

Matching an Old Marine Paradigm: Limitless Connectivity in a Deep-Water Fish over a Large Distance

 ,
,  , ,
, ,  , , , ,
, , , ,
Abstract
Simple Summary
Abstract
1. Introduction
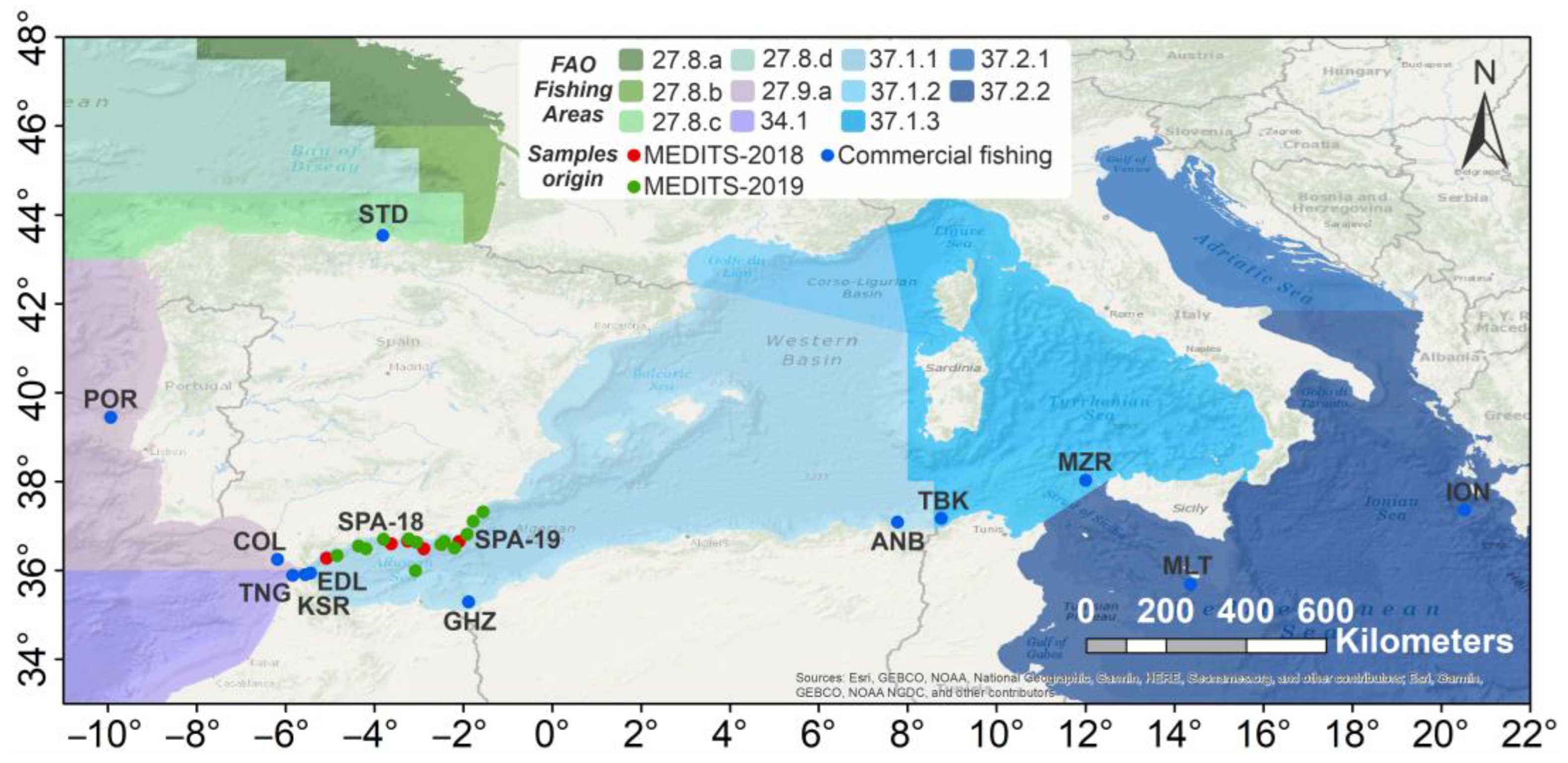
2. Materials and Methods
2.1. Sampling
2.2. Genetic Data Analysis
2.3. Data Analysis
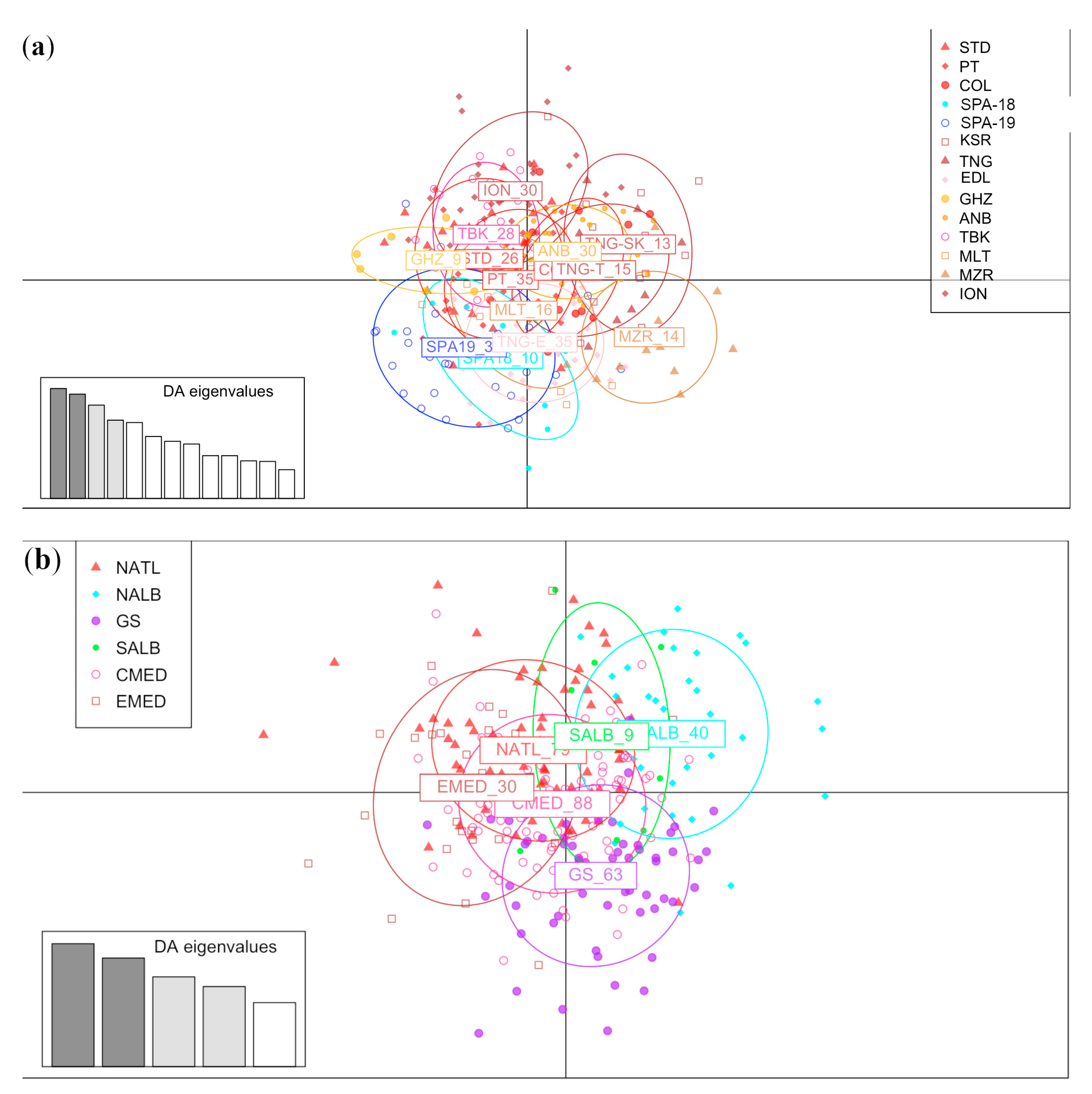
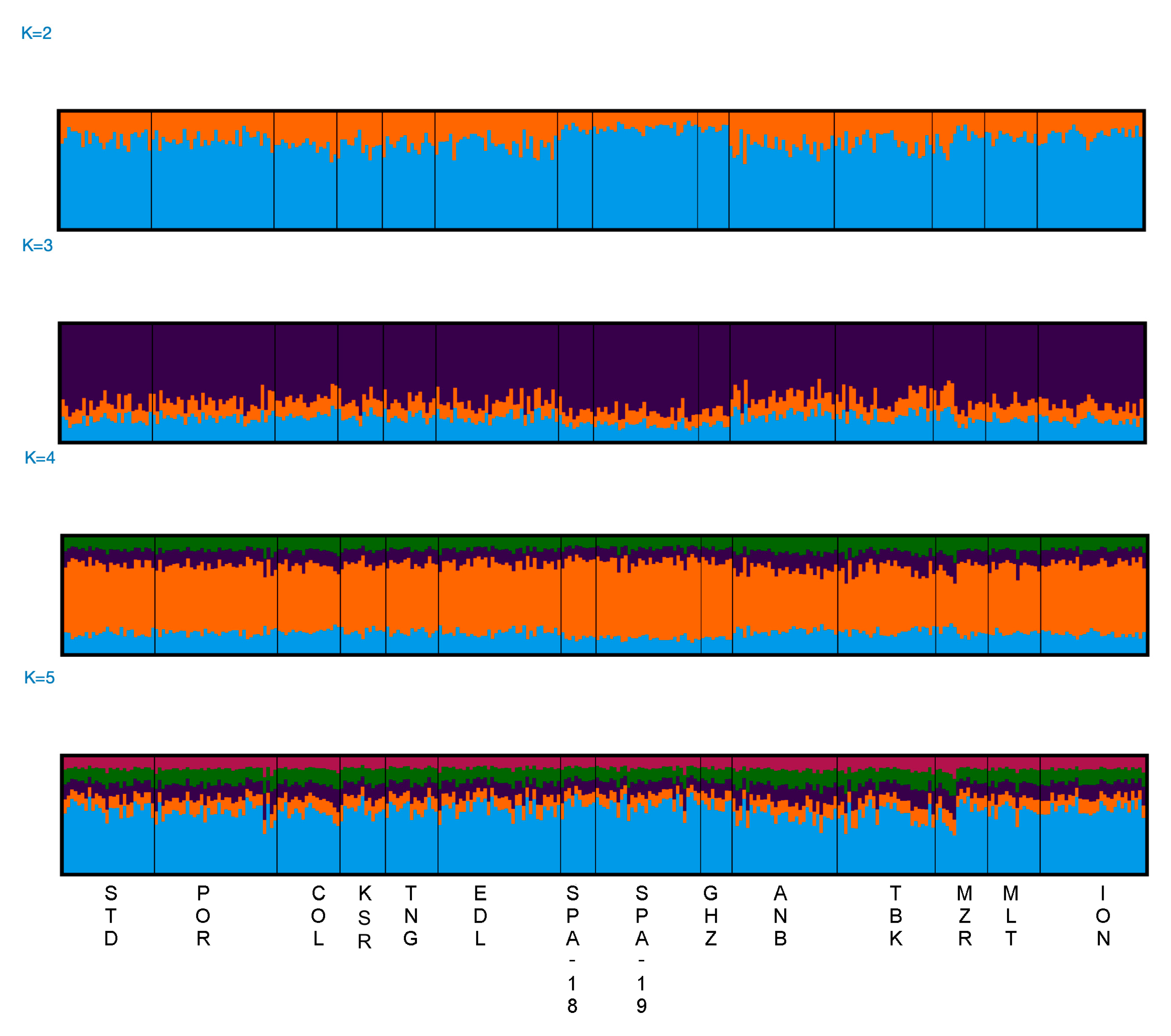
3. Results
3.1. Genetic Diversity
3.2. Population Differentiation and Demography
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palumbi, S.R. Genetic divergence, reproductive isolation, and marine speciation. Annu. Rev. Ecol. Syst. 1994, 25, 547–572. [Google Scholar] [CrossRef]
- Palumbi, S.R. Molecular biogeography of the Pacific. Coral Reefs 1997, 16, S47–S52. [Google Scholar] [CrossRef]
- Palumbi, S.R.; Grabosky, G.; Duda, T.; Geyerand, L.; Tachino, N. Speciation and population genetic structure in tropical Pacific sea urchins. Evolution 1997, 51, 1506–1517. [Google Scholar] [CrossRef] [PubMed]
- Palumbi, S.R. Using genetics as an indirect estimator of larval dispersal. In Ecology of Marine Invertebrate Larvae; McEdward, L., Ed.; CRC Press: New York, NY, USA, 1995; pp. 369–387. [Google Scholar]
- Caira, J.N.; Rodriguez, N.; Pickering, M. New African species of Echinobothrium (Cestoda: Diphyllidea) and implications for the identities of their skate hosts. J. Parasitol. 2013, 99, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Braun, C.D.; Gaube, P.; Afonso, P.; Fontes, J.; Skomal, G.B.; Thorrold, S.R. Assimilating electronic tagging, oceanographic modelling, and fisheries data to estimate movements and connectivity of swordfish in the North Atlantic. ICES J. Mar. Sci. 2019, 76, 2305–2317. [Google Scholar] [CrossRef]
- Reis-Santos, P.; Gillanders, B.M.; Sturrock, A.M.; Izzo, C.; Oxman, D.S.; Lueders-Dumont, J.A.; Hüssy, K.; Tanner, S.E.; Rogers, T.; Doubleday, Z.A.; et al. Reading the biomineralized book of life: Expanding otolith biogeochemical research and applications for fisheries and ecosystem-based management. Rev. Fish. Biol. Fish. 2023, 33, 411–449. [Google Scholar] [CrossRef]
- Pascual, M.; Rives, B.; Schunter, C.; Macpherson, E. Impact of life history traits on gene flow: A multispecies systematic review across oceanographic barriers in the Mediterranean Sea. PLoS ONE 2017, 12, e0176419. [Google Scholar] [CrossRef]
- Henriques, R.; Potts, W.M.; Santos, C.V.; Sauer, W.H.H.; Shaw, P.W. Population connectivity of an overexploited coastal fish, Argyrosomus coronus (Sciaenidae), in an ocean-warming hotspot. Afr. J. Mar. Sci. 2018, 40, 13–24. [Google Scholar] [CrossRef]
- Nielsen, E.S.; Henriques, R.; Toonen, R.J.; Knapp, I.S.S.; Guo, B.; von der Heyden, S. Complex signatures of genomic variation of two non-model marine species in a homogeneous environment. BMC Genom. 2018, 19, 347. [Google Scholar] [CrossRef]
- Seitz, A.C.; Courtney, M.B.; Evans, M.D.; Manishin, K. Pop-up satellite archival tags reveal evidence of intense predation on large immature Chinook salmon (Oncorhynchus tshawytscha) in the North Pacific Ocean. Can. J. Fish. Aquat. Sci. 2019, 76, 1608–1615. [Google Scholar] [CrossRef]
- Stockley, B.; Menezes, G.; Pinho, M.R.; Rogers, A.D. Genetic population structure in the black-spot sea bream (Pagellus bogaraveo Brunnich, 1768) from the NE Atlantic. Mar. Biol. 2005, 146, 793–804. [Google Scholar] [CrossRef]
- Belcaid, S.; Benchoucha, S.; Pérez Gil, J.L.; Gil-Herrera, J.; González Costas, F.; García Prieto, F.; Talbaoui, E.M.; El Arraf, S.; Hamdi, H.; Abid, N.; et al. Preliminary Joint Assessment of Pagellus Bogaraveo Stock of the Strait of Gibraltar Area between Spain and Morocco (GSAs 01 and 03); FAO-CopeMed II Occasional Paper No 15; Centro Oceanográfico de Vigo: Madrid, Spain, 2018; Volume 15, p. 18. [Google Scholar]
- Mytilineou, C.; Tsagarakis, K.; Bekas, P.; Anastasopoulou, A.; Kavadas, S.; Machias, A.; Haralabous, J.; Smith, C.J.; Petrakis, G.; Dokos, J.; et al. Spatial distribution and life-history aspects of blackspot seabream Pagellus bogaraveo (Osteichthyes: Sparidae). J. Fish. Biol. 2013, 83, 1551–1575. [Google Scholar] [CrossRef] [PubMed]
- Paruğ, Ş.; Cengiz, Ö. The maximum length record of the blackspot seabream (Pagellus bogaraveo Brünnich, 1768) for the entire Aegean Sea and Turkish territorial waters. Tur. J. Agr. For. 2020, 8, 2125–2130. [Google Scholar] [CrossRef]
- Chilari, A.; Petrakis, G.; Tsamis, E. Aspects of the biology of blackspot seabream (Pagellus bogaraveo) in the Ionian Sea, Greece. Fish. Res. 2006, 77, 84–91. [Google Scholar] [CrossRef]
- Lorance, P. History and dynamics of the overexploitation of the blackspot sea bream (Pagellus bogaraveo) in the Bay of Biscay. ICES J. Mar. Sci. 2011, 68, 290–301. [Google Scholar] [CrossRef]
- Pinho, M.; Diogo, H.; Carvalho, J.; Pereira, J.G. Harvesting juveniles of blackspot sea bream (Pagellus bogaraveo) in the Azores (Northeast Atlantic): Biological implications, management, and life cycle considerations. ICES J. Mar. Sci. 2014, 71, 2448–2456. [Google Scholar] [CrossRef]
- Sion, L.; Calculli, C.; Capezzuto, F.; Carlucci, R.; Carluccio, A.; Cornacchia, L.; Maiorano, P.; Pollice, A.; Ricci, P.; Tursi, A.; et al. Does the Bari Canyon (Central Mediterranean) influence the fish distribution and abundance? Prog. Oceanogr. 2019, 170, 81–92. [Google Scholar] [CrossRef]
- D’Onghia, G.; Indennidate, A.; Giove, A.; Savini, A.; Capezzuto, F.; Sion, L.; Vertino, A.; Maiorano, P. Distribution and behaviour of deep-sea benthopelagic fauna observed using towed cameras in the Santa Maria di Leuca cold-water coral province. Mar. Ecol. Prog. Ser. 2011, 443, 95–110. [Google Scholar] [CrossRef]
- Sanz-Fernández, V.; Gutiérrez-Estrada, J.C.; Pulido-Calvo, I.; Gil-Herrera, J.; Benchoucha, S.; el Arraf, S. Environment or catches? Assessment of the decline in blackspot seabream (Pagellus bogaraveo) abundance in the Strait of Gibraltar. J. Mar. Syst. 2019, 190, 15–24. [Google Scholar] [CrossRef]
- ICES. Working Group on Biology and Assessment of Deep-Sea Fisheries Resources (WGDEEP) 2020; ICES: Copenhagen, Denmark, 2020. [Google Scholar]
- ICES. Working Group on Biology and Assessment of Deep-Sea Fisheries Resources (WGDEEP) 2021; ICES: Copenhagen, Denmark, 2021. [Google Scholar]
- Carpenter, K.E.; Russell, B. Pagellus bogaraveo, Blackspot Seabream. The IUCN Red List of Threatened Species. Available online: https://www.iucnredlist.org/species/170244/1300216 (accessed on 21 May 2023).
- FAO. General Fisheries Commission for the Mediterranean—Report of the Twenty-Third Session of the Scientific Advisory Committee on Fisheries; FAO Fisheries and Aquaculture Report No. 1395; FAO Headquarters: Rome, Italy, 2022. [Google Scholar]
- ICES. Working Group on the Biology and Assessment of Deep-Sea Fisheries Resources (WGDEEP) 2019; ICES: Copenhagen, Denmark, 2019. [Google Scholar]
- ICES. Report of the Working Group on Biology and Assessment of Deep-Sea Fisheries Resources (WGDEEP) 2014; ICES: Copenhagen, Denmark, 2014. [Google Scholar]
- ICES. Report of the Working Group on the Biology and Assessment of Deep-Sea Fisheries Resources (WGDEEP) 2018; ICES: Copenhagen, Denmark, 2018. [Google Scholar]
- ICES. Working Group on the Biology and Assessment of Deep-Sea Fisheries Resources (WGDEEP) 2022; ICES Scientific Reports 2022; ICES: Copenhagen, Denmark, 2022. [Google Scholar]
- Nadal, I.; Sammartino, S.; García-Lafuente, J.; Sánchez Garrido, J.C.; Gil-Herrera, J.; Hidalgo, M.; Hernández, P. Hydrodynamic connectivity and dispersal patterns of a transboundary species (Pagellus bogaraveo) in the Strait of Gibraltar and adjacent basins. Fish. Oceanogr. 2022, 31, 384–401. [Google Scholar] [CrossRef]
- Fassatoui, C.; Mdelgi, E.; Romdhane, M.S. A preliminary investigation of allozyme genetic variation and population structure in common pandora (Pagellus erythrinus, Sparidae) from Tunisian and Libyan coasts. Ichthyol. Res. 2009, 56, 301–307. [Google Scholar] [CrossRef]
- Angiullim, E.; Sola, L.; Ardizzone, G.; Fassatoui, C.; Rossi, A.R. Phylogeography of the common pandora Pagellus erythrinus in the central Mediterranean Sea: Sympatric mitochondrial lineages and genetic homogeneity. Mar. Biol. Res. 2016, 12, 4–15. [Google Scholar] [CrossRef]
- Rossi, A.R.; Colangelo, P.; Berline, L.; Angiulli, E.; Ardizzone, G.; Fassatoui, C.; Sola, L. Influence of hydrodynamic connectivity on the genetic structure and gene flow of the common pandora Pagellus erythrinus. Hydrobiologia 2019, 834, 103–117. [Google Scholar] [CrossRef]
- Hidalgo, M.; Hermández, P.; Vasconcellos, M. Transboundary Population Structure of Sardine, European Hake and Blackspot Seabream in the Alboran Sea and Adjacent Waters: A Multidisciplinary Approach; GFCM Studies and Reviews; FAO: Rome, Italy, 2023; p. 69, in press. [Google Scholar]
- Bargelloni, L.; Alarcon, J.A.; Alvarez, M.C.; Penzo, E.; Magoulas, A.; Reis, C.; Patarnello, T. Discord in the family Sparidae (Teleostei): Divergent phylogeographical patterns across the Atlantic-Mediterranean divide. J. Evol. Biol. 2003, 16, 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Lemos, A.; Freitas, A.I.; Fernandes, A.T.; Goncalves, R.; Jesus, J.; Andrade, C.; Brehm, A. Microsatellite variability in natural populations of the blackspot seabream Pagellus bogaraveo (Brunnick, 1768): A database to access parentage assignment in aquaculture. Aquac. Res. 2006, 37, 1028–1033. [Google Scholar] [CrossRef]
- Piñera, J.A.; Blanco, G.; Vázquez, E.; Sánchez, J.A. Genetic diversity of blackspot seabream (Pagellus bogaraveo) populations of Spanish coasts: A preliminary study. Mar. Biol. 2007, 151, 2153–2158. [Google Scholar] [CrossRef]
- Robalo, J.I.; Farias, I.; Francisco, S.M.; Avellaneda, K.; Castilho, R.; Figueiredo, I. Genetic population structure of the Blackspot seabream (Pagellus bogaraveo): Contribution of mtDNA control region to fisheries management. Mitochondrial DNA Part A 2021, 32, 115–119. [Google Scholar] [CrossRef]
- Palma, J.; Andrade, J.P. Morphological study of Pagrus pagrus, Pagellus bogaraveo, and Dentex dentex (Sparidae) in the eastern Atlantic and the Mediterranean Sea. J. Mar. Biol. Assoc. UK 2004, 84, 449–454. [Google Scholar] [CrossRef]
- Hermida, M.; Cruz, C.; Saraiva, A. Parasites as biological tags for stock identification of blackspot Seabream, Pagellus bogaraveo, in Portuguese northeast Atlantic waters. Sci. Mar. 2013, 77, 607–615. [Google Scholar]
- MacKenzie, K.; Abaunza, P. Parasites as Biological Tags. In Stock Identification Methods: Applications in Fishery Science; Elsevier: Amsterdam, The Netherlands, 2004; pp. 211–226. [Google Scholar]
- Catalano, S.R.; Whittington, I.D.; Donnellan, S.C.; Gillanders, B.M. Parasites as biological tags to assess host population structure: Guidelines, recent genetic advances and comments on a holistic approach. Int. J. Parasitol. Parasites Wild 2014, 3, 220–226. [Google Scholar] [CrossRef]
- Bertrand, J.A.; Gil de Sola, L.; Papaconstantinou, C.; Relini, G.; Souplet, A. The general specifications of the MEDITS surveys. Sci. Mar. 2022, 66, 9. [Google Scholar] [CrossRef]
- Stockley, B.M.; Rogers, A.D.; Iyengar, A.; Menezes, G.; Santos, R.; Long, A. Ten microsatellite loci isolated and developed for the blackspot seabream, Pagellus bogaraveo (Brunnich 1768). Mol. Ecol. 2000, 9, 999–1000. [Google Scholar] [CrossRef] [PubMed]
- Piñera, J.A.; Bernardo, D.; Blanco, G.; Vazquez, E.; Sanchez, J.A. Isolation and characterization of polymorphic microsatellite markers in Pagellus bogaraveo, and cross-species amplification in Sparus aurata and Dicentrarchus labrax. Mol. Ecol. Notes 2006, 6, 33–35. [Google Scholar] [CrossRef]
- Ramšak, A.; Garoia, F.; Guarniero, I.; Mannini, P.; Tinti, F. Novel polymorphic microsatellite markers for the common pandora (Pagellus erythrinus). Mol. Ecol. Notes 2003, 3, 553–555. [Google Scholar] [CrossRef][Green Version]
- Guichoux, E.; Lagache, L.; Wagner, S.; Chaumeil, P.; Léger, P.; Lepais, O.; Lepoittevin, C.; Malausa, T.; Revardel, E.; Salin, F.; et al. Current trends in microsatellite genotyping. Mol. Ecol. Resour. 2011, 11, 591–611. [Google Scholar] [CrossRef] [PubMed]
- Flores-Rentería, L.; Krohn, A. Scoring Microsatellite Loci. In Without Lying Down; University of California Press: Berkeley, CA, USA, 2013; pp. 319–336. [Google Scholar]
- Raymond, M.; Rousset, F. GENEPOP (Version 1.2): Population Genetics Software for Exact Tests and Ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rousset, F. GENEPOP’007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Chapuis, M.P.; Estoup, A. Microsatellite Null Alleles and Estimation of Population Differentiation. Mol. Biol. Evol. 2007, 24, 621–631. [Google Scholar] [CrossRef]
- Belkhir, K.; Borsa, P.; Chikhi, L.; Raufaste, N.; Bonhomme, F. GENETIX 4.05, Logiciel Sous Windows TM Pour la Génétique des Populations. 1996. Available online: https://www.scienceopen.com/document?_vid=7cfcd230-1958-4cfc-a571-ce0ba003e63f (accessed on 1 April 2023).
- Goudet, J. FSTAT 2.9.3, a Program to Estimate and Test Gene Diversities and Fixation Indices. 2001. Available online: http://www2.unil.ch/popgen/softwares/fstat.htm (accessed on 15 March 2023).
- Kalinowski, S.T.; Wagner, A.P.; Taper, M.L. Ml-relate: A computer program for maximum likelihood estimation of relatedness and relationship. Mol. Ecol. Notes 2006, 6, 576–579. [Google Scholar] [CrossRef]
- Ryman, N.; Palm, S. POWSIM: A computer program for assessing statistical power when testing for genetic differentiation. Mol. Ecol. Notes 2006, 6, 600–602. [Google Scholar] [CrossRef]
- Ryman, N.; Palm, S.; André, C.; Carvalho, G.R.; Dahlgren, T.G.; Jorde, P.E.; Laikre, L.; Larsson, L.C.; Palmé, A.; Ruzzante, D.E. Power for detecting genetic divergence: Differences between statistical methods and marker loci. Mol. Ecol. 2006, 15, 2031–2045. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver. 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Smouse, P.E.; Quattro, J.M. Analysis of molecular variance inferred from metric distances among DNA haplotypes: Application to human mitochondrial DNA restriction Data. Genetics 1992, 131, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org (accessed on 15 March 2023).
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Kopelman, N.M.; Mayzel, J.; Jakobsson, M.; Rosenberg, N.A.; Mayrose, I. Clumpak: A program for identifying clustering modes and packaging population structure inferences across K. Mol. Ecol. Resour. 2015, 15, 1179–1191. [Google Scholar] [CrossRef]
- Luikart, G.; Cornuet, J.-M. Empirical Evaluation of a Test for Identifying Recently Bottlenecked Populations from Allele Frequency Data. Conserv. Biol. 1998, 12, 237. [Google Scholar] [CrossRef]
- Piry, S.; Luikart, G.; Cornuet, J.-M. Computer note. BOTTLENECK: A computer program for detecting recent reductions in the effective size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Cadrin, S.X. Defining spatial structure for fishery stock assessment. Fish. Res. 2020, 221, 105397. [Google Scholar] [CrossRef]
- Hedgecock, D.; Li, G.; Hubert, S.; Bucklin, K.; Ribes, V. Widespread null alleles and poor cross-species amplification of microsatellite DNA loci cloned from the pacific oyster, Crassostrea gigas. J. Shellfish Res. 2004, 23, 379–385. [Google Scholar]
- Peyran, C.; Planes, S.; Tolou, N.; Iwankow, G.; Boissin, E. Development of 26 highly polymorphic microsatellite markers for the highly endangered fan mussel Pinna nobilis and cross-species amplification. Mol. Biol. Rep. 2020, 47, 2551–2559. [Google Scholar] [CrossRef] [PubMed]
- Peng, Q.; Vijaya Satya, R.; Lewis, M.; Randad, P.; Wang, Y. Reducing amplification artifacts in high multiplex amplicon sequencing by using molecular barcodes. BMC Genom. 2015, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Delghandi, M.; Delghandi, M.P.; Goddard, S. The significance of PCR primer design in genetic diversity studies: Exemplified by recent research into the genetic structure of marine species. In PCR Primer Design. Methods in Molecular Biology; Basu, C., Ed.; Humana: New York, NY, USA, 2022; pp. 3–15. [Google Scholar] [CrossRef]
- Carlsson, J. Effects of microsatellite null alleles on assignment testing. J. Hered. 2008, 99, 616–623. [Google Scholar] [CrossRef]
- Yafeng, W.; Kentaro, U.; Wenjun, H.; Saneyoshi, U.; Weidong, X.; Gangbiao, X.; Yoshihiko, T. Null alleles in microsatellite markers. Biodivers. Sci. 2013, 21, 117–126. [Google Scholar] [CrossRef]
- Wu, X.; Wang, L.; Zhang, D.; Wen, Y. Microsatellite null alleles affected population genetic analyses: A case study of Maire yew (Taxus chinensis var. mairei). J. For. Res. 2019, 24, 230–234. [Google Scholar] [CrossRef]
- FAO CopeMed II. Report of the Joint Copemed—GFCM Data Preparation Meeting on Blackspot Seabream (Pagellus bogaraveo) in the Strait of Gibraltar; CopeMed II Tech Doc N. 51; Food and Agriculture Organisation of the United Nations: Malaga, Spain, 2019. [Google Scholar]
- Pitcher, T.J.; Morato, T.; Hart, P.J.; Clark, M.R.; Haggan, N.; Santos, R.S. (Eds.) Seamounts: Ecology, Fisheries and Conservation; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Gil-Herrera, J. Biología y Pesca del Voraz Pagellus bogaraveo en el Estrecho de Gibraltar. Ph.D. Thesis, Universidad de Cádiz, Cadiz, Spain, October 2006. [Google Scholar]
- Nadal, I.; Sammartino, S.; Garrido, J.C.S.; García-Lafuente, J. Tidal dynamics effect on the connectivity patterns of the blackspot seabream (Pagellus bogaraveo) in the Alboran Sea. In Proceedings of the II International Congress of Young Marine Researchers, Malaga, Spain, 1–4 October 2019; pp. 356–359, ISBN 978-84-09-13664-3. [Google Scholar]
- D’Iglio, C.; Albano, M.; Famulari, S.; Savoca, S.; Panarello, G.; Di Paola, D.; Perdichizzi, A.; Rinelli, P.; Lanteri, G.; Spanò, N.; et al. Intra- and interspecific variability among congeneric Pagellus otoliths. Sci. Rep. 2021, 11, 16315. [Google Scholar] [CrossRef]
- Almada, F.; Francisco, S.M.; Lima, C.S.; Fitzgerald, R.; Mirimin, L.; Villegas-Ríos, D.; Saborido-Rey, F.; Afonso, P.; Morato, T.; Bexiga, S.; et al. Historical gene flow constraints in a northeastern Atlantic fish: Phylogeography of the ballan wrasse Labrus bergylta across its distribution range. R. Soc. Open Sci. 2017, 4, 160773. [Google Scholar] [CrossRef]
- Stefanni, S.; Domingues, V.; Bouton, N.; Santos, R.S.; Almada, F.; Almada, V. Phylogeny of the shanny, Lipophrys pholis, from the NE Atlantic using mitochondrial DNA markers. Mol. Phylogenet. Evol. 2006, 39, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Patarnello, T.; Volckaert, F.A.M.J.; Castilho, R. Pillars of Hercules: Is the Atlantic-Mediterranean transition a phylogeographical break? Mol. Ecol. 2007, 16, 4426–4444. [Google Scholar] [CrossRef] [PubMed]
- Robalo, J.I.; Castilho, R.; Francisco, S.M.; Almada, F.; Knutsen, H.; Jorde, P.E.; Pereira, A.M.; Almada, V.C. Northern refugia and recent expansion in the North Sea: The case of the wrasse Symphodus melops (Linnaeus, 1758). Ecol. Evol. 2012, 2, 153–164. [Google Scholar] [CrossRef]
- Hauser, L.; Adcock, G.J.; Smith, P.J.; Ramírez, J.H.B.; Carvalho, G.R. Loss of microsatellite diversity and low effective population size in an overexploited population of New Zealand snapper (Pagrus auratus). Proc. Natl. Acad. Sci. USA 2002, 99, 11742–11747. [Google Scholar] [CrossRef] [PubMed]
- Árnason, E. Mitochondrial cytochrome B DNA variation in the high-fecundity atlantic cod: Trans-atlantic clines and shallow gene genealogy. Genetics 2004, 166, 1871. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, P. Large variance in reproductive success and the Ne/N ratio. Evolution 2005, 59, 1596–1599. [Google Scholar]
- Chévolot, M.; Hoarau, G.; Rijnsdorp, A.D.; Stam, W.T.; Olsen, J.L. Phylogeography and population structure of thornback ray (Raja clavata L., Rajidae). Mol. Ecol. 2006, 15, 3693–3705. [Google Scholar] [CrossRef]
- Cadrin, S.X.; Karr, L.A.; Mariani, S. Stock identification methods: An overview. In Stock Identification Methods; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1–5. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| FAO Fishing Area | FAO Zone | Regional Sea | Country | Location Name | Macro-Area/Macro-Area Code | Location Code | N |
|---|---|---|---|---|---|---|---|
| 27.8.c | Bay of Biscay—South | Atlantic Ocean | Spain | Santander | Atlantic Ocean NATL | STD | 26 |
| 27.9.a | Portugal | N. a. | POR | 36 | |||
| 27.9.a | Spain | Conil | COL | 18 | |||
| 27.9.a | Portuguese Waters—East | Atlantic Ocean/Alboran Sea | Morocco | Ksar Sghir | Western Gibraltar Strait GS | KSR | 15 |
| 27.9.a | Morocco | Tangier | TNG | 18 | |||
| 27.9.a | Morocco | Eddalya | EDL | 35 | |||
| 37.01.01 | Western Mediterranean—Balearic | Alboran Sea | Spain | MEDITS 2018 | Northern Alboran Sea NALB | SPA-18 | 10 |
| 37.01.01 | Spain | MEDITS 2019 | SPA-19 | 30 | |||
| 37.01.01 | Alboran Sea/Balearic Sea | Algeria | Ghazaouet | Southern Alboran Sea SALB | GHZ | 9 | |
| 37.01.01 | Balearic Sea | Algeria | Annaba | Central Mediterranean Sea CMED | ANB | 30 | |
| 37.01.01 | Balearic Sea/Tyrrhenian Sea | Tunisia | Tabarka | TBK | 30 | ||
| 37.01.03 | Western Mediterranean—Sardinia | Sea of Sicily | Italy | Marettimo | MZR | 16 | |
| 37.02.02 | Central Mediterranean—Ionian | Sea of Sicily | Malta | N. a. | MLT | 17 | |
| 37.02.02 | Ionian Sea | Greece | N. a. | Eastern Mediterranean Sea EMED | ION | 30 | |
| Total N | 320 |
| Locus | N. Alleles | Hexp | Hobs | Mean AR | FIS | HWE |
|---|---|---|---|---|---|---|
| Pb-OVI-B2 | 23 | 0.86 | 0.82 | 8.65 | 0.07 | 0.004119 |
| PbMS2 | 20 | 0.85 | 0.56 | 8.38 | 0.37 | <0.00 × 100 |
| PbMS6 | 37 | 0.94 | 0.87 | 12.39 | 0.10 | <4.91 × 10−12 |
| PbMS16 | 42 | 0.94 | 0.86 | 12.56 | 0.11 | <6.52 × 10−15 |
| PbMS19 | 10 | 0.67 | 0.59 | 4.24 | 0.15 | <9.43 × 10−7 |
| Pb-OVI-D108 | 7 | 0.46 | 0.46 | 2.89 | 0.00 | 7.27 × 10−6 |
| Pb-OVI-A5 | 41 | 0.91 | 0.83 | 10.95 | 0.13 | <5.30 × 10−11 |
| Pb-OVI-D114 | 11 | 0.81 | 0.74 | 6.36 | 0.12 | 0.00568 |
| Pb-OVI-D102 | 20 | 0.88 | 0.72 | 8.86 | 0.21 | <6.98 × 10−13 |
| Pb-OVI-D20 | 12 | 0.82 | 0.86 | 6.70 | −0.03 | 0.790455 |
| Pb-OVI-D21 | 14 | 0.84 | 0.87 | 7.38 | 0.00 | 0.978329 |
| Pb-OVI-D106 | 11 | 0.79 | 0.81 | 5.75 | 0.00 | 0.647982 |
| Pb-OVI-A3 | 22 | 0.88 | 0.71 | 9.16 | 0.22 | <2.32 × 10−15 |
| PbMS20 | 21 | 0.83 | 0.45 | 7.32 | 0.49 | <0.00 × 100 |
| PbMS18 | 23 | 0.83 | 0.50 | 8.37 | 0.42 | <0.00 × 100 |
| PbMS1 | 38 | 0.92 | 0.46 | 11.04 | 0.52 | <0.00 × 100 |
| Pb-OVI-D22 | 12 | 0.82 | 0.83 | 6.89 | 0.02 | 0.568561 |
| PbMS4 | 29 | 0.91 | 0.90 | 10.30 | 0.04 | <1.18 × 10−5 |
| PbMS15 | 40 | 0.92 | 0.74 | 11.70 | 0.23 | <3.24 × 10−31 |
| Pb-OVI-C103 | 11 | 0.76 | 0.72 | 5.53 | 0.08 | 0.02496 |
| STD | POR | COL | KSR | TNG | EDL | SPA-18 | SPA-19 | GHZ | ANB | TBK | MZR | MLT | ION | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| STD | 0.926 | 0.915 | 0.248 | 0.117 | 0.639 | 0.358 | 0.700 | 0.206 | 0.021 | 0.304 | 0.866 | 0.340 | 0.635 | |
| POR | −0.003 | 0.706 | 0.176 | 0.595 | 0.84 | 0.792 | 0.553 | 0.608 | 0.197 | 0.503 | 0.444 | 0.779 | 0.874 | |
| COL | −0.004 | −0.001 | 0.139 | 0.131 | 0.342 | 0.361 | 0.194 | 0.050 | 0.159 | 0.036 | 0.870 | 0.350 | 0.123 | |
| KSR | 0.006 | 0.006 | 0.009 | 0.641 | 0.344 | 0.091 | 0.075 | 0.196 | 0.024 | 0.015 | 0.731 | 0.394 | 0.307 | |
| TNG | 0.007 | 0.000 | 0.006 | 0.002 | 0.89 | 0.743 | 0.070 | 0.213 | 0.530 | 0.048 | 0.156 | 0.255 | 0.390 | |
| EDL | 0.000 | −0.001 | 0.002 | 0.004 | −0.002 | 0.693 | 0.395 | 0.163 | 0.189 | 0.104 | 0.499 | 0.435 | 0.439 | |
| SPA-18 | 0.003 | −0.002 | 0.002 | 0.015 | −0.002 | −0.001 | 0.477 | 0.059 | 0.073 | 0.065 | 0.330 | 0.300 | 0.173 | |
| SPA-19 | 0.000 | 0.001 | 0.004 | 0.010 | 0.008 | 0.002 | 0.002 | 0.569 | 0.001 | 0.044 | 0.590 | 0.297 | 0.172 | |
| GHZ | 0.007 | 0.000 | 0.012 | 0.013 | 0.008 | 0.007 | 0.015 | 0.001 | 0.644 | 0.454 | 0.066 | 0.434 | 0.201 | |
| ANB | 0.007 | 0.003 | 0.004 | 0.011 | 0.001 | 0.003 | 0.008 | 0.011 | −0.001 | 0.001 | 0.004 | 0.065 | 0.019 | |
| TBK | 0.002 | 0.001 | 0.008 | 0.014 | 0.008 | 0.004 | 0.009 | 0.006 | 0.002 | 0.010 | 0.041 | 0.423 | 0.088 | |
| MZR | −0.003 | 0.002 | −0.004 | 0.000 | 0.008 | 0.002 | 0.004 | 0.001 | 0.015 | 0.014 | 0.009 | 0.322 | 0.514 | |
| MLT | 0.002 | −0.002 | 0.001 | 0.004 | 0.004 | 0.001 | 0.003 | 0.003 | 0.001 | 0.006 | 0.001 | 0.003 | 0.122 | |
| ION | 0.000 | −0.002 | 0.004 | 0.004 | 0.002 | 0.001 | 0.005 | 0.003 | 0.005 | 0.006 | 0.004 | 0.000 | 0.004 |
| Source of Variation | % of Variation | F Statistics | p-Value | |
|---|---|---|---|---|
| Scenario 1 | ||||
| Among population | 0.20 | FST | 0.002 | 1.00 |
| Among individuals within population | 5.17 | FIS | 0.052 | 0.000 |
| Within individuals | 94.63 | FIT | 0.054 | 0.000 |
| Scenario 2 | ||||
| Among groups | −0.12 | FCT | −0.001 | 0.815 |
| Among population within groups | 0.31 | FSC | 0.003 | 0.037 |
| Among individuals within population | 5.17 | FIS | 0.052 | 0.000 |
| Within individuals | 94.64 | FIT | 0.054 | 0.000 |
| Scenario 3 | ||||
| Among groups | 0.26 | FCT | 0.003 | 0.014 |
| Among population within groups | 0.10 | FSC | 0.001 | 0.162 |
| Among individuals Within population | 5.16 | FIS | 0.052 | 0.000 |
| Within individuals | 94.48 | FIT | 0.055 | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrari, A.; Spiga, M.; Rodriguez, M.D.; Fiorentino, F.; Gil-Herrera, J.; Hernandez, P.; Hidalgo, M.; Johnstone, C.; Khemiri, S.; Mokhtar-Jamaï, K.; et al. Matching an Old Marine Paradigm: Limitless Connectivity in a Deep-Water Fish over a Large Distance. Animals 2023, 13, 2691. https://doi.org/10.3390/ani13172691
Ferrari A, Spiga M, Rodriguez MD, Fiorentino F, Gil-Herrera J, Hernandez P, Hidalgo M, Johnstone C, Khemiri S, Mokhtar-Jamaï K, et al. Matching an Old Marine Paradigm: Limitless Connectivity in a Deep-Water Fish over a Large Distance. Animals. 2023; 13(17):2691. https://doi.org/10.3390/ani13172691
Chicago/Turabian StyleFerrari, Alice, Martina Spiga, Miriam Dominguez Rodriguez, Fabio Fiorentino, Juan Gil-Herrera, Pilar Hernandez, Manuel Hidalgo, Carolina Johnstone, Sana Khemiri, Kenza Mokhtar-Jamaï, and et al. 2023. "Matching an Old Marine Paradigm: Limitless Connectivity in a Deep-Water Fish over a Large Distance" Animals 13, no. 17: 2691. https://doi.org/10.3390/ani13172691
APA StyleFerrari, A., Spiga, M., Rodriguez, M. D., Fiorentino, F., Gil-Herrera, J., Hernandez, P., Hidalgo, M., Johnstone, C., Khemiri, S., Mokhtar-Jamaï, K., Nadal, I., Pérez, M., Sammartino, S., Vasconcellos, M., & Cariani, A. (2023). Matching an Old Marine Paradigm: Limitless Connectivity in a Deep-Water Fish over a Large Distance. Animals, 13(17), 2691. https://doi.org/10.3390/ani13172691