
Wing Shape Variation between Terrestrial and Coastal Populations of the Invasive Box Tree Moth, Cydalima perspectalis, in Croatia

 ,
,  ,
, 
 , , , ,
, , , , 
Abstract
:Featured Application
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Identification
2.2. Data Acquisition
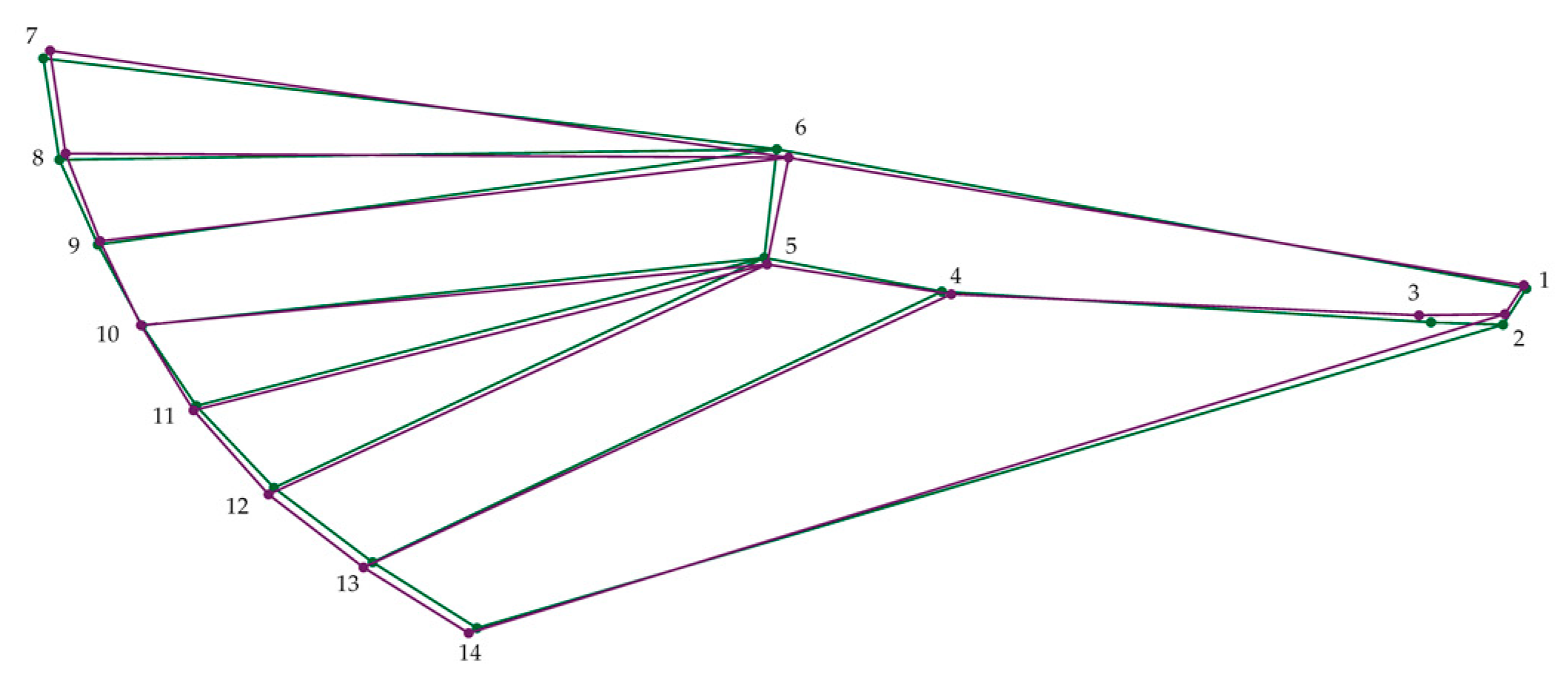
2.3. Designation of Specific Landmarks
2.4. Data Analysis
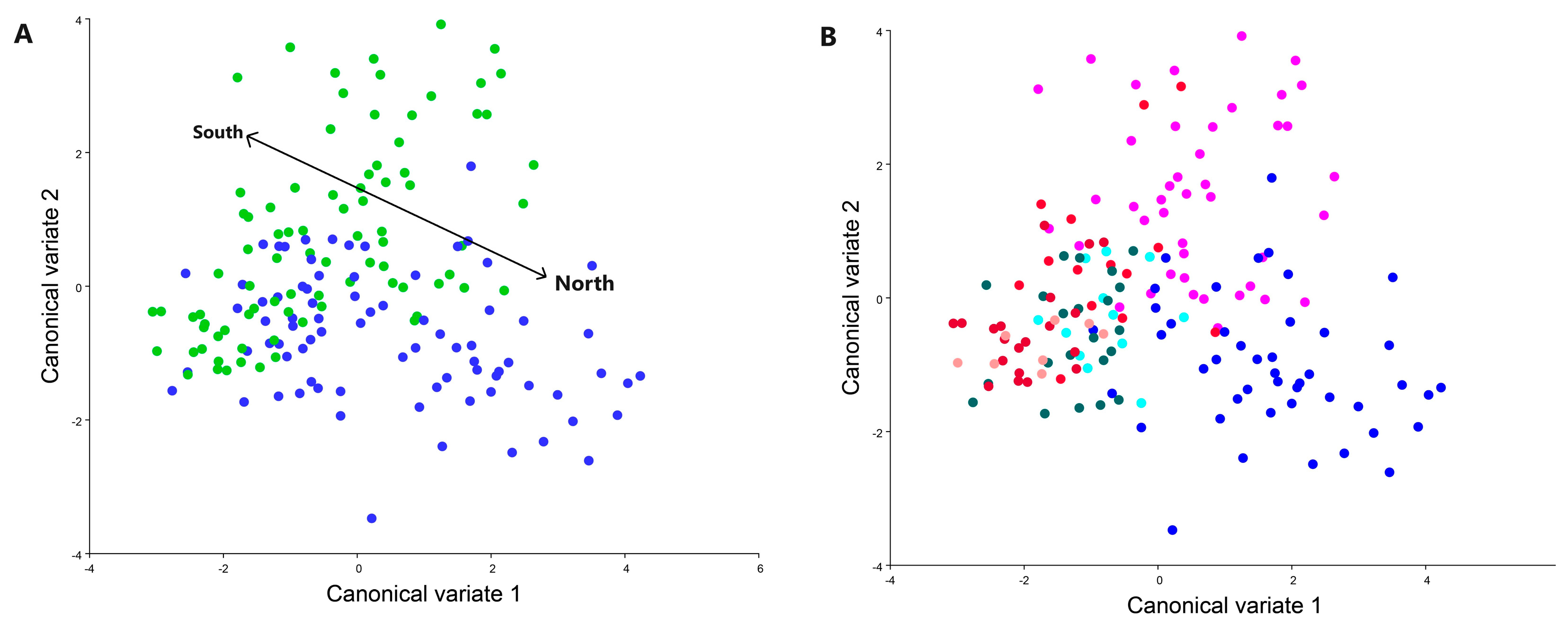
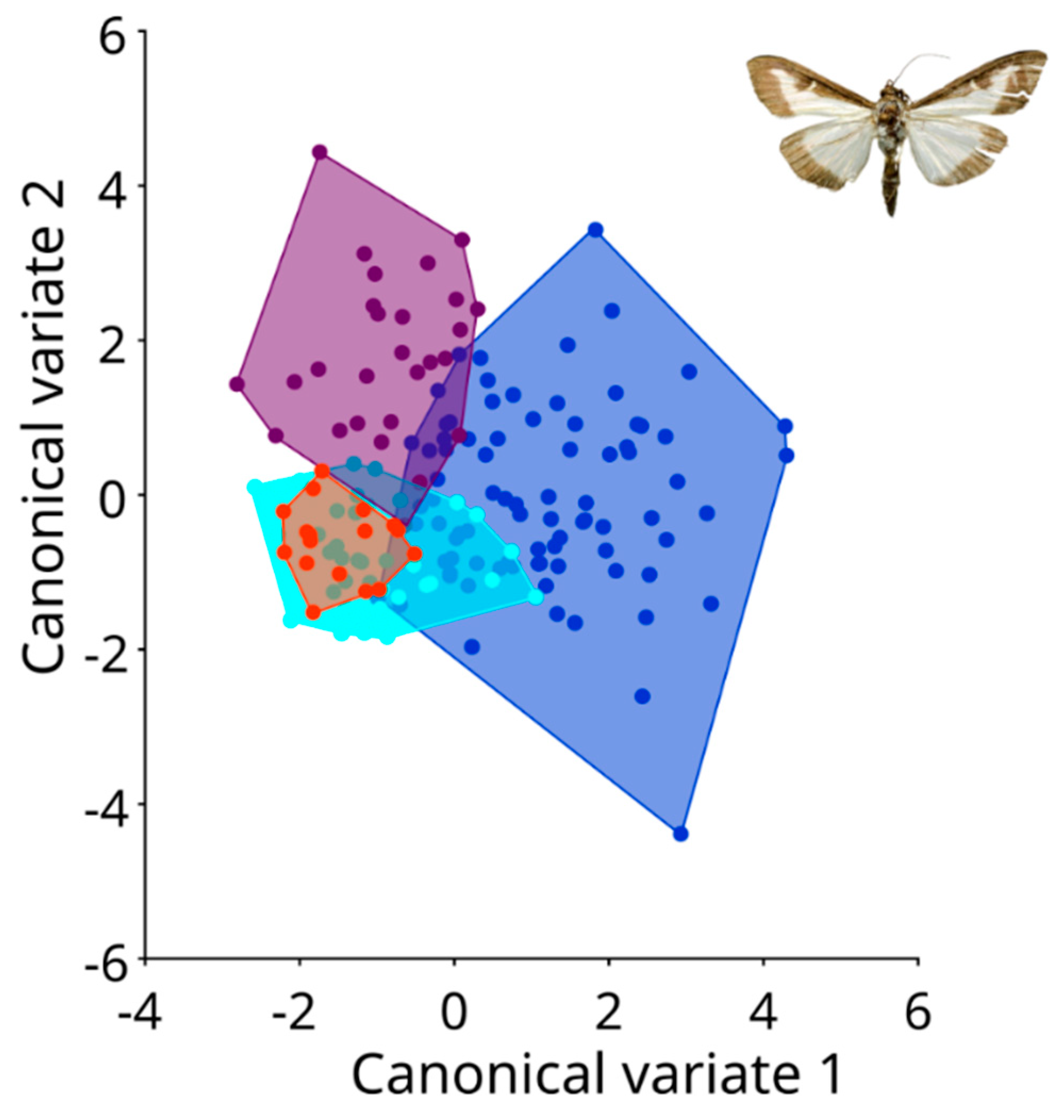
3. Results
4. Discussion
- Distinct Geographical Wing Shape Disparities: Our research convincingly establishes significant differences in wing shapes between terrestrial (Zagreb, Sveti Ivan Zelina, Garešnica) and coastal (Kaštela, Seget, Split, Sinj) populations of C. perspectalis. This underscores the pivotal role of wing morphology in influencing the species’ spread.
- Intraspecific Variation: Within terrestrial populations, notable differences were observed between Garešnica, Zagreb, and Sveti Ivan Zelina. Similarly, discernible wing shape variations were evident among coastal populations, including Split, Seget, Sinj, and Kaštela. These nuanced findings underscore the complexity of the species’ adaptation and dispersal mechanisms.
- Exploring Sexual Dimorphism: Despite marked variability in the upper wings of male and female C. perspectalis individuals, our study does not definitively establish sexual dimorphism in wing shape. This suggests the potential involvement of other factors driving the observed variations.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, J.R.; Dormontt, E.E.; Prentis, P.J.; Lowe, A.J.; Richardson, D.M. Something in the way you move: Dispersal pathways affect invasion success. Trends Ecol. Evol. 2009, 24, 136–144. [Google Scholar] [CrossRef]
- Meurisse, N.; Rassati, D.; Hurley, B.P.; Brockerhoff, E.G.; Haack, R.A. Common pathways by which non-native forest insects move internationally and domestically. J. Pest Sci. 2019, 92, 13–27. [Google Scholar] [CrossRef]
- Hulme, P.E. Unwelcome exchange: International trade as a direct and indirect driver of biological invasions worldwide. One Earth 2021, 4, 666–679. [Google Scholar] [CrossRef]
- Tay, W.T.; Gordon, K.H.J. Going global–genomic insights into insect invasions. Curr. Opin. Insect Sci. 2019, 31, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Roques, A.; Auger-Rozenberg, M.-A.; Blackburn, T.M.; Garnas, J.; Pyšek, P.; Rabitsch, W.; Richardson, D.M.; Wingfield, M.J.; Liebhold, A.M.; Duncan, R.P. Temporal and interspecific variation in rates of spread for insect species invading Europe during the last 200 years. Biol. Invasions 2016, 18, 907–920. [Google Scholar] [CrossRef]
- Estoup, A.; Guillemaud, T. Reconstructing routes of invasion using genetic data: Why, how and so what? Mol. Ecol. 2010, 19, 4113–4130. [Google Scholar] [CrossRef]
- Dlugosch, K.M.; Anderson, S.R.; Braasch, J.; Cang, F.A.; Gillette, H.D. The devil is in the details: Genetic variation in introduced populations and its contributions to invasion. Mol. Ecol. 2015, 24, 2095–2111. [Google Scholar] [CrossRef]
- Lombaert, E.; Guillemaud, T.; Cornuet, J.-M.; Malausa, T.; Facon, B.; Estoup, A. Bridgehead effect in the worldwide invasion of the biocontrol harlequin ladybird. PLoS ONE 2010, 5, e9743. [Google Scholar] [CrossRef] [PubMed]
- Araujo, R.N.; Pereira, M.H.; Soares, A.C.; Pereira, I.D.; Diotaiuti, L.; Gontijo, N.F.; Lehane, M.J.; Guarneri, A.A. Effect of intestinal erythrocyte agglutination on the feeding performance of Triatoma brasiliensis (Hemiptera: Reduviidae). J. Insect Physiol. 2009, 55, 862–868. [Google Scholar] [CrossRef]
- Matošević, D. Box tree moth (Cydalima perspectalis, Lepidoptera; Crambidae), new invasive insect pest in Croatia. South-East Eur. For. SEEFOR 2013, 4, 89–94. [Google Scholar] [CrossRef]
- Bras, A.; Lombaert, E.; Kenis, M.; Li, H.; Bernard, A.; Rousselet, J.; Roques, A.; Auger-Rozenberg, M.-A. The fast invasion of Europe by the box tree moth: An additional example coupling multiple introduction events, bridgehead effects and admixture events. Biol. Invasions 2022, 24, 3865–3883. [Google Scholar] [CrossRef]
- Krüger, E. Glyphodes perspectalis (Walker, 1859)-new for the European fauna (Lepidoptera: Crambidae). Entomol. Z. Mit Insekten-Börse 2008, 118, 81–83. [Google Scholar]
- Mally, R.; Nuss, M. Phylogeny and nomenclature of the box tree moth, Cydalima perspectalis (Walker, 1859) comb. n., which was recently introduced into Europe (Lepidoptera: Pyraloidea: Crambidae: Spilomelinae). Eur. J. Èntomol. 2010, 107, 393–400. [Google Scholar] [CrossRef]
- Koren, T.; Črne, M. The first record of the box tree moth, Cydalima perspectalis (Walker, 1859) (Lepidoptera, Crambidae) in Croatia. Nat. Croat. Period. Musei Hist. Nat. Croat. 2012, 21, 507–510. [Google Scholar]
- Ostojić, I.; Zovko, M.; Petrović, D.; Elez, D. Novi Nalazi Šimširova Moljca Cydalima perspectalis (Walker, 1859.) u Bosni i Hercegovini. Rad. Poljoprivredno- prehrambenog fak. Univ. Sarajevu. 2015, 65, 139–143. [Google Scholar]
- Canelles, Q.; Bassols, E.; Vayreda, J.; Brotons, L. Predicting the potential distribution and forest impact of the invasive species Cydalima perspectalis in Europe. Ecol. Evol. 2021, 11, 5713–5727. [Google Scholar] [CrossRef]
- Nacambo, S.; Leuthardt, F.L.; Wan, H.; Li, H.; Haye, T.; Baur, B.; Weiss, R.M.; Kenis, M. Development characteristics of the box-tree moth Cydalima perspectalis and its potential distribution in Europe. J. Appl. Entomol. 2014, 138, 14–26. [Google Scholar] [CrossRef]
- Maruyama, T.; Shinkaji, N. The life-cycle of the box-tree pyralid, Glyphodes perspectalis (Walker) (Lepidoptera: Pyralidae). II. Developmental characteristics of larvae. Jpn. J. Appl. Entomol. Zool. 1991, 35, 221–230. [Google Scholar] [CrossRef]
- Bras, A.; Avtzis, D.N.; Kenis, M.; Li, H.; Vétek, G.; Bernard, A.; Courtin, C.; Rousselet, J.; Roques, A.; Auger-Rozenberg, M.-A. A complex invasion story underlies the fast spread of the invasive box tree moth (Cydalima perspectalis) across Europe. J. Pest. Sci. 2019, 92, 1187–1202. [Google Scholar] [CrossRef]
- Ribeiro, G., Jr.; Reis, J.; Vaccarezza, F.; de Oliveira Silva, A.C.; Cardoso Lanza, F.; Paim Miranda, D.L.; Gurgel-Gonçalves, R.; Reis, M.G. Sometimes, the Size Matters: Wing Geometric Morphometrics as a Tool to Assess House Invasion and Colonization by Triatoma sordida (Stäl 1859); SSRN: Amsterdam, The Netherlands, 2023. [Google Scholar]
- Zelditch, M.L.; Swiderski, D.L.; Sheets, H.D.; Fink, W.L. Geometric Morphometrics for Biologists: A Primer; Academic Press: Cambridge, MA, USA, 2004. [Google Scholar]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Bookstein, F.L. Biometrics, biomathematics and the morphometric synthesis. Bull. Math. Biol. 1996, 58, 313–365. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Marcus, L.F. A revolution in morphometrics. Trends Ecol. Evol. 1993, 8, 129–132. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, W.; Benítez, H.; Addison, P. The use of geometric morphometric analysis to illustrate the shape change induced by different fruit hosts on the wing shape of Bactrocera dorsalis and Ceratitis capitata (Diptera: Tephritidae). Zool. Anz. 2017, 269, 110–116. [Google Scholar] [CrossRef]
- Mikac, K.M.; Lemic, D.; Benítez, H.A.; Bažok, R. Changes in corn rootworm wing morphology are related to resistance development. J. Pest Sci. 2019, 92, 443–451. [Google Scholar] [CrossRef]
- Mikac, K.M.; Lemic, D.; Bažok, R.; Benítez, H.A. Wing shape changes: A morphological view of the Diabrotica virgifera virgifera European invasion. Biol. Invasions 2016, 18, 3401–3407. [Google Scholar] [CrossRef]
- Lemic, D.; Benítez, H.A.; Bjeliš, M.; Órdenes-Claveria, R.; Ninčević, P.; Mikac, K.M.; Živković, I.P. Agroecological effect and sexual shape dimorphism in medfly Ceratitis capitata (Diptera: Tephritidae) an example in Croatian populations. Zool. Anz.—A J. Comp. Zool. 2020, 288, 118–124. [Google Scholar] [CrossRef]
- Espinoza-Donoso, S.; Angulo-Bedoya, M.; Lemic, D.; Benítez, H.A. Assessing the influence of allometry on sexual and non-sexual traits: An example in Cicindelidia trifasciata (Coleoptera: Cicindelinae) using geometric morphometrics. Zool. Anz.—A J. Comp. Zool. 2020, 287, 61–66. [Google Scholar] [CrossRef]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. Geometric morphometrics: Ten years of progress following the ‘revolution’. Ital. J. Zool. 2004, 71, 5–16. [Google Scholar] [CrossRef]
- Toro Ibacache, M.V.; Manriquez Soto, G.; Suazo Galdames, I. Morfometría geométrica y el estudio de las formas biológicas: De la morfología descriptiva a la morfología cuantitativa. Int. J. Morphol. 2010, 28, 977–990. [Google Scholar] [CrossRef]
- Villalobos-Leiva, A.; Benítez, H.A. Morfometría geométrica y sus nuevas aplicaciones en ecología y biología evolutiva. Parte 2. Int. J. Morphol. 2020, 38, 1818–1836. [Google Scholar] [CrossRef]
- Bookstein, F.L. Morphometric Tools for Landmark Data; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Gould, S.J. Allometry and size in ontogeny and phylogeny. Biol. Rev. 1966, 41, 587–638. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Multivariate allometry. In Advances in Morphometrics; Marcus, L.F., Loy, M.C.A., Naylor, G.J.P., Slice, D.E., Eds.; Plenum Press: New York, NY, USA, 1996; pp. 23–49. [Google Scholar]
- Voje, K.L.; Hansen, T.F.; Egset, C.K.; Bolstad, G.H.; Pélabon, C. Allometric constraints and the evolution of allometry. Evolution 2014, 68, 866–885. [Google Scholar] [CrossRef]
- Pélabon, C.; Firmat, C.; Bolstad, G.H.; Voje, K.L.; Houle, D.; Cassara, J.; Rouzic, A.L.; Hansen, T.F. Evolution of morphological allometry. Ann. N. Y. Acad. Sci. 2014, 1320, 58–75. [Google Scholar] [CrossRef]
- Lemic, D.; Benítez, H.A.; Bažok, R. Intercontinental effect on sexual shape dimorphism and allometric relationships in the beetle pest Diabrotica virgifera virgifera LeConte (Coleoptera: Chrysomelidae). Zool. Anz. A J. Comp. Zool. 2014, 253, 203–206. [Google Scholar] [CrossRef]
- Göttig, S. Development of Eco-Friendly Methods for Monitoring and Regulating the Box Tree Pyralid, Cydalima perspectalis (Lepidoptera: Crambidae), an Invasive Pest in Ornamentals. Ph.D. Thesis, Technische Universität, Darmstadt, Germany, 2017. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, F.J.; Slice, D. Extensions of the Procrustes Method for the Optimal Superimposition of Landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef]
- Monteiro, L.R. Multivariate Regression Models and Geometric Morphometrics: The Search for Causal Factors in the Analysis of Shape. Syst. Biol. 1999, 48, 192–199. [Google Scholar] [CrossRef]
- Mitrovski Bogdanović, A.S. Molekularna i Morfološka Karakterizacija Vrsta iz Kompleksa Praon Dorsale-Yomenae (Hymenoptera, Braconidae). Ph.D. Thesis, University of Belgrade, Beograd, Serbia, 2012. [Google Scholar]
- Benítez, H.A.; Püschel, T.A.; Suazo, M.J. Drosophila Wing Integration and Modularity: A Multi-Level Approach to Understand the History of Morphological Structures. Biology 2022, 11, 567. [Google Scholar] [CrossRef] [PubMed]
- Bouyer, J.; Ravel, S.; Dujardin, J.-P.; De Meeüs, T.; Vial, L.; Thévenon, S.; Guerrini, L.; Sidibé, I.; Solano, P. Population structuring of Glossina palpalis gambiensis (Diptera: Glossinidae) according to landscape fragmentation in the Mouhoun river, Burkina Faso. J. Med. Entomol. 2007, 44, 788–795. [Google Scholar] [CrossRef]
- Lemic, D.; Mikac, K.M.; Kozina, A.; Benitez, H.A.; McLean, C.M.; Bažok, R. Monitoring techniques of the western corn rootworm are the precursor to effective IPM strategies. Pest Manag. Sci. 2016, 72, 405–417. [Google Scholar] [CrossRef]
- Kadoić Balaško, M.; Mikac, K.M.; Benítez, H.A.; Bažok, R.; Lemic, D. Genetic and Morphological Approach for Western Corn Rootworm Resistance Management. Agriculture 2021, 11, 585. [Google Scholar] [CrossRef]
- Kadoić Balaško, M.; Bažok, R.; Mikac, K.M.; Benítez, H.A.; Correa, M.; Lemic, D. Assessing the Population Structure of Colorado Potato Beetle Populations in Croatia Using Genetic and Geometric Morphometric Tools. Agronomy 2022, 12, 2361. [Google Scholar] [CrossRef]
- Andersson, M. Sexual Selection; Princeton University Press: Princeton, NJ, USA, 1994. [Google Scholar]
- Rodríguez-Muñoz, R.; Bretman, A.; Slate, J.; Walling, C.; Tregenza, T. Natural and Sexual Selection in a Wild Insect Population. Science 2010, 328, 1269–1272. [Google Scholar] [CrossRef]
- Stillwell, R.C.; Blanckenhorn, W.U.; Teder, T.; Davidowitz, G.; Fox, C.W. Sex Differences in Phenotypic Plasticity Affect Variation in Sexual Size Dimorphism in Insects: From Physiology to Evolution. Annu. Rev. Èntomol. 2010, 55, 227–245. [Google Scholar] [CrossRef]
- Benítez, H.A.; Vidal, M.; Briones, R.; Jerez, V. Sexual dimorphism and morphological variation in populations of Ceroglossus chilensis (Eschscholtz, 1829) (Coleoptera, Carabidae). J. Entomol. Res. Soc. 2010, 12, 87–95. [Google Scholar]
- Allen, C.E.; Zwaan, B.J.; Brakefield, P.M. Evolution of Sexual Dimorphism in the Lepidoptera. Annu. Rev. Èntomol. 2011, 56, 445–464. [Google Scholar] [CrossRef]
- Benítez, H.A.; Sukhodolskaya, R.A.; Órdenes-Clavería, R.; Avtaeva, T.A.; Kushalieva, S.A.; Saveliev, A.A. Measuring the Inter and Intraspecific Sexual Shape Dimorphism and Body Shape Variation in Generalist Ground Beetles in Russia. Insects 2020, 11, 361. [Google Scholar] [CrossRef]
- Hernández, -L.N.; Barragán, A.; Dupas, S.; Silvain, J.-F.; Dangles, O. Wing shape variations in an invasive moth are related to sexual dimorphism and altitude. Bull. Èntomol. Res. 2010, 100, 529–541. [Google Scholar] [CrossRef]
- Gidaszewski, N.A.; Baylac, M.; Klingenberg, C.P. Evolution of sexual dimorphism of wing shape in the Drosophila melanogaster subgroup. BMC Evol. Biol. 2009, 9, 110–111. [Google Scholar] [CrossRef]
- Pretorius, E. Using geometric morphometrics to investigate wing dimorphism in males and females of Hymenoptera—A case study based on the genus Tachysphex Kohl (Hymenoptera: Sphecidae: Larrinae). Aust. J. Èntomol. 2005, 44, 113–121. [Google Scholar] [CrossRef]
- Clarke, G.M. Developmental Stability and Fitness: The Evidence Is Not Quite So Clear. Am. Nat. 1998, 152, 762–766. [Google Scholar] [CrossRef]
- Wang, M.-Q.; Zhagn, H.-R.; Xi, Y.-Q.; Wang, G.-P.; Man, Z.; Zhang, L.-J.; Guo, X.-R. Population genetic variation and historical dynamics of the natural enemy insect Propylea japonica (Coleoptera: Coccinellidae) in China. J. Integr. Agric. 2022, 22, 2456–2469. [Google Scholar] [CrossRef]
- Dehnen-Schmutz, K.; Touza, J.; Perrings, C.; Williamson, M. A century of the ornamental plant trade and its impact on invasion success. Divers. Distrib. 2007, 13, 527–534. [Google Scholar] [CrossRef]
- Haeuser, E.; Dawson, W.; Thuiller, W.; Dullinger, S.; Block, S.; Bossdorf, O.; Carboni, M.; Conti, L.; Dullinger, I.; Essl, F.; et al. European ornamental garden flora as an invasion debt under climate change. J. Appl. Ecol. 2018, 55, 2386–2395. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Population | n | Females | Males | |
|---|---|---|---|---|---|
| Hand Collection | Hand Collection | Pheromone Trap | |||
| Terrestrial | Sveti Ivan Zelina | 20 | 20 | ||
| Zagreb | 78 | 22 | 56 | ||
| Garešnica | 47 | 28 | 19 | ||
| Coastal | Seget | 10 | 5 | 5 | |
| Split | 19 | 18 | 1 | ||
| Sinj | 33 | 17 | 16 | ||
| Kaštela | 62 | 41 | 21 | ||
| Locality/p-value | Sveti Ivan Zelina | Seget | Garešnica | Zagreb | Kaštela | Sinj |
|---|---|---|---|---|---|---|
| Seget | 2.168 0.0471 | |||||
| Garešnica | 2.7558 <0.0001 | 3.7601 <0.0001 | ||||
| Zagreb | 1.3877 0.6057 | 1.8757 0.0727 | 3.0898 <0.0001 | |||
| Kaštela | 2.5234 0.0015 | 3.534 <0.0001 | 2.8234 <0.0001 | 2.8728 <0.0001 | ||
| Sinj | 1.7339 0.0502 | 1.4956 0.2306 | 3.7198 <0.0001 | 1.3753 0.0922 | 3.264 <0.0001 | |
| Split | 2.2419 0.0436 | 2.9868 0.0001 | 3.4462 <0.0001 | 2.4283 <0.0001 | 2.6359 <0.0001 | 2.705 <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemic, D.; Viric Gasparic, H.; Majcenic, P.; Pajač Živković, I.; Bjeliš, M.; Suazo, M.J.; Correa, M.; Hernández, J.; Benítez, H.A. Wing Shape Variation between Terrestrial and Coastal Populations of the Invasive Box Tree Moth, Cydalima perspectalis, in Croatia. Animals 2023, 13, 3044. https://doi.org/10.3390/ani13193044
Lemic D, Viric Gasparic H, Majcenic P, Pajač Živković I, Bjeliš M, Suazo MJ, Correa M, Hernández J, Benítez HA. Wing Shape Variation between Terrestrial and Coastal Populations of the Invasive Box Tree Moth, Cydalima perspectalis, in Croatia. Animals. 2023; 13(19):3044. https://doi.org/10.3390/ani13193044
Chicago/Turabian StyleLemic, Darija, Helena Viric Gasparic, Patricija Majcenic, Ivana Pajač Živković, Mario Bjeliš, Manuel J. Suazo, Margarita Correa, Jordan Hernández, and Hugo A. Benítez. 2023. "Wing Shape Variation between Terrestrial and Coastal Populations of the Invasive Box Tree Moth, Cydalima perspectalis, in Croatia" Animals 13, no. 19: 3044. https://doi.org/10.3390/ani13193044
APA StyleLemic, D., Viric Gasparic, H., Majcenic, P., Pajač Živković, I., Bjeliš, M., Suazo, M. J., Correa, M., Hernández, J., & Benítez, H. A. (2023). Wing Shape Variation between Terrestrial and Coastal Populations of the Invasive Box Tree Moth, Cydalima perspectalis, in Croatia. Animals, 13(19), 3044. https://doi.org/10.3390/ani13193044