Promoting Apoptosis. Animals. 2023; 13(20):3203. https://doi.org/10.3390/ani13203203
Chicago/Turabian StyleLi, Junwei, Wenming Zhao, Jiaqiao Zhu, Shuaibiao Wang, Huiming Ju, Shufang Chen, Athina Basioura, Graça Ferreira-Dias, and Zongping Liu. 2023. "Temperature Elevation during Semen Delivery Deteriorates Boar Sperm Quality by Promoting Apoptosis" Animals 13, no. 20: 3203. https://doi.org/10.3390/ani13203203
APA StyleLi, J., Zhao, W., Zhu, J., Wang, S., Ju, H., Chen, S., Basioura, A., Ferreira-Dias, G., & Liu, Z. (2023). Temperature Elevation during Semen Delivery Deteriorates Boar Sperm Quality by Promoting Apoptosis. Animals, 13(20), 3203. https://doi.org/10.3390/ani13203203