
Prevalence and Diversity of Blood Parasites (Plasmodium, Leucocytozoon and Trypanosoma) in Backyard Chickens (Gallus gallus domesticus) Raised in Southern Thailand
 , ,
, ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Method
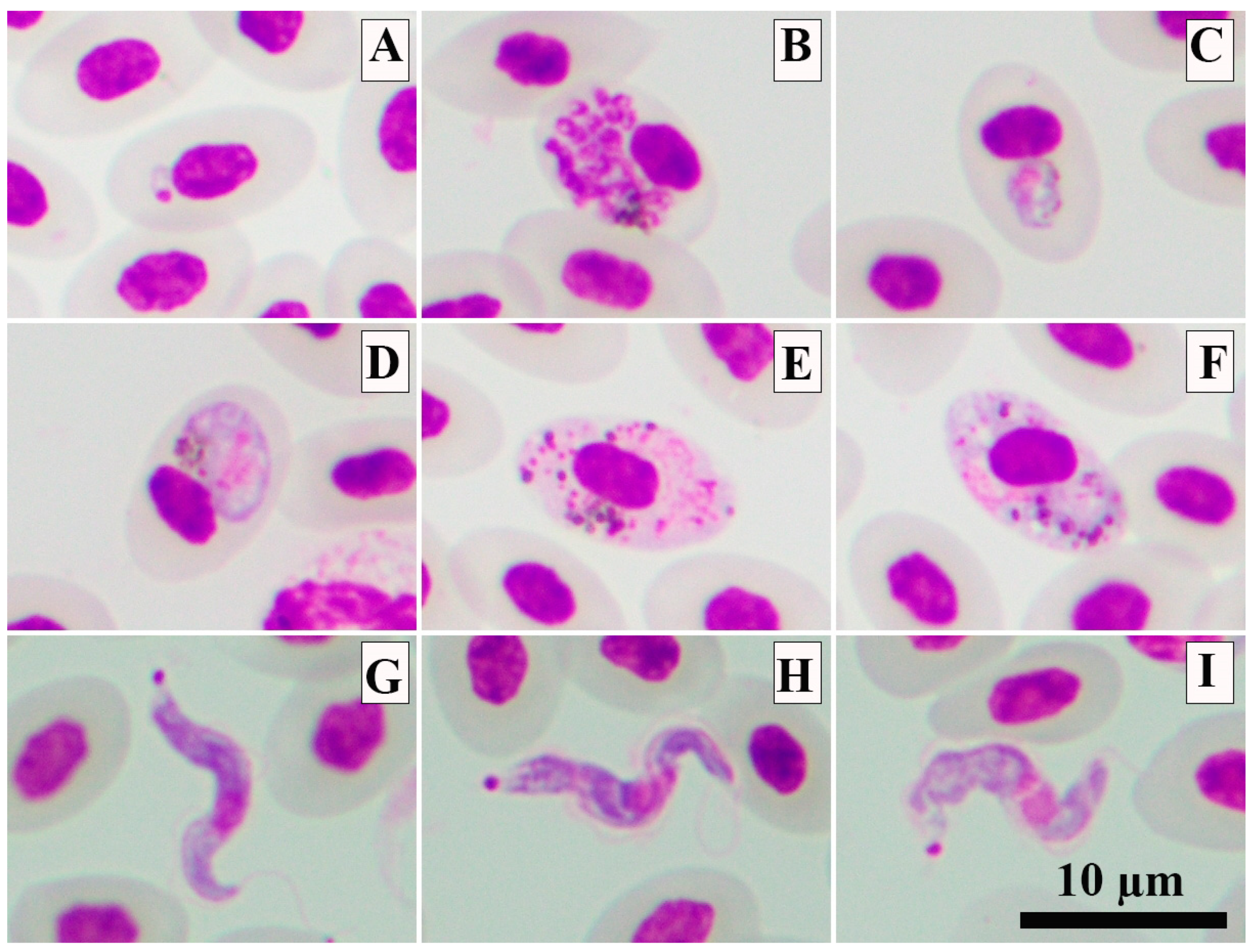
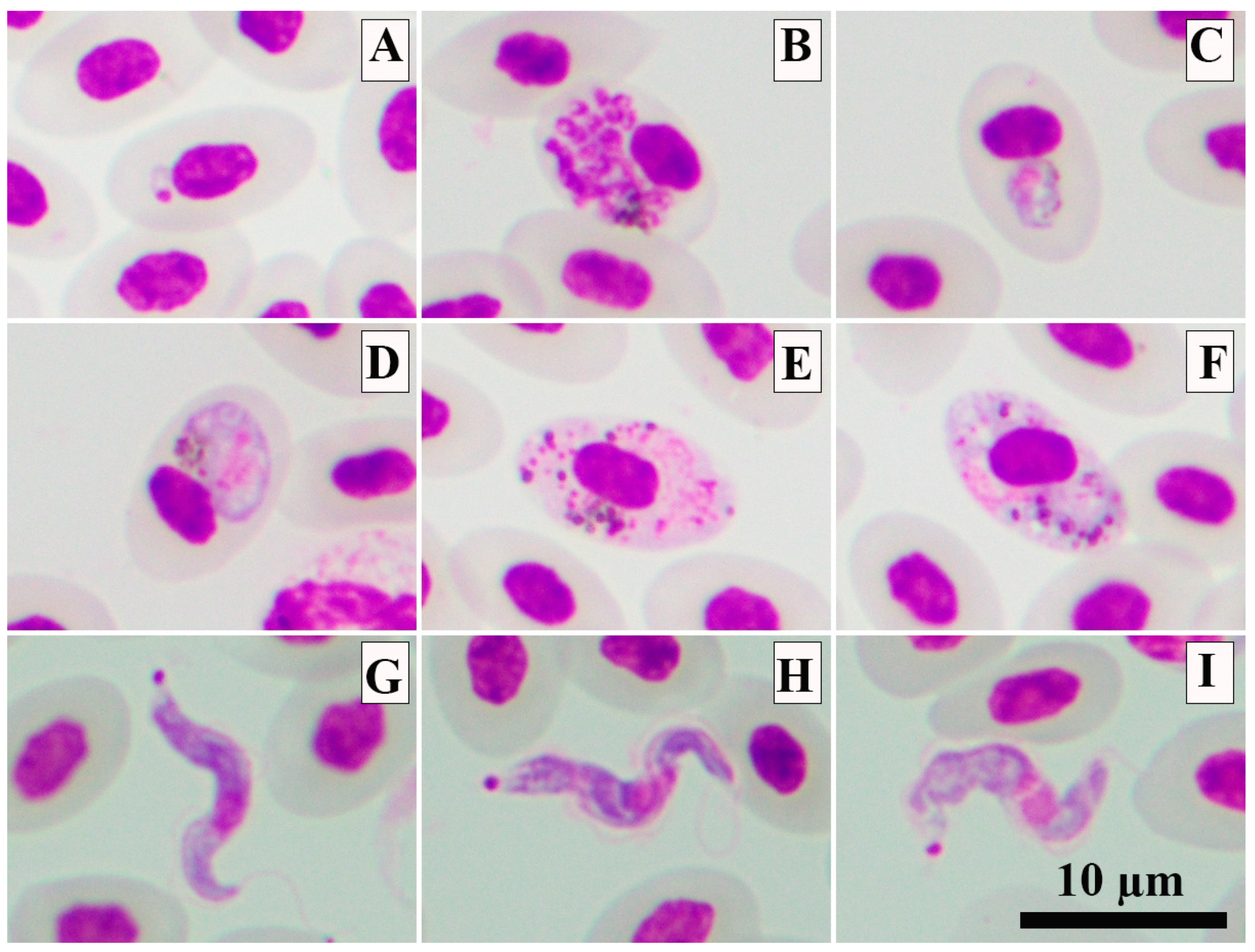
2.1. Sample Collection and Processing and Microscopic Examination
2.2. DNA Extraction, Nested-PCR and Sequencing
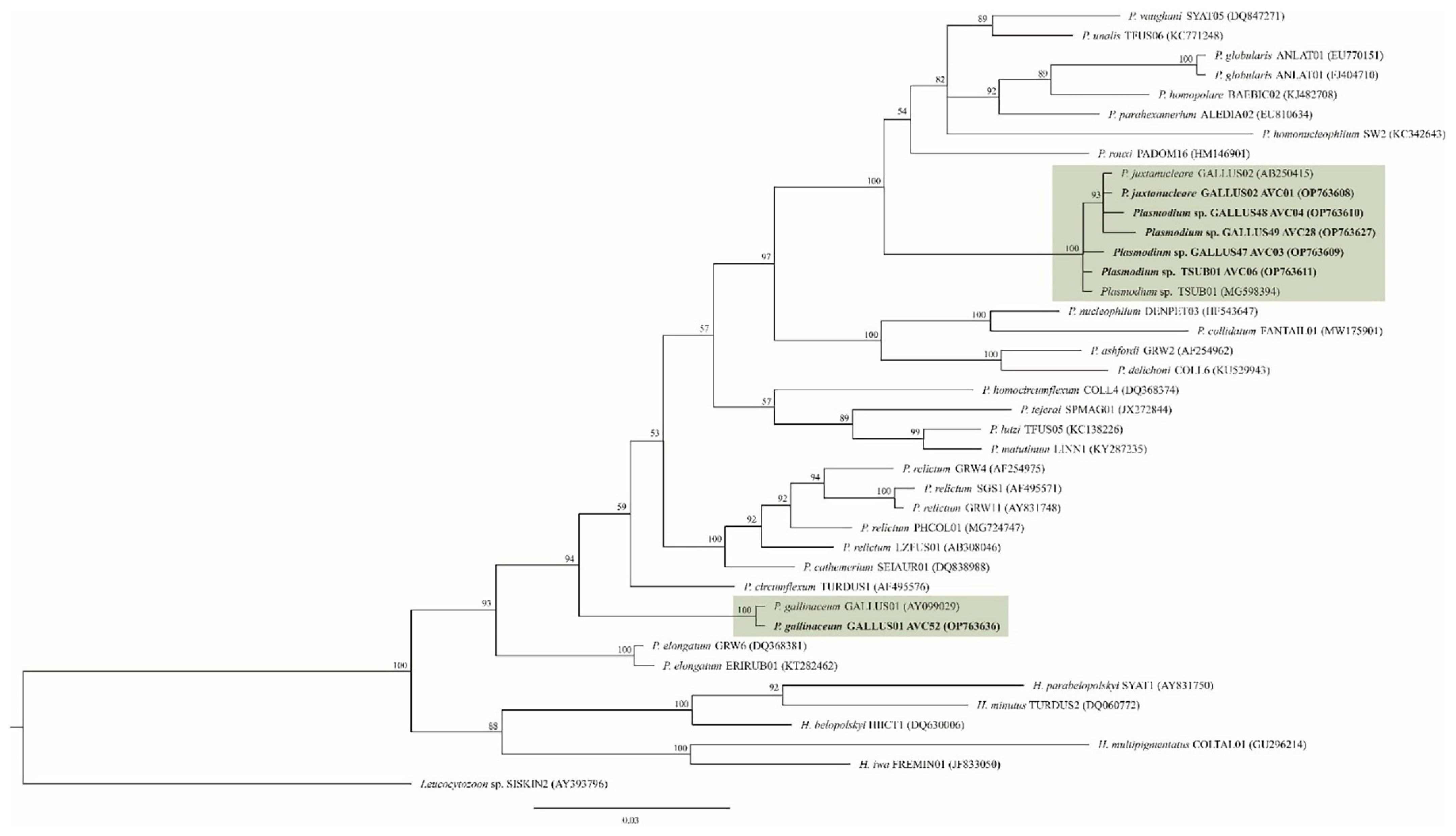
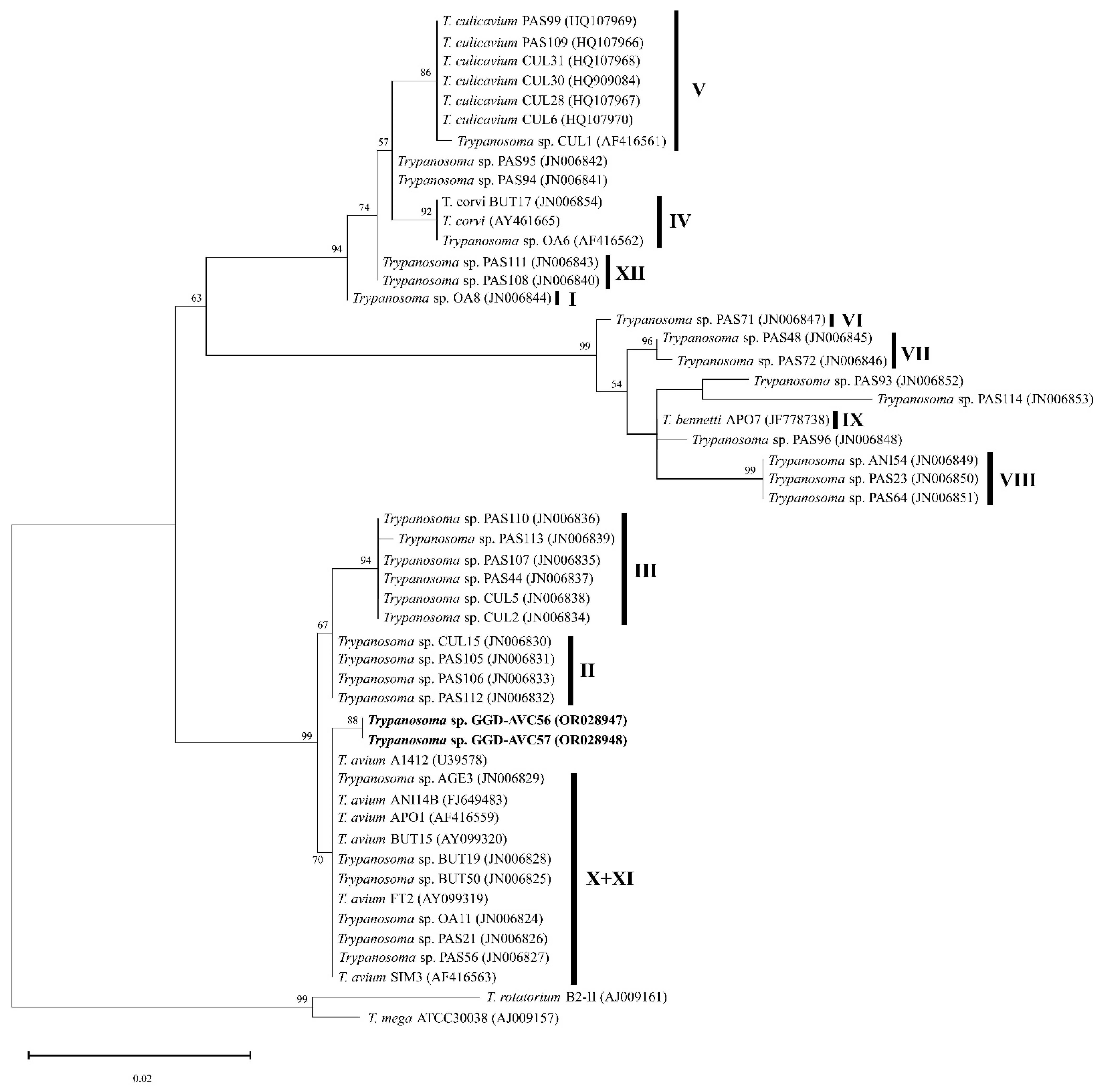
2.3. Sequence Analysis and Phylogenetics
2.4. Statistical Analysis
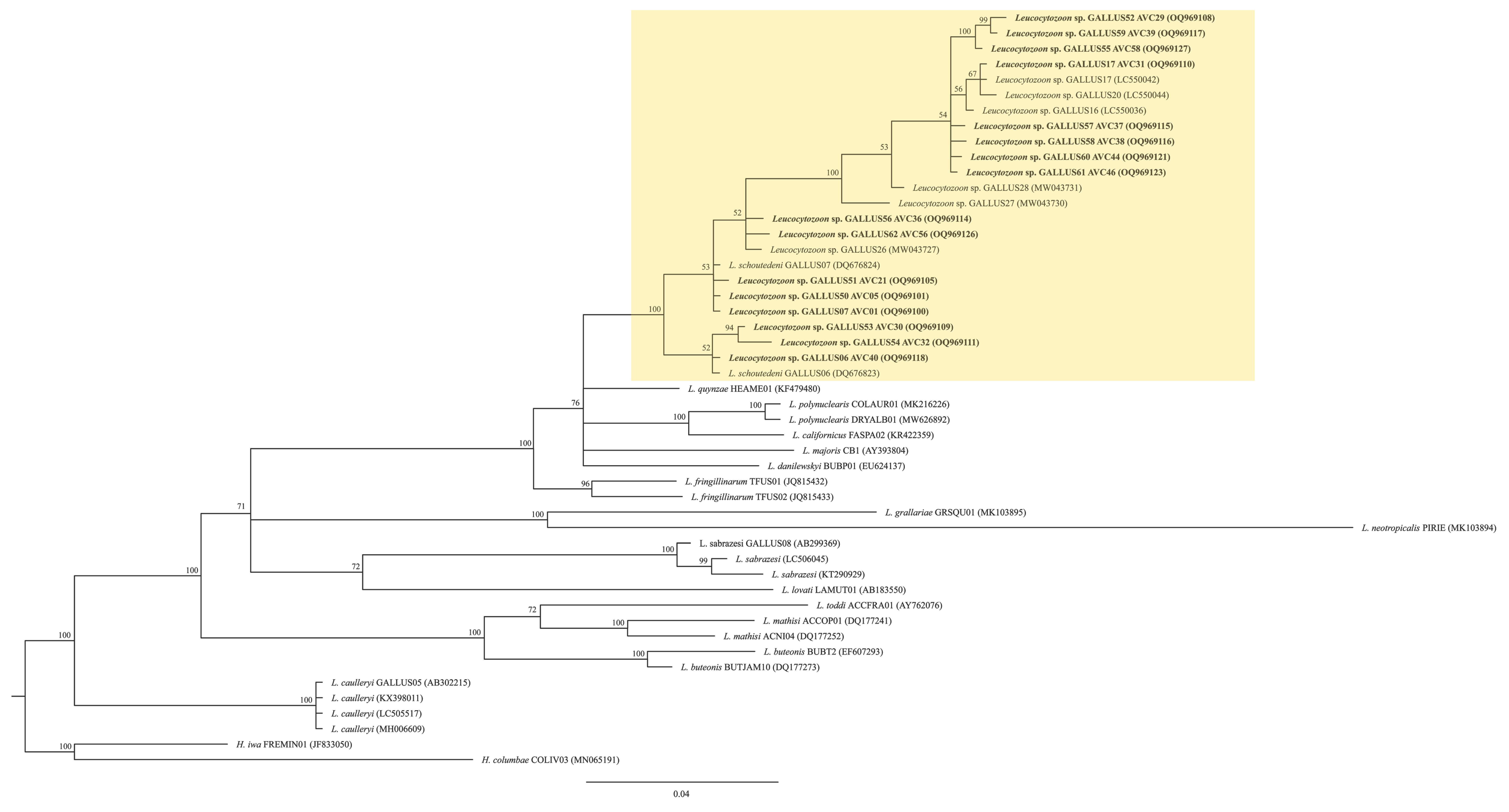
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Braga, E.M.; Silveira, P.; Belo, N.O.; Valkiūnas, G. Recent advances in the study of avian malaria: An overview with an emphasis on the distribution of Plasmodium spp in Brazil. Mem. Inst. Oswaldo. Cruz. 2011, 106, 3–11. [Google Scholar]
- Permin, A.; Juhl, J. The development of Plasmodium gallinaceum infections in chickens following single infections with three different dose levels. Vet. Parasitol. 2002, 105, 1–10. [Google Scholar] [PubMed]
- Valkiūnas, G.; Sehgal, R.N.M.; Iezhova, T.A.; Hull, A.C. Identification of Leucocytozoon toddi group (Haemosporida: Leucocytozoidae), with remarks on the species taxonomy of leucocytozoids. J. Pasasitol. 2010, 96, 170–177. [Google Scholar]
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC Press: Boca Roton, FL, USA, 2005. [Google Scholar]
- Valkiūnas, G.; Iezhova, T.A. Insights into the biology of Leucocytozoon Species (Haemosporida, Leucocytozoidae): Why is there slow research progress on agents of leucocytozoonosis? Microorganisms 2023, 11, 1251. [Google Scholar]
- Valkiūnas, G.; Iezhova, T.A. Keys to the avian malaria parasites. Malar. J. 2018, 17, 212. [Google Scholar] [PubMed]
- Valkiūnas, G.; Iezhova, T.A. Keys to the avian Haemoproteus parasites (Haemosporida, Haemoproteidae). Malar. J. 2022, 21, 269. [Google Scholar] [PubMed]
- Ferreira-Junior, F.; de Angeli Dutra, D.; Silveira, P.; Pacheco, R.; Witter, R.; de Souza Ramos, D.; Pacheco, M.A.; Escalante, A.; Braga, É. A new pathogen spillover from domestic to wild animals: Plasmodium juxtanucleare infects free-living passerines in Brazil. Parasitol 2016, 145, 1949–1958. [Google Scholar]
- Sehgal, R.N.M.; Valkiunas, G.; Iezhova, T.A.; Smith, T.B. Blood parasites of chickens in Uganda and Cameroon with molecular descriptions of Leucocytozoon schoutedeni and Trypanosoma gallinarum. J. Pasasitol. 2006, 92, 1336–1343. [Google Scholar]
- Pruck-Ngern, M.; Pattaradilokrat, S.; Chumpolbanchorn, K.; Pimnon, S.; Harnyuttanakorn, P.; Buddhirakkul, P.; Saiwichai, T. Refractoriness of the natural malaria vector Culex quinquefasciatus to Plasmodium gallinaceum. J. Trop. Med. Parasitol. 2014, 37, 60–68. [Google Scholar]
- Wiegmann, A.; Springer, A.; Rinaud, T.; Ottensmann, M.; Legler, M.; Krüger, O.; Fehr, M.; Chakarov, N.; Strube, C. The prevalence of Leucocytozoon spp. in nestlings of three wild raptor species including implications on haematological and blood chemistry values. Int. J. Parasitol. Parasites. Wild. 2021, 16, 236–243. [Google Scholar]
- Chawengkirttikul, R.; Junsiri, W.; Watthanadirek, A.; Poolsawat, N.; Minsakorn, S.; Srionrod, N.; Anuracpreeda, P. Molecular detection and genetic diversity of Leucocytozoon sabrazesi in chickens in Thailand. Sci. Rep. 2021, 11, 16686. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A.; Carlson, J.S.; Sehgal, R.N.M. Two new Trypanosoma species from African birds, with notes on the taxonomy of avian trypanosomes. J. Pasasitol. 2011, 97, 924–930. [Google Scholar] [CrossRef]
- Molyneux, D.H. Vector relationships in the Trypanosomatidae. Adv. Parasitol. 1977, 15, 1–82. [Google Scholar]
- Miltgen, F.; Landau, I. Culicoides nubeculosus, vecteur expérimental d’un nouveau trypanosome de psittaciforme: Trypanosoma bakeri n. sp. Ann. Parasitol. Hum. Comp 1982, 57, 423–428. [Google Scholar] [CrossRef]
- Votýpka, J.; Svobodová, M. Trypanosoma avium: Experimental transmission from black flies to canaries. Parasitol. Res. 2004, 92, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.B. Avian malaria: Clinical and chemical pathology of Plasmodium gallinaceum in the domesticated fowl Gallus gallus. Avian. Pathol. 2005, 34, 29–47. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.R.; Koo, B.-S.; Jeon, E.-O.; Han, M.-S.; Min, K.-C.; Lee, S.B.; Bae, Y.; Mo, I.-P. Pathology and molecular characterization of recent Leucocytozoon caulleryi cases in layer flocks. J. Biomed. Res. 2016, 30, 517–524. [Google Scholar] [PubMed]
- Prasopsom, P.; Salakij, C.; Lertwatcharasarakul, P.; Pornpranom, P. Hematological and phylogenetic studies of Leucocytozoon spp. in backyard chickens and fighting cocks around Kamphaeng Saen, Thailand. Agr. Nat. Resour. 2020, 54, 595–602. [Google Scholar]
- Desser, S.S.; Stuht, J.; Fallis, A.M. Leucocytozoonosis in Canada geese in upper Michigan,. I. Strain differences among geese from different localities. J. Wildl. Dis. 1978, 14, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Desser, S.S. Schizogony and gametogony of Leucocytozoon simondi and associated reactions in the avian host. J. Protozool. 1967, 14, 244–254. [Google Scholar] [CrossRef]
- Desser, S.S.; Ryckman, A.K. The development and pathogenesis of Leucocytozoon simondi in Canada and domestic geese in Algonquin Park, Ontario. Can. J. Zool. 1976, 54, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.A.; Fallis, A.M. Comparison of infections with Leucocytozoon simondi in black ducks (Anas rubripes), mallards (Anas platyrhynchos), and white Pekins (Anas bochas). Can. J. Zool. 1968, 46, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Win, S.Y.; Chel, H.M.; Hmoon, M.M.; Htun, L.L.; Bawm, S.; Win, M.M.; Murata, S.; Nonaka, N.; Nakao, R.; Katakura, K. Detection and molecular identification of Leucocytozoon and Plasmodium species from village chickens in different areas of Myanmar. Acta Trop. 2020, 212, 105719. [Google Scholar] [CrossRef] [PubMed]
- Pohuang, T.; Jittimanee, S.; Junnu, S. Pathology and molecular characterization of Leucocytozoon caulleryi from backyard chickens in Khon Kaen Province, Thailand. Vet. World 2021, 14, 2634–2639. [Google Scholar] [CrossRef] [PubMed]
- Šlapeta, J.; Morin-Adeline, V.; Thompson, P.; McDonell, D.; Shiels, M.; Gilchrist, K.; Votýpka, J.; Vogelnest, L. Intercontinental distribution of a new trypanosome species from Australian endemic Regent Honeyeater (Anthochaera phrygia). Parasitol 2016, 143, 1012–1025. [Google Scholar] [CrossRef] [PubMed]
- Zídková, L.; Cepicka, I.; Szabová, J.; Svobodová, M. Biodiversity of avian trypanosomes. Infect. Genet. Evol. 2012, 12, 102–112. [Google Scholar] [CrossRef]
- Votýpka, J.; Szabová, J.; Rádrová, J.; Zídková, L.; Svobodová, M. Trypanosoma culicavium sp. nov., an avian trypanosome transmitted by Culex mosquitoes. Int. J. Syst. Evol. Microbiol. 2012, 62, 745–754. [Google Scholar] [CrossRef]
- Pornpanom, P.; Salakij, C.; Prasopsom, P.; Lertwatcharasarakul, P.; Kasorndorkbua, C. Morphological and molecular characterization of avian trypanosomes in raptors from Thailand. Parasitol. Res. 2019, 118, 2419–2429. [Google Scholar] [CrossRef]
- Chantong, W.; Kaneene, J.B. Poultry raising systems and highly pathogenic avian influenza outbreaks in Thailand: The situation, associations, and impacts. Southeast Asian J. Trop. Med. Public Health 2011, 42, 596–608. [Google Scholar]
- Ayala, A.J.; Yabsley, M.J.; Hernandez, S.M. A review of pathogen transmission at the backyard chicken–wild bird interface. Front. Vet. Sci. 2020, 7, 539925. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.-A.; Krizˇanauskiene˙, A.; Palinauskas, V.; Sehgal, R.N.M.; Bensch, S. A comparative analysis of microscopy and PCR-based detection methods for blood parasites. J. Parasitol. 2008, 94, 1395–1401. [Google Scholar] [CrossRef]
- Chagas, C.R.F.; Binkienė, R.; Ilgūnas, M.; Iezhova, T.; Valkiūnas, G. The buffy coat method: A tool for detection of blood parasites without staining procedures. Parasit. Vectors 2020, 13, 104. [Google Scholar] [CrossRef]
- Godfrey, R.D.; Fedynich, A.M.; Pence, D.B. Quantification of hematozoa in blood smears. J. Wildl. Dis. 1987, 23, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Bensch, S.; Stjerman, M.; Hasselquist, D.; Östman, Ö.; Hansson, B.; Westerdahl, H.; Pinheiro, R.T. Host specificity in avian blood parasites: A study of Plasmodium and Haemoproteus mitochondrial DNA amplified from birds. Proc. Roy. Soc. Lond. B 2000, 267, 1583–1589. [Google Scholar] [CrossRef] [PubMed]
- Hellgren, O.; WaldenstrÖm, J.; Bensch, S. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avain blood. J. Pasasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Pornpanom, P.; Chagas, C.R.F.; Lertwatcharasarakul, P.; Kasorndorkbua, C.; Valkiūnas, G.; Salakij, C. Molecular prevalence and phylogenetic relationship of Haemoproteus and Plasmodium parasites of owls in Thailand: Data from a rehabilitation centre. Int. J. Parasitol. Parasites Wildl. 2019, 9, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Lertwatcharasarakul, P.; Salakij, C.; Prasopsom, P.; Kasorndorkbua, C.; Jakthong, P.; Santavakul, M.; Suwanasaeng, P.; Ploypan, R. Molecular and morphological analyses of Leucocytozoon parasites (Haemosporida: Leucocytozoidae) in raptors from Thailand. Acta Parasitol. 2021, 66, 1406–1416. [Google Scholar] [CrossRef]
- Pornpanom, P.; Kasorndocbau, C.; Lertwatcharasarakul, P.; Salakij, C. Prevalence and genetic diversity of Haemoproteus and Plasmodium in raptors from Thailand: Data from rehabilitation center. Int. J. Parasitol. Parasites Wildl. 2021, 16, 75–82. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Harl, J.; Himmel, T.; Valkiūnas, G.; Ilgūnas, M.; Nedorost, N.; Matt, J.; Kübber-Heiss, A.; Alic, A.; Konicek, C.; Weissenböck, H. Avian haemosporidian parasites of accipitriform raptors. Malar. J. 2022, 21, 14. [Google Scholar] [CrossRef]
- Cadena-Ortiz, H.; Mantilla, J.S.; de Aguilar, J.R.; Flores, D.; Bahamonde, D.; Matta, N.E.; Bonaccorso, E. Avian haemosporidian infections in rufous-collared sparrows in an Andean dry forest: Diversity and factors related to prevalence and parasitaemia. Parasitol 2019, 146, 765–773. [Google Scholar] [CrossRef]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef]
- Chagas, C.R.F.; Valkiūnas, G.; de Oliveira Guimarães, L.; Monteiro, E.F.; Vaz Guida, F.J.; Simões, R.F.; Rodrigues, P.T.; de Albuquerque Luna, E.J.; Karin Kirchgatter, K. Diversity and distribution of avian malaria and related haemosporidian parasites in captive birds from a Brazilian megalopolis. Malar. J. 2017, 16, 83. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, K.; Zehtindjiev, P.; Mariaux, J.; Georgiev, B.B. Genetic diversity of avian haemosporidians in Malaysia: Cytochrome b lineages of the genera Plasmodium and Haemoproteus (Haemosporida) from Selangor. Infect. Genet. Evol. 2015, 31, 33–39. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest 2.3. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Chumsri, A.; Tina, F.W.; Jaroensutasinee, M.; Jaroensutasinee, K. Seasons and water container types affecting Culex spp. in southern Thailand. J. Anim. Behav. Biometeorol. 2020, 8, 55–62. [Google Scholar] [CrossRef]
- Sunantaraporn, S.; Hortiwakul, T.; Kraivichian, K.; Siriyasatien, P.; Brownell, N. Molecular identification of host blood meals and detection of blood parasites in Culicoides Latreille (Diptera: Ceratopogonidae) collected from Phatthalung province, Southern Thailand. Insects 2022, 13, 912. [Google Scholar] [CrossRef]
- Srisuka, W.; Takaoka, H.; Otsuka, Y.; Fukuda, M.; Thongsahuan, S.; Taai, K.; Saeung, K. Biodiversity, seasonal abundance, and distribution of blackflies (Diptera: Simuliidae) in six different regions ofThailand. Parasit. Vectors 2017, 10, 574. [Google Scholar] [CrossRef]
- Muriel, J.; Marzal, A.; Magallanes, S.; García-Longoria, L.; Suarez-Rubio, M.; Bates, P.J.J.; Lin, H.H.; Soe, A.N.; Oo, K.S.; Aye, A.A.; et al. Prevalence and diversity of avian haemosporidians may vary with anthropogenic disturbance in tropical habitats in Myanmar. Diversity 2021, 13, 111. [Google Scholar] [CrossRef]
- Ferraguti, M.; Martínez-de la Puente, J.; Bensch, S.; Roiz, D.; Ruiz, S.; Viana, D.S.; Soriguer, R.C.; Figuerola, J. Ecological determinants of avian malaria infections: An integrative analysis at landscape, mosquito and vertebrate community levels. J. Anim. Ecol. 2018, 87, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Valkiūnas, G.; Bensch, S.; Iezhova, T.A.; Križanauskienė, A.; Hellgren, O.; Bolshakov, C.V. Nested cytochrome b polymerase chain reaction diagnostics underestimate mixed infections of avian blood haemosporidian parasites: Microscopy is still essential. J. Pasasitol. 2006, 92, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Zehtindjiev, P.; Križanauskienė, A.; Bensch, S.; Palinauskas, V.; Asghar, M.; Dimitrov, D.; Scebba, S.; Valkiūnas, G. A new morphologically distinct avian malaria parasite that fails detection by established polymerase chain reaction–based protocols for amplification of the cytochrome b gene. J. Pasasitol. 2012, 98, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Tris, J.; Bensch, S. Diagnosing genetically diverse avian malarial infections using mixed-sequence analysis and TA-cloning. Parasitol 2005, 131, 15–23. [Google Scholar] [CrossRef] [PubMed]
- LaPointe, D.A.; Goff, M.L.; Atkinson, C.T. Thermal constraints to the sporogonic development and altitudinal dstribution of avian malaria Plasmodium relictum in Hawaii. J. Parasitol. 2010, 96, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Chaiphongpachara, T.; Sumruayphol, S. Species diversity and distribution of mosquito vectors in coastal habitats of Samut Songkhram province, Thailand. Trop. Biomed. 2017, 34, 524–532. [Google Scholar]
- Perkins, S.L.; Schall, J.J. A molecular phylogeny of malarial parasites recovered from cytochrome b gene sequences. J. Pasasitol. 2002, 88, 972–978. [Google Scholar] [CrossRef]
- Pattaradilokrat, S.; Tiyamaneea, W.; Simpalipana, P.; Kaewthamasornb, M.; Saiwichai, T.; Li, J.; Harnyuttanakorna, P. Molecular detection of the avian malaria parasite Plasmodium gallinaceum in Thailand. Vet. Parasitol. 2015, 210, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Matis, C.; Léger, M. Trypanosome de la poule. Comptes Rendus Des Se’Ances Socie´Te´ Biol. Ses Fil. 1909, 67, 452–454. [Google Scholar]
- Fallis, A.M.; Jacobson, R.L.; Raybould, J.N. Experimental transmission of Trypanosoma numidae Wenyon to guinea fowl and chickens in Tanzania. J. Protozool. 1973, 20, 436–437. [Google Scholar] [CrossRef]
- Cendron, F.; Perini, F.; Mastrangelo, S.; Tolone, M.; Criscione, A.; Bordonaro, S.; Iaffaldano, N.; Castellini, C.; Marzoni, M.; Buccioni, A.; et al. Genome-wide SNP analysis reveals the population structure and the conservation status of 23 Italian chicken breeds. Animals 2020, 10, 1441. [Google Scholar] [CrossRef] [PubMed]
- Kaya, S.; Kabasakal, B.; Erdoğan, A. Geographic genetic structure of Alectoris chukar in Türkiye: Post-LGM-induced hybridization and human-mediated contaminations. Biology 2023, 12, 401. [Google Scholar] [CrossRef] [PubMed]
- Anjos, C.C.; Chagas, C.R.F.; Fecchio, A.; Schunck, F.; Costa-Nascimento, M.J.; Monteiro, E.F.; Mathias, B.S.; Bell, J.F.; Guimarães, L.O.; Comiche, K.J.M.; et al. Avian malaria and related parasites from resident and migratory birds in the brazilian atlantic forest, with description of a new Haemoproteus species. Pathogens 2021, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Pereira, I.T.; Hargrove, G.H.; Cornbleet, P.J. Preparation of buffy coat smears in leukopenic patients. Lab. Med. 1981, 12, 96–98. [Google Scholar] [CrossRef]
- Marcos, R.; Pereira, C.; Santos, M.; Luzzago, C.; Lauzi, S.; Maia, J.P.; Faustino, A.; Puente-Payo, P. Buffy coat smear or Knott’s test: Which to choose for canine microfilaria screening in field studies? Vet. Clin. Pathol. 2016, 45, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Salakij, C.; Kasorndorkbua, C.; Lertwatcharasarakul, P.; Salakij, J. Ultra-structure of blood cells and molecular characteristics of Haemoproteus sp. in Blyth’s hawk eagle. Comp. Clin. Pathol. 2015, 24, 1293–1299. [Google Scholar] [CrossRef]
- Setiawan, A.; Nurcahyo, W.; Priyowidodo, D.; Budiati, R.T.; Susanti, D.S.R. Genetic and parasitological identification of Trypanosoma evansi infecting cattle in South Sulawesi, Indonesia. Vet. World 2021, 14, 113–119. [Google Scholar] [CrossRef]
- Adeoye, G.O.; Nga, I.C. Comparison of quantitative buffy coat technique (QBC) with Giemsa-stained thick film (GTF) for diagnosis of malaria. Parasitol. Int. 2007, 56, 308–312. [Google Scholar] [CrossRef]
- Kocharekar, M.M.; Sarkar, S.S.; Dasgupta, D. Comparative study of modified quantitative buffy coat and two rapid tests in comparison with peripheral blood smear in malaria diagnosis in Mumbai, India. J. Parasitol. Res. 2014, 2014, 194651. [Google Scholar] [CrossRef]
- Ifeorah, I.K.; Brown, B.J.; Sodeinde, O.O. A comparison of rapid diagnostic testing (by Plasmodium lactate dehydrogenase), and quantitative buffy coat technique in malaria diagnosis in children. Afr. J. Infect. Dis. 2017, 11, 31–38. [Google Scholar]
- Charpentier, E.; Benichou, E.; Pagès, A.; Chauvin, P.; Fillaux, J.; Valentin, A.; Guegan, H.; Guemas, E.; Salabert, A.-S.; Armengol, C.; et al. Performance evaluation of different strategies based on microscopy techniques, rapid diagnostic test and molecular loop-mediated isothermal amplification assay for the diagnosis of imported malaria. Clin. Microbiol. Infect. 2020, 26, 115–121. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Buffy Coat Smear (n = 55) | Nested-PCR (n = 57) | Prevalence | |
|---|---|---|---|
| Plasmodium sp. | 5 | 37 a | 64.91% |
| Leucocytozoon sp. | 48 | 51 b | 89.47% |
| Trypanosoma sp. | 4 | 19 c | 33.33% |
| Microfilaria | 15 | NA | NA |
| Linages | Isolates | Parasite Species | Host Breed | Locality | GenBank | |
|---|---|---|---|---|---|---|
| District | Province | |||||
| GALLUS01 | AVC16 | Plasmodium gallinaceum | N | LAS | NST | OP763632 |
| AVC32 | N | KOR | PHL | OP763633 | ||
| AVC39 | N | KOR | PHL | OP763634 | ||
| AVC46 | N | THL | NST | OP763635 | ||
| AVC52 | N | KHK | PHL | OP763636 | ||
| AVC56 | N | KHK | PHL | OP763637 | ||
| AVC58 | N | KHK | PHL | OP763638 | ||
| GALLUS02 | AVC01 | Plasmodium juxtanucleare | H | THL | NST | OP763608 |
| AVC08 | H | THL | NST | OP763613 | ||
| AVC09 | H | THL | NST | OP763614 | ||
| AVC13 | LH | THL | NST | OP763616 | ||
| AVC18 | N | LAS | NST | OP763620 | ||
| TSUB01 | AVC06 | Plasmodium sp. | H | THL | NST | OP763611 |
| AVC07 | H | THL | NST | OP763612 | ||
| AVC11 | LH | THL | NST | OP763615 | ||
| AVC14 | LH | THL | NST | OP763617 | ||
| AVC15 | LH | THL | NST | OP763618 | ||
| AVC17 | N | LAS | NST | OP763619 | ||
| AVC20 | N | LAS | NST | OP763621 | ||
| AVC22 | FR | PHS | SUT | OP763622 | ||
| AVC23 | FR | PHS | SUT | OP763623 | ||
| AVC24 | FR | PHS | SUT | OP763624 | ||
| AVC25 | FR | PHS | SUT | OP763625 | ||
| AVC26 | FR | BND | SUT | OP763626 | ||
| AVC29 | FR | BND | SUT | OP763628 | ||
| AVC31 | N | KOR | PHL | OP763629 | ||
| AVC51 | N | KHK | PHL | OP763630 | ||
| AVC57 | N | KHK | PHL | OP763631 | ||
| GALLUS47 | AVC03 | Plasmodium sp. | H | THL | NST | OP763609 |
| GALLUS48 | AVC04 | Plasmodium sp. | H | THL | NST | OP763610 |
| GALLUS49 | AVC28 | Plasmodium sp. | FR | BND | SUT | OP763627 |
| GALLUS06 | AVC40 | Leucocytozoon schoutedeni | N | KOR | PHL | OQ969118 |
| GALLUS07 | AVC01 | Leucocytozoon schoutedeni | H | THL | NST | OQ969100 |
| AVC07 | H | THL | NST | OQ969102 | ||
| AVC22 | FR | PHS | SUT | OQ969106 | ||
| AVC34 | N | KOR | PHL | OQ969112 | ||
| AVC43 | N | KOR | PHL | OQ969120 | ||
| GALLUS17 | AVC31 | Leucocytozoon sp. | N | KOR | PHL | OQ969110 |
| AVC41 | N | KOR | PHL | OQ969119 | ||
| AVC50 | N | KHK | PHL | OQ969125 | ||
| GALLUS50 | AVC05 | Leucocytozoon sp. | H | THL | NST | OQ969101 |
| AVC09 | H | THL | NST | OQ969103 | ||
| AVC18 | N | LAS | NST | OQ969104 | ||
| GALLUS51 | AVC21 | Leucocytozoon sp. | FR | PHS | SUT | OQ969105 |
| AVC23 | FR | PHS | SUT | OQ969107 | ||
| GALLUS52 | AVC29 | Leucocytozoon sp. | FR | BND | SUT | OQ969108 |
| AVC45 | N | KOR | PHL | OQ969122 | ||
| GALLUS53 | AVC30 | Leucocytozoon sp. | FR | BND | SUT | OQ969109 |
| GALLUS54 | AVC32 | Leucocytozoon sp. | N | KOR | PHL | OQ969111 |
| GALLUS55 | AVC35 | Leucocytozoon sp. | N | KOR | PHL | OQ969113 |
| AVC58 | N | KHK | PHL | OQ969127 | ||
| GALLUS56 | AVC36 | Leucocytozoon sp. | N | KOR | PHL | OQ969114 |
| GALLUS57 | AVC37 | Leucocytozoon sp. | N | KOR | PHL | OQ969115 |
| GALLUS58 | AVC38 | Leucocytozoon sp. | N | KOR | PHL | OQ969116 |
| GALLUS59 | AVC39 | Leucocytozoon sp. | N | KOR | PHL | OQ969117 |
| GALLUS60 | AVC44 | Leucocytozoon sp. | N | KOR | PHL | OQ969121 |
| GALLUS61 | AVC46 | Leucocytozoon sp. | N | THL | NST | OQ969123 |
| GALLUS62 | AVC56 | Leucocytozoon sp. | N | KHK | PHL | OQ969126 |
| Parameter * | Trypanosoma sp. (n = 5) | ||||
|---|---|---|---|---|---|
| Mean | SD | Min | Max | ||
| AK | (µm2) | 0.68 | 0.06 | 0.61 | 0.75 |
| AN | (µm2) | 4.45 | 0.79 | 3.62 | 5.63 |
| AT | (µm2) | 31.68 | 3.79 | 27.71 | 36.72 |
| BW | (µm) | 2.22 | 0.24 | 2.01 | 2.60 |
| FF | (µm) | 8.01 | 0.84 | 6.59 | 8.67 |
| KN | (µm) | 8.70 | 0.82 | 7.35 | 9.53 |
| NA | (µm) | 7.51 | 1.15 | 6.01 | 9.15 |
| PA | (µm) | 18.04 | 1.55 | 15.81 | 20.12 |
| PK | (µm) | 0.91 | 0.22 | 0.76 | 1.29 |
| PN | (µm) | 10.61 | 0.64 | 9.78 | 11.56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boonchuay, K.; Thomrongsuwannakij, T.; Chagas, C.R.F.; Pornpanom, P. Prevalence and Diversity of Blood Parasites (Plasmodium, Leucocytozoon and Trypanosoma) in Backyard Chickens (Gallus gallus domesticus) Raised in Southern Thailand. Animals 2023, 13, 2798. https://doi.org/10.3390/ani13172798
Boonchuay K, Thomrongsuwannakij T, Chagas CRF, Pornpanom P. Prevalence and Diversity of Blood Parasites (Plasmodium, Leucocytozoon and Trypanosoma) in Backyard Chickens (Gallus gallus domesticus) Raised in Southern Thailand. Animals. 2023; 13(17):2798. https://doi.org/10.3390/ani13172798
Chicago/Turabian StyleBoonchuay, Kanpapat, Thotsapol Thomrongsuwannakij, Carolina Romeiro Fernandes Chagas, and Pornchai Pornpanom. 2023. "Prevalence and Diversity of Blood Parasites (Plasmodium, Leucocytozoon and Trypanosoma) in Backyard Chickens (Gallus gallus domesticus) Raised in Southern Thailand" Animals 13, no. 17: 2798. https://doi.org/10.3390/ani13172798
APA StyleBoonchuay, K., Thomrongsuwannakij, T., Chagas, C. R. F., & Pornpanom, P. (2023). Prevalence and Diversity of Blood Parasites (Plasmodium, Leucocytozoon and Trypanosoma) in Backyard Chickens (Gallus gallus domesticus) Raised in Southern Thailand. Animals, 13(17), 2798. https://doi.org/10.3390/ani13172798