Genomic Characterization of Canis Familiaris Papillomavirus Type 25, a Novel Papillomavirus Associated with a Viral Plaque from the Pinna of a Dog


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
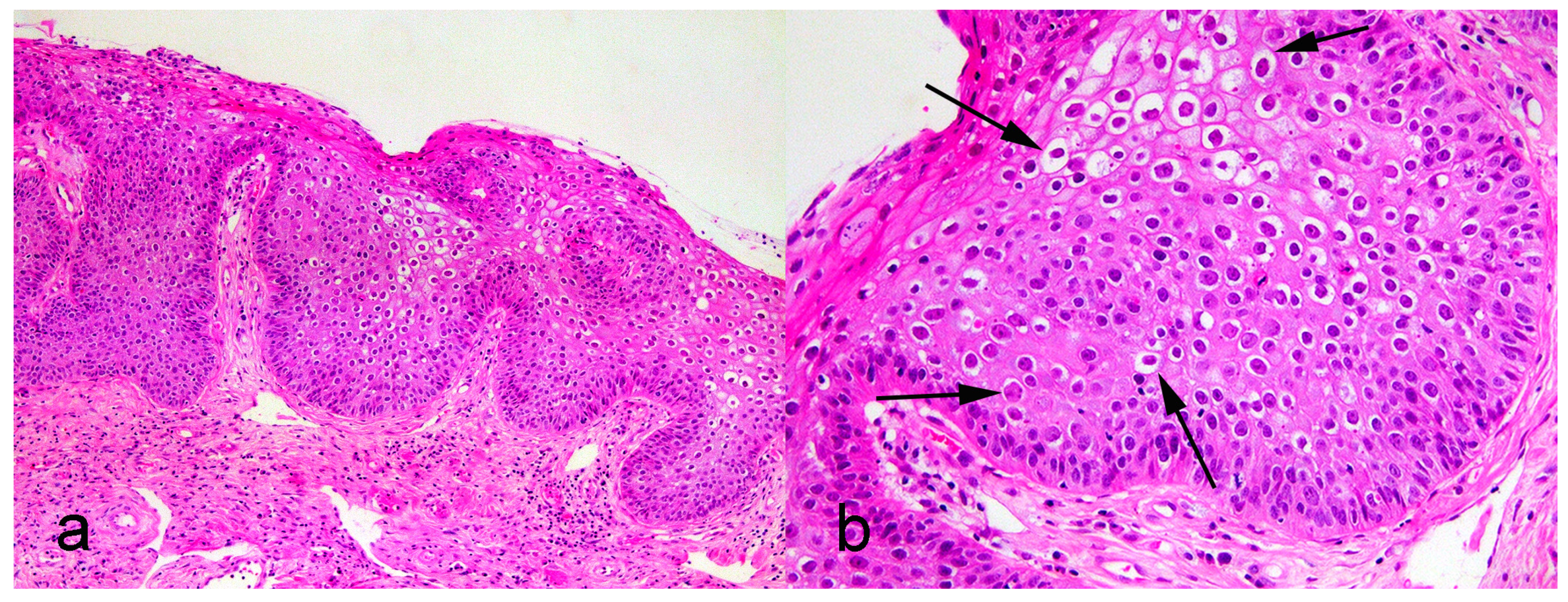
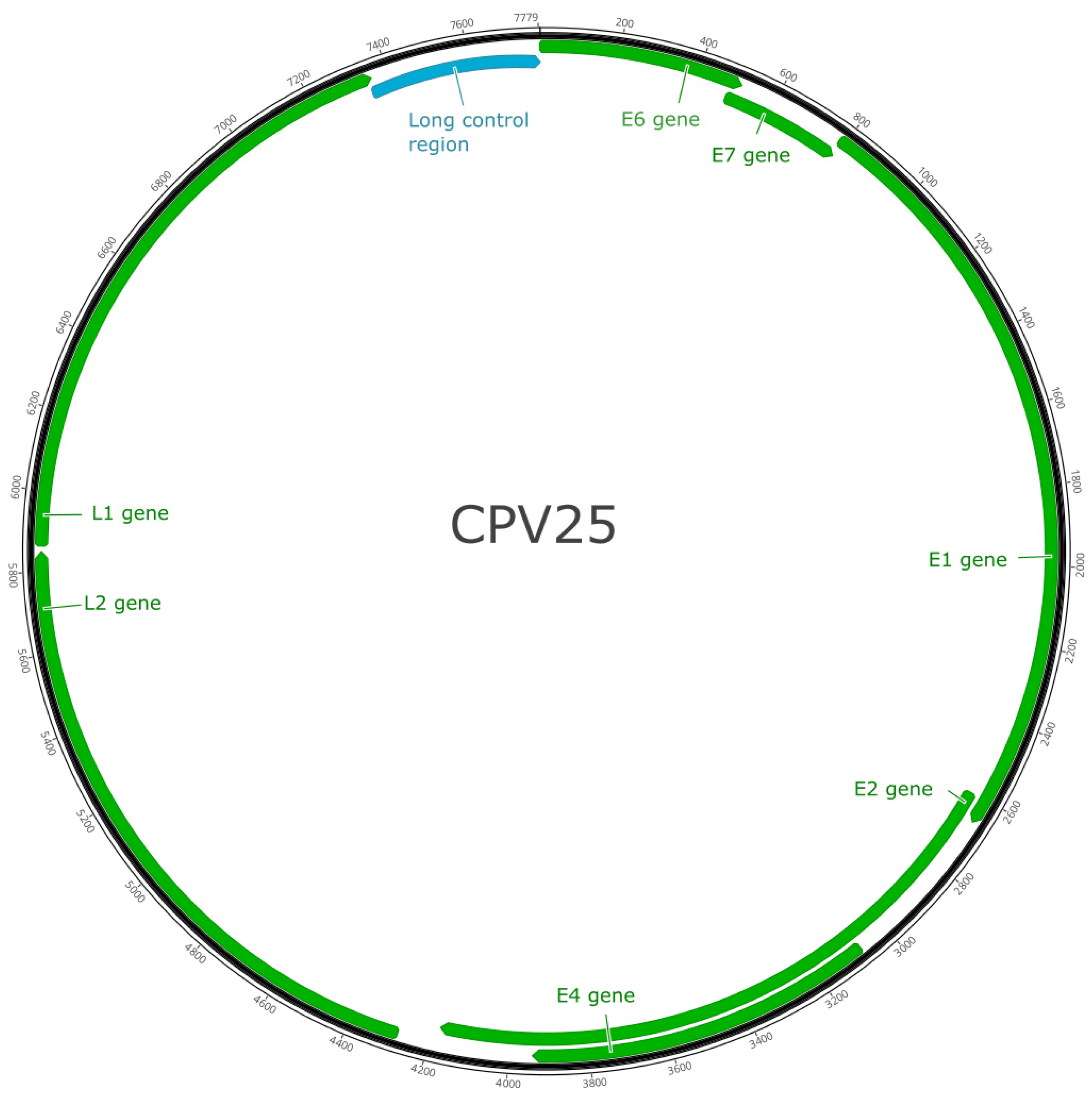
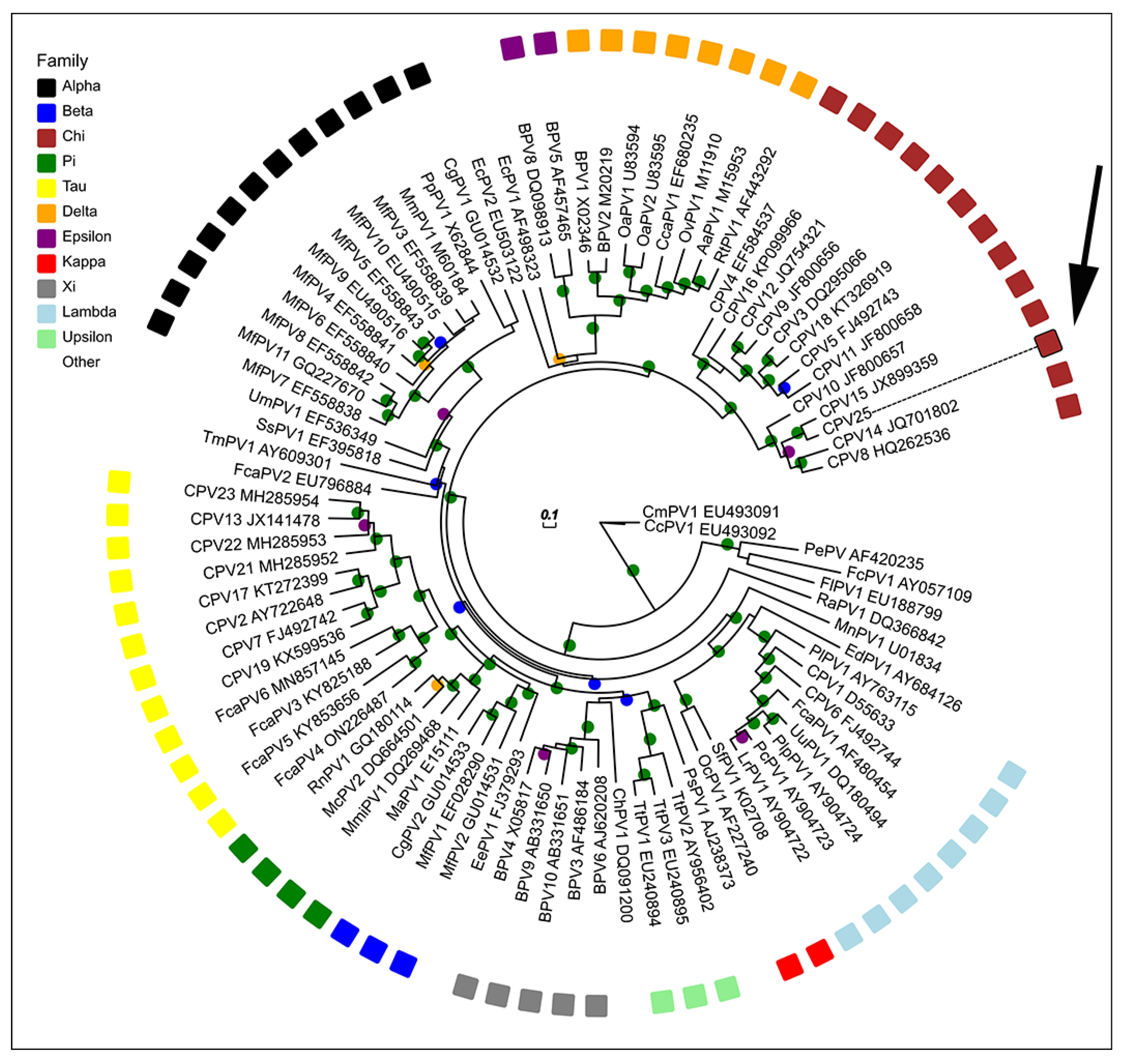
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Munday, J.S.; Knight, C.G.; Luff, J.A. Papillomaviral skin diseases of humans, dogs, cats and horses: A comparative review. Part 2: Pre-neoplastic and neoplastic diseases. Vet. J. 2022, 288, 105898. [Google Scholar] [CrossRef] [PubMed]
- Bernard, H.U.; Burk, R.D.; Chen, Z.; van Doorslaer, K.; Hausen, H.; de Villiers, E.M. Classification of papillomaviruses (PVs) based on 189 PV types and proposal of taxonomic amendments. Virology 2010, 401, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Munday, J.S.; Gedye, K.; Knox, M.A.; Ravens, P.; Lin, X. Genomic characterisation of Canis familiaris papillomavirus type 24, a novel papillomavirus associated with extensive pigmented plaque formation in a Pug dog. Viruses 2022, 14, 2357. [Google Scholar] [CrossRef] [PubMed]
- Luff, J.; Rowland, P.; Mader, M.; Orr, C.; Yuan, H. Two canine papillomaviruses associated with metastatic squamous cell carcinoma in two related Basenji dogs. Vet. Pathol. 2016, 53, 1160–1163. [Google Scholar] [CrossRef] [PubMed]
- Munday, J.S.; Lam, A.T.H.; Sakai, M. Extensive progressive pigmented viral plaques in a Chihuahua dog. Vet. Dermatol. 2022, 33, 252–254. [Google Scholar] [CrossRef] [PubMed]
- Munday, J.S.; Thomson, N.A.; Luff, J.A. Papillomaviruses in dogs and cats. Vet. J. 2017, 225, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Munday, J.S.; Orbell, G.; Robinson, L. Detection of a novel papillomaviral sequence in viral plaques confined to the pinna of a dog. Vet. Dermatol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Ashton, B.; Buxton, S.; Cheung, M.; Cooper, A.; Duran, C.; Field, M.; Heled, J.; Kearse, M.; Markowitz, S.; et al. Geneious, v5.3; 2010. Available online: https://www.geneious.com (accessed on 19 April 2023).
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Zou, N.; Lin, B.Y.; Chow, L.T.; Harper, J.W. Interaction between cyclin-dependent kinases and human papillomavirus replication-initiation protein E1 is required for efficient viral replication. Proc. Natl. Acad. Sci. USA 1999, 96, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Antonsson, A.; Hansson, B.G. Healthy skin of many animal species harbors papillomaviruses which are closely related to their human counterparts. J. Virol. 2002, 76, 12537–12542. [Google Scholar] [CrossRef] [PubMed]
- Stokking, L.B.; Ehrhart, E.J.; Lichtensteiger, C.A.; Campbell, K.L. Pigmented epidermal plaques in three dogs. J. Am. Anim. Hosp. Assoc. 2004, 40, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Sundberg, J.P.; Van Ranst, M.; Montali, R.; Homer, B.L.; Miller, W.H.; Rowland, P.H.; Scott, D.W.; England, J.J.; Dunstan, R.W.; Mikaelian, I.; et al. Feline papillomas and papillomaviruses. Vet. Pathol. 2000, 37, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Munger, K.; Baldwin, A.; Edwards, K.M.; Hayakawa, H.; Nguyen, C.L.; Owens, M.; Grace, M.; Huh, K. Mechanisms of human papillomavirus-induced oncogenesis. J. Virol. 2004, 78, 11451–11460. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| ORF | ORF Location | Length (nt) | Length (aa) | Molecular Mass (kDa) | pI |
|---|---|---|---|---|---|
| E1 | 783–2663 | 1881 | 626 | 71.18 | 5.53 |
| E2 | 2605–4176 | 1572 | 524 | 56.63 | 7.36 |
| E4 | 3119–3940 | 822 | 274 | 31.48 | 10.37 |
| E6 | 1–507 | 507 | 168 | 19.08 | 6.72 |
| E7 | 482–793 | 312 | 103 | 11.65 | 4.53 |
| L1 | 5863–7356 | 1494 | 497 | 56.02 | 7.37 |
| L2 | 4279–5850 | 1572 | 523 | 56.33 | 5.08 |
| Papillomavirus | Host Species | Classification | L1 Similarity (%) |
|---|---|---|---|
| Canine familiaris papillomavirus 15 (JX899359) | Domestic dog | ChiPV3 | 83.9 |
| Canine familiaris papillomavirus 8 (HQ262536) | Domestic dog | ChiPV3 | 70.8 |
| Canine familiaris papillomavirus 10 (JF800657) | Domestic dog | ChiPV3 | 69.5 |
| Canine familiaris papillomavirus 14 (JQ701802) | Domestic dog | ChiPV3 | 67.8 |
| Canine familiaris papillomavirus 12 (JQ754321) | Domestic dog | ChiPV1 | 62.3 |
| Canine familiaris papillomavirus 4 (EF584537) | Domestic dog | ChiPV2 | 61.1 |
| Canine familiaris papillomavirus 1 (D55633) | Domestic dog | LambdaPV | 55.5 |
| Canine familiaris papillomavirus 2 (AY722648) | Domestic dog | TauPV | 54.5 |
| Felis catis papillomavirus 2 (EU796884) | Domestic cat | DyothetaPV1 | 59.1 |
| Sus scrofa papillomavirus 1 (EF395818) | Domestic pig | DyodeltaPV | 59.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munday, J.S.; Gedye, K.; Knox, M.A.; Robinson, L.; Lin, X. Genomic Characterization of Canis Familiaris Papillomavirus Type 25, a Novel Papillomavirus Associated with a Viral Plaque from the Pinna of a Dog. Animals 2023, 13, 1859. https://doi.org/10.3390/ani13111859
Munday JS, Gedye K, Knox MA, Robinson L, Lin X. Genomic Characterization of Canis Familiaris Papillomavirus Type 25, a Novel Papillomavirus Associated with a Viral Plaque from the Pinna of a Dog. Animals. 2023; 13(11):1859. https://doi.org/10.3390/ani13111859
Chicago/Turabian StyleMunday, John S., Kristene Gedye, Matthew A. Knox, Lynne Robinson, and Xiaoxiao Lin. 2023. "Genomic Characterization of Canis Familiaris Papillomavirus Type 25, a Novel Papillomavirus Associated with a Viral Plaque from the Pinna of a Dog" Animals 13, no. 11: 1859. https://doi.org/10.3390/ani13111859
APA StyleMunday, J. S., Gedye, K., Knox, M. A., Robinson, L., & Lin, X. (2023). Genomic Characterization of Canis Familiaris Papillomavirus Type 25, a Novel Papillomavirus Associated with a Viral Plaque from the Pinna of a Dog. Animals, 13(11), 1859. https://doi.org/10.3390/ani13111859