
Morphology of the Ventral Process of the Sixth Cervical Vertebra in Extinct and Extant Equus: Functional Implications


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Terminology
2.3. Materials
2.4. Methods
3. Results
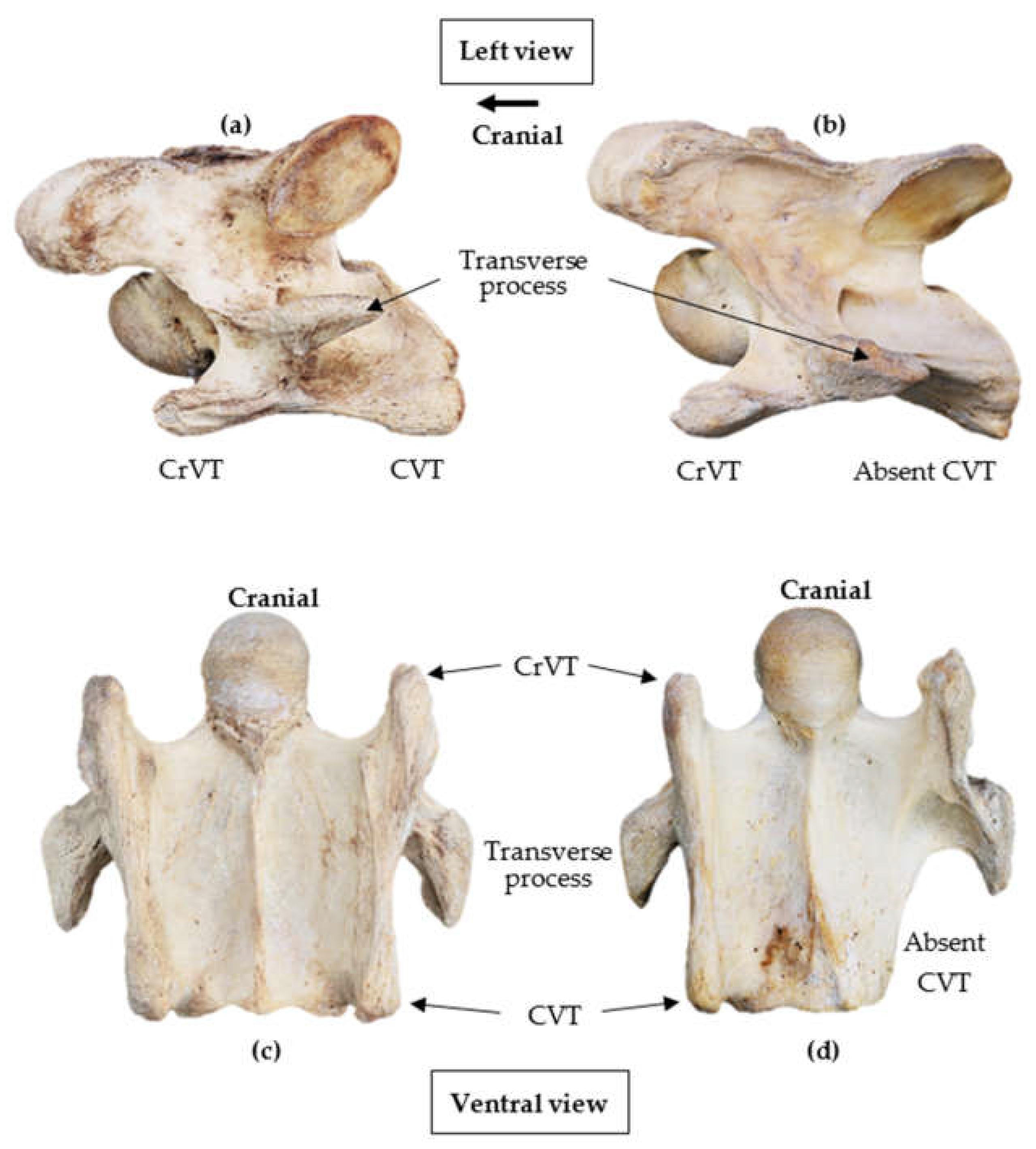
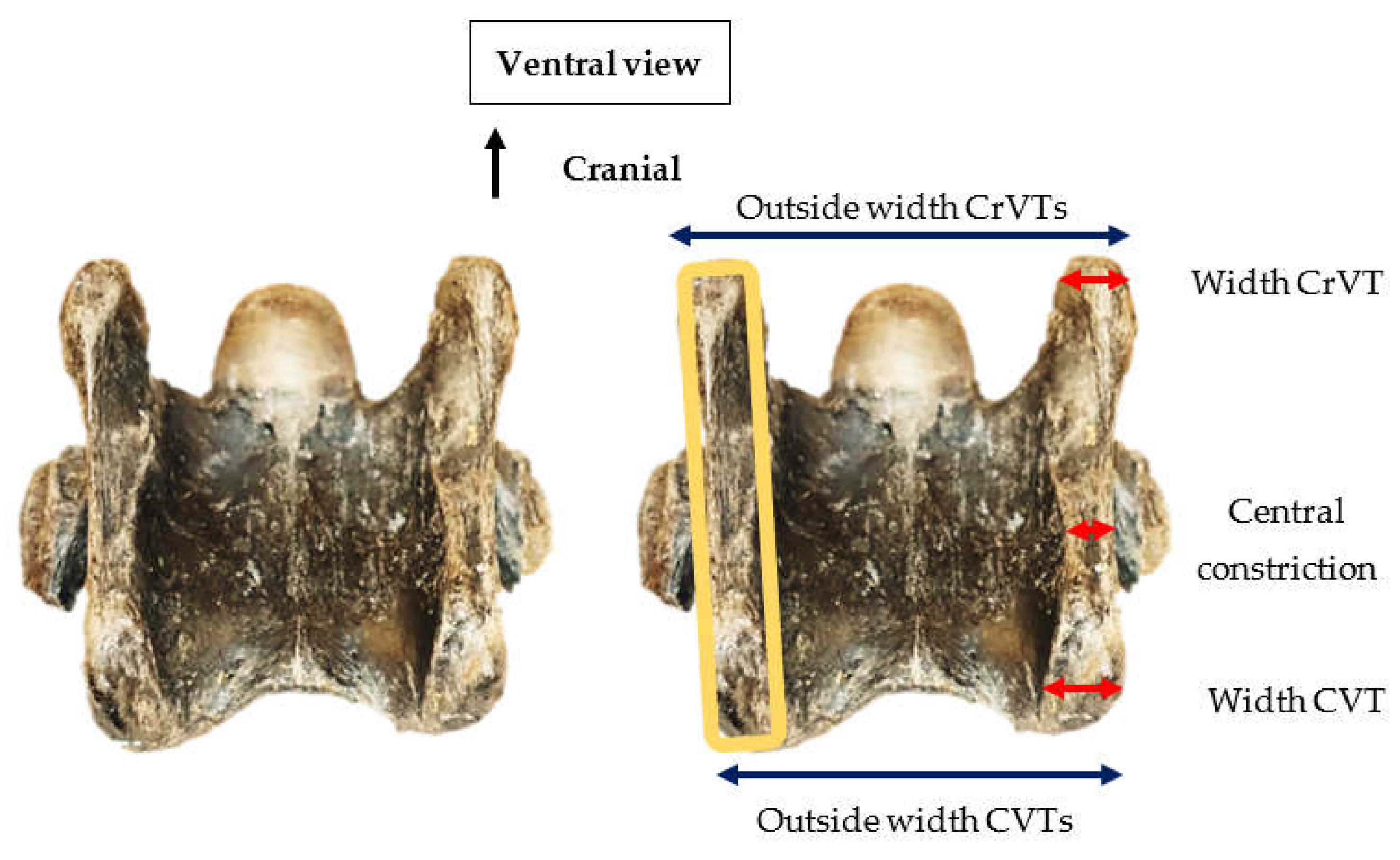
3.1. General Anatomy
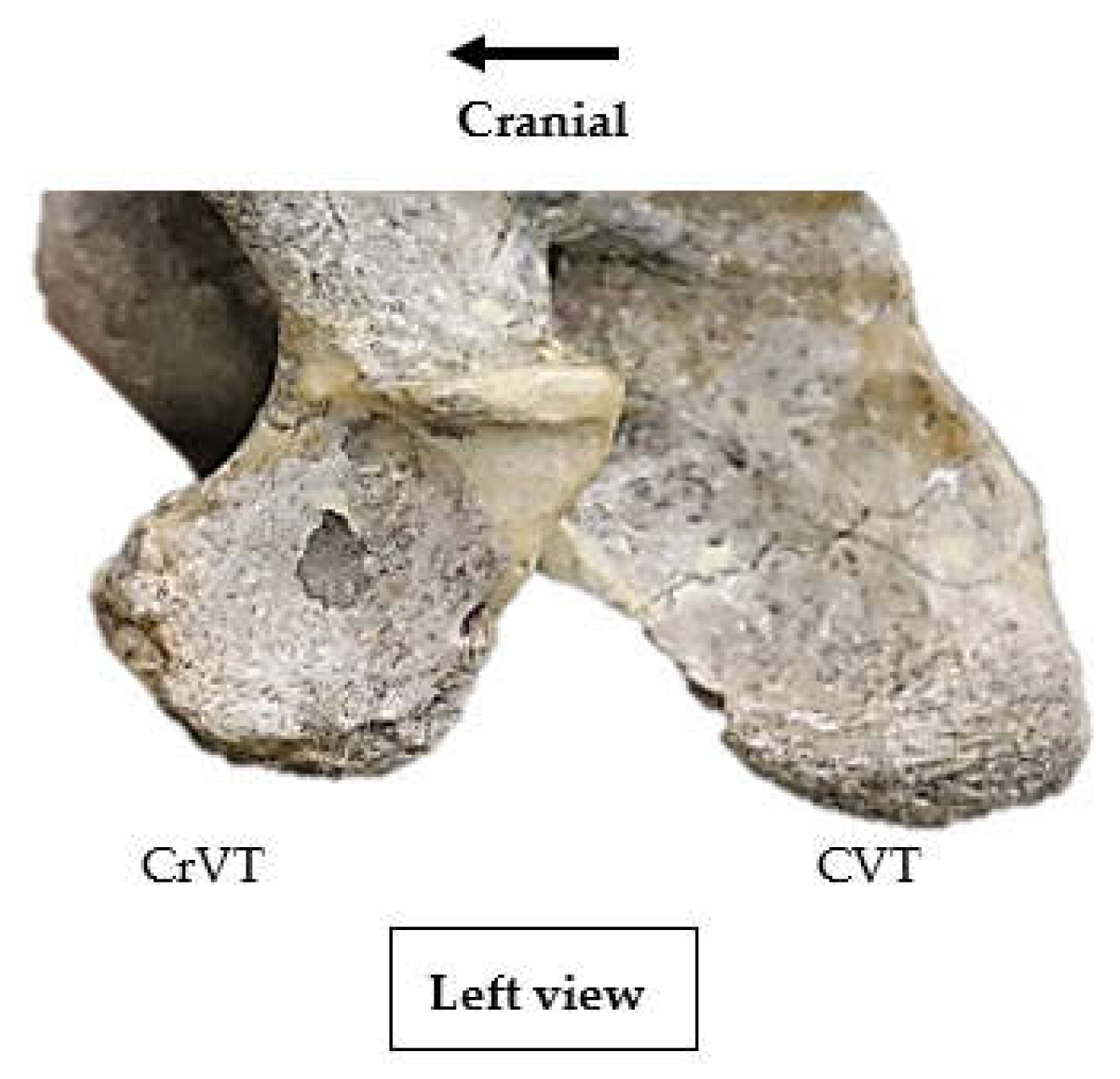
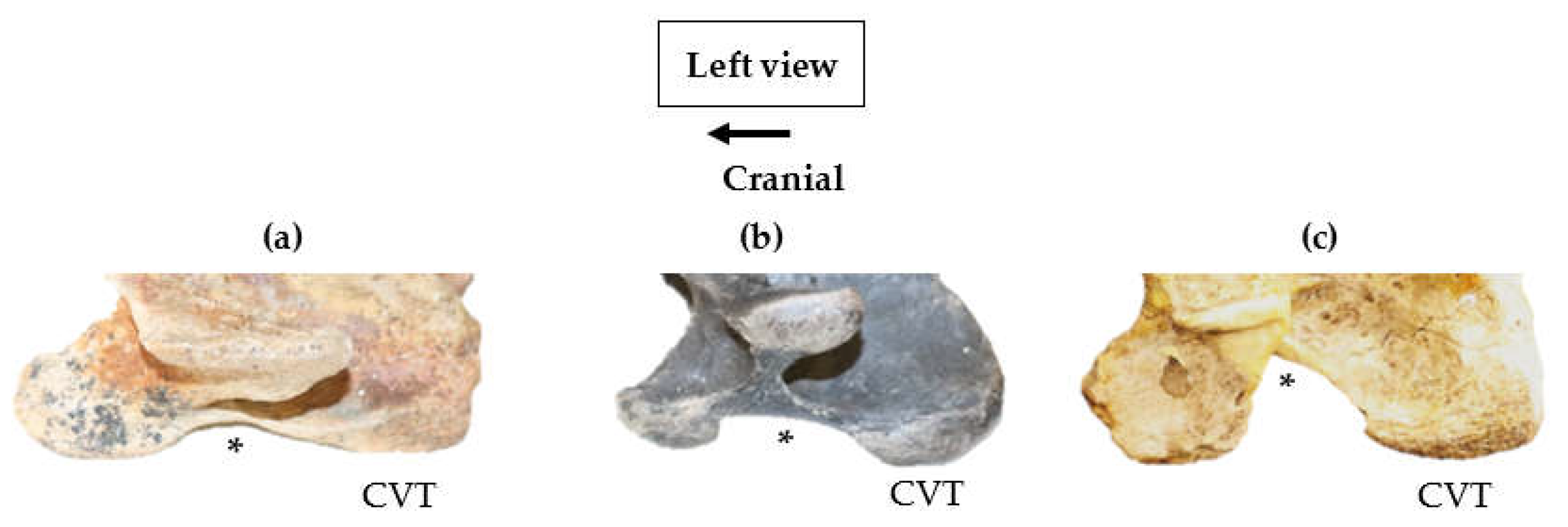
3.2. Lateral Profile of the Ventral Process
3.3. Variations of the Eqpiphyseal Growth Plate
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arnold, P. Evolution of the mammalian neck and development, morpho-functional, and paleontological perspectives. J. Mamm. Evol. 2021, 28, 173–183. [Google Scholar] [CrossRef]
- Arnold, P.; Amson, E.; Fischer, M.S. Differential scaling patterns of vertebrae and the evolution of the neck length in mammals. Evolution 2017, 71, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Buchholtz, E.A.; Bailin, H.G.; Laves, S.A.; Yang, J.T.; Chan, M.-Y.; Drozd, L.E. Fixed cervical count and the origin of the mammalian diaphragm. Evol. Dev. 2012, 5, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Willmore, K.E. The Body Plan Concept and Its Centrality in Evo-Devo. Evol. Educ. Outreach 2012, 5, 219–230. [Google Scholar] [CrossRef]
- Flower, W.H. An Introduction to the Osteology of the Mammalia, 3rd ed.; Gadow, H., Ed.; Macmillion: London, UK, 1885; pp. 12–32. [Google Scholar]
- Chauveau, A. The Comparative Anatomy of the Domesticated Animals, 2nd ed.; Fleming, G.D., Ed.; Appleton and Company: New York, NY, USA, 1905; pp. 7–167. [Google Scholar]
- Getty, R. Equine Osteology. In The Anatomy of the Domestic Animals, 5th ed.; Sisson, S., Grossman, J.D., Eds.; Saunders: Philadelphia, PA, USA, 1975; pp. 255–348. [Google Scholar]
- Frandson, R.D.; Wilke, W.L.; Fails, A.D. Anatomy and Physiology of Farm Animals, 7th ed.; Wiley-Blackwell: Ames, IA, USA, 2009; pp. 59–71. [Google Scholar]
- Janis, C.M. New ideas in ungulate phylogeny and evolution. Trends Ecol. Evol. 1988, 11, 291–297. [Google Scholar] [CrossRef]
- Prothero, D.R. Evolutionary transitions in the fossil record of terrestrial hoofed mammals. Evol. Educ. Outreach 2009, 2, 289–302. [Google Scholar] [CrossRef][Green Version]
- Narita, Y.; Kuratani, S. Evolution of the vertebral formulae in mammals: A perspective on developmental constraints. J. Exp. Zool. 2005, 304, 91–106. [Google Scholar] [CrossRef]
- Sisson, S. Osteology. In A Textbook of Veterinary Anatomy; W.B. Saunders Company: Philadelphia, PA, USA, 1910; pp. 31–35. [Google Scholar]
- Bradley, O. The Topical Anatomy of the Head and Neck of the Horse, 2nd ed.; Green and Sons: Edinburgh, UK, 1947. [Google Scholar]
- Denoix, J.-M.; Pailloux, J.-P. Anatomy and Basic Biomechanical Concepts. In Physical Therapy and Massage for the Horse, 2nd ed.; Trafalgar Square Publishing: North Pomfret, VT, USA, 2001; pp. 19–69. [Google Scholar]
- Bainbridge, D. The Normal Anatomy of the Neck. In Equine Neck and Back Pathology, 2nd ed.; Henson, F.M.D., Ed.; John Wiley and Sons Ltd.: Hoboken, UK, 2018; pp. 1–9. [Google Scholar]
- Rombach, N.; Stubbs, N.; Clayton, H. Gross anatomy of the deep perivertebral musculature in horses. Am. J. Vet. Res. 2014, 75, 433–440. [Google Scholar] [CrossRef]
- Arnold, P.; Esteve-Altava, B.; Fischer, M.S. Musculoskeletal networks reveal topological disparity in mammalian neck evolution. BMC Evol. Biol. 2017, 17, 251. [Google Scholar] [CrossRef]
- Davies, Z. Equine Science, 2nd ed.; John and Sons Inc.: Hoboken, NJ, USA, 2018; pp. 80–135. [Google Scholar]
- Haussler, K.K. Functional Anatomy and Clinical Biomechanics of the Equine Cervical Spine. In Proceedings of the Association of American Equine Practitioners, 360° Pain in the Neck Conference, Fort Collins, CO, USA, June 2016; Available online: https://www.researchgate.net/publication/316053036 (accessed on 18 March 2021).
- Popesko, P. Atlas of Topographical Anatomy of the Domestic Animals, 4th ed.; W.B. Saunders: Philadelphia, PA, USA, 1985; Volume 4, p. 185. [Google Scholar]
- Ballou, W.R. A Compendium of Equine Anatomy and Physiology; Replicated by Kessinger Legacy Reprints; P. Blakiston’s Son and Co.: Philadelphia, PA, USA, 1907. [Google Scholar]
- Ashdown, R.R.; Done, S.H. Color Atlas of Veterinary Anatomy, 2nd ed.; Mosby-Wolfe: London, UK, 2000. [Google Scholar]
- Budras, K.-D.; Sack, W.O.; Röck, S. Anatomy of the Horse, 5th ed.; Schlütersche Verlagsgesellschaft & Co.: Hannover, Germany, 2009. [Google Scholar]
- De Lahunta, A. Veterinary Neuroanatomy and Clinical Neurology, 4th ed.; Elsevier: St. Louis, MO, USA, 2015. [Google Scholar]
- Dyce, K.M.; Sack, W.O.; Wensing, C.J. Textbook of Veterinary Anatomy, 5th ed.; Elsevier: Pennsylvania, PA, USA, 2018. [Google Scholar]
- Fails, A.D. Functional anatomy of the equine musculoskeletal system. In Adams and Stashak’s Lameness in Horses, 7th ed.; Baxter, G., Ed.; Blackwell Publishing: Oxford, UK, 2020; pp. 1–65. [Google Scholar]
- Hodgins, J.E.; Haskett, T.H. The Anatomy and Diseases and Treatment of Domestic Animals; The Veterinary Science Company: Detroit, MI, USA, 1905. [Google Scholar]
- Percivall, W. Anatomy, Physiology, and Pathology of the Horse; Longman, Hurst, Rees, Orme, Brown & Green: London, UK, 1826; Available online: https://archive.org/ (accessed on 10 January 2019).
- Steel, J.H. Outlines of Equine Anatomy: A Manual for the Use of Veterinary Students in the Dissection Room; Replicated by Hardpress Publishing; William Wood and Co.: New York, NY, USA, 1876. [Google Scholar]
- Agerholm, J.S.; Bendixen, C.; Andersen, O.; Arnbjerg, J. Complex vertebral malformation in Holstein calves. J. Vet. Diagn. Investig. 2001, 13, 283–289. [Google Scholar] [CrossRef]
- Agerholm, J.S. Complex vertebral malformation Syndrome in Holstein cattle: The Story so Far. Acta Vet. Scan. 2007, 49, S5. [Google Scholar] [CrossRef][Green Version]
- Nielsen, U.S.; Aamand, G.P.; Andersen, O.; Bendixen, C.; Nielsen, V.H.; Agerholm, J.S. Effects of complex vertebral malformation on fertility traits in Holstein cattle. Livest. Prod. Sci. 2003, 79, 233–238. [Google Scholar] [CrossRef]
- May-Davis, S. The occurrence of a congenital malformation in the sixth and seventh cervical vertebrae predominantly observed in thoroughbred horses. J. Equine. Vet. Sci. 2014, 18, 1313–1317. [Google Scholar] [CrossRef]
- DeRouen, A.; Spriet, M.; Aleman, M. Prevalence of anatomical variation of the sixth cervical vertebra and association with vertebral canal stenosis and articular process osteoarthritis in the horse. Vet. Radiol. Ultrasound 2016, 57, 253–258. [Google Scholar] [CrossRef]
- Santinelli, I.; Beccati, F.; Arcelli, R. Anatomical variation of the spinous and transverse processes in the caudal cervical vertebrae and the first thoracic vertebra in horses. Equine Vet. J. 2016, 48, 45–49. [Google Scholar] [CrossRef]
- Veraa, S.; Bergmann, W.; van den Belt, A.-J.; Wijnberg, I.; Back, W. Ex vivo computed tomographic evaluation of morphology variations in equine cervical vertebrae. Vet. Radiol. Ultrasound 2016, 57, 482–488. [Google Scholar] [CrossRef]
- Veraa, S.; de Graaf, K.; Wijnberg, I.D.; Back, W.; Vernooij, H.; Nielen, M.; Belt, A.J. Caudal Cervical Vertebral Morphological Variation is not Associated with Clinical Signs in Warmblood Horses. Equine Vet. J. 2019, 52, 210–224. [Google Scholar] [CrossRef]
- Beccati, F.; Pepe, M.; Santinelli, I.; Gialletti, R.; Di Meo, A.; Romero, J.M. Radiographic findings and anatomical variations of the caudal cervical area in horses with neck pain and ataxia: Case–control study on 116 horses. Vet. Rec. 2020, 187, e79. [Google Scholar] [CrossRef]
- May-Davis, S. Congenital Malformations of the 1st Sternal Rib. J. Equine Vet. Sci. 2014, 49, 92–100. [Google Scholar] [CrossRef]
- May-Davis, S.; Walker, C. Variations and Implications of the Gross Morphology in the Longus colli Muscle in Thoroughbred and Thoroughbred Derivative Horses Presenting with a Congenital Malformation of the Sixth and Seventh Cervical Vertebrae. J. Equine Vet. Sci. 2015, 35, 560–568. [Google Scholar] [CrossRef]
- Witzmann, F.; Haridy, Y.; Hilger, A.; Manke, I.; Asbach, P. Rarity of congenital malformation and deformity in the fossil record of vertebrates—A non-human perspective. Int. J. Paeopatho. 2021, 33, 30–42. [Google Scholar] [CrossRef]
- Van der Geer, A.A.E.; Galis, F. High incidence of cervical ribs indicates vulnerable condition in Late Pleistocene woolly rhinoceroses. PeerJ 2017, 5, e3684. [Google Scholar] [CrossRef] [PubMed]
- Reumer, J.W.F.; ten Broek, C.M.A.; Galis, F. Extraordinary incidence of cervical ribs indicates vulnerable condition in Late Pleistocene mammoths. PeerJ 2014, 2, 318. [Google Scholar] [CrossRef][Green Version]
- Varela-Lasheras, I.; Bakker, A.J.; van der Mije, S.D.; Metz, J.A.J.; van Alphen, J.; Galis, F. Breaking evolutionary and pleiotropic constraints in mammals: On sloths, manatees and homeotic mutations. EvoDevo 2011, 2, 11. [Google Scholar] [CrossRef]
- Gasse, H.W.; Van Den, B.; Simeons, P. Nomina Anatomica Veterinaria, 6th ed.; Editorial Committee for the World Association of Veterinary Anatomists: Hanover, NJ, USA; Ghent, Belgium; Columbia, MO, USA; Rio de Janeiro, Brazil, 2017; p. 17. [Google Scholar]
- Gidley, J.W. A new three-toed horse. J. Morhol. 1903, 29, 465–476. [Google Scholar]
- Wood, A.R.; Bebej, R.M.; Manz, C.L.; Begun, D.L.; Gingerich, P.D. Postcranial functional morphology of Hyracotherium (Equidae, Perissodactyla) and locomotion in the earliest horse. J. Mammal. Evol. 2011, 18, 1–32. [Google Scholar] [CrossRef]
- Franzen, J.F.; Habersetzer, J. Complete skeleton of Eurohippus messelensis (Mammalia, Perissodactyla, Equoidea) from the early middle Eocene of Grube Messel (Germany). Palaeobio. Palaeoenv. 2017, 97, 807–832. [Google Scholar] [CrossRef]
- MacFadden, B.J. Fossil Horses: Systematics, Paleobiology, and Evolution of the Family Equidae; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Randau, M.; Goswani, A. Morphological modularity in the vertebral column of Felidae (Mammalia, Carnivora). BMC Evol. Biol. 2017, 17, 133. [Google Scholar] [CrossRef] [PubMed]
- Schlosser, G.; Wagner, G.P. Modularity in Development and Evolution; University of Chicago Press: Chicago, IL, USA, 2004. [Google Scholar]
- Figueirido, B.; Pérez-Ramos, A.; Martín-Serra, A. Intravertebral vs. intervertebral integration and modularity in the vertebral column of mammalian carnivorans. J. Anat. 2022, 4, 642–656. [Google Scholar] [CrossRef]
- Vidal, P.P.; Graf, W.; Berthoz, A. The orientation of the cervical vertebral column in unrestrained awake animals. Exp. Brain Res. 1986, 61, 549–559. [Google Scholar] [CrossRef]
- Graf, W.; de Waele, C.; Vidal, P.; Wang, D.; Evinger, C. The orientation of the cervical vertebral column in unrestrained awake animals. II. Movement strategies. Brain Behav. Evol. 1995, 45, 209–231. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, C.M.; Wright, L.; Kristoffersen, M.; Puchalski, S.M. Computed tomography and myelography of the equine cervical spine: 180 cases (2013–2018). Equine Vet. Educ. 2020, 9, 475–483. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species (n); Geological Period | Institute (n = 8) | Collection/Catalogue No. | Adult or Subadult |
|---|---|---|---|
| Equus sp. (4), | NHMB | MB. Ma. 45354 | Adult |
| MB. Ma. 24107 | Subadult | ||
| MB. Ma. 24410 | Adult | ||
| MB. Ma. 24463 | Adult | ||
| Equus mosbachensis (4), 80–600 kya | MMHBE | GO 374 3’ 8P 197 | Adult |
| Scho 13 II 4 95 690 22 14 | |||
| Scho 13 II 4 95 690 18 45 | |||
| Scho 13 II 4 96 692 27 | |||
| Equus sp. (1), 10–40 kya | Ko 100–108 | Subadult | |
| Equus occidentalis (20), 10–40 kya | TPMRLB | 2-T22, 8’9 | Adult |
| E-2, 9 | Subadult | ||
| 3 E2, 16 | Subadult (left side only) | ||
| 3, E-1, 17.5 | Adult | ||
| 3, E-2, 12 | |||
| 3, F4, 12 | Subadult | ||
| 3, F-4, 12 | Adult | ||
| 4 B2 M-19 | |||
| 4 B5, 20 | |||
| 4 C-2, 17–19 | |||
| 4 C-4 13–15 | |||
| 4 D2–4, 8,17 | |||
| 4 D2–99 | |||
| 4, C5, 17 246 | Subadult | ||
| 60 D-12, 14–16 | Adult | ||
| 4, G + H-2 + 3,8 | Subadult | ||
| 67 H-9, 15–18.5 | |||
| 77 E-9, 11 | Adult | ||
| Circle 61 | Subadult | ||
| GIZ, 14.5–16.5 | Adult (right side only) | ||
| Pliohippus sp. (1), | YHM | Mounted replica | Adult |
| Hyracotherium sp. (1), | |||
| Mesohippus sp. (1), | |||
| Hyracotherium angustidens (1), 55 mya | AMNH | 15428 | Adult (right side only) |
| Merychippus isonesus (1), 15–5 mya | ESP. 5263638 | ||
| Mesohippus bairdii (1), 37–32 mya | 1477 | Adult | |
| Miohippus obliquidens (1), 32–25 mya | 39115 | Adult (left side only) | |
| Pliohippus mirabilis (1), 12–6 mya | 60810 | ||
| Pliohippus pernix (1), 12–6 mya | 60803 | Adult | |
| Mesohippus sp. (cast) (1) | OUMNH | Mounted Replica | Adult |
| Pliohippus sp. (1) 12–6 mya | LACM | (CIT) 210 | Adult |
| Equus sp. (6) | LACM | (CIT) 138 120 | Subadult |
| (CIT) 138 138 | Adult (left side only) | ||
| (CIT) 138 699 | Adult | ||
| 192 L8 | Adult (right side only) | ||
| LACM San Josecito | (CIT) 192 | Adult (right side only) | |
| LACM Row XIX | (CIT) 192 (duplicate) | Adult (right side only) | |
| Equus simplicidens (15), 5.3–1.8 mya | USNM | USNM 13791 | Adult |
| USNM 12573 | |||
| USNM PAL 785552 | Subadult | ||
| USNM V 12575 | Adult | ||
| USNM PAL 785561 | |||
| USNM 785560 | |||
| USNM PAL 785553 | |||
| USNM 78559 | |||
| USNM 12155 | |||
| USNM 12580 | |||
| USNM PAL 785557 | |||
| USNM PAL 785556 | |||
| USNM PAL 785559 | |||
| USNM PAL 785555 | |||
| USNM PAL 785554 | |||
| Equus sp. (7) | UF | Leisey 1A UF 82283 | Adult |
| Leisey 1A UF 151092 | |||
| Leisey 1A UF 151020 | Adult (right side only) | ||
| UF 242520 | |||
| UF 242521 | Adult | ||
| Leisey 1A UF 151094 | Adult (Left side only) | ||
| UF 242522 | Adult (right side only) |
| Species (n); Author; Geological Period | Institute | Collection/Catalogue No. | Adult or Subadult |
|---|---|---|---|
| Equus africanus (1), | NBC | N.A.M. 28.-11-1972 | Adult |
| Equus hermionus (1), | No. 509 | ||
| Equus sp. (1) | Equus sp. mounted skeleton | ||
| Equus asinus (3), | ACEPT | Donald | Adult |
| Sebastian | |||
| Kam | |||
| Equus przewalskii (5), | ES | Heteni | |
| Rideg | Subadult | ||
| YHM | YHM—Live Pony Park | Adult | |
| NHMB | MB. Ma. 16464 | ||
| MB. Ma. 45373 | |||
| Equus quagga boehmi (1), | ES | Kimberley—Zoo bred |
| Species (n); Author; Geological Period | Institute | Collection/Catalogue No. | Adult or Subadult |
|---|---|---|---|
| Neohipparion whitneyi (1), (Gidley 1903) 13.6–4.9 mya [46] | AMNH | 9815 | Adult |
| Hyracotherium grangeri (1), (Gingerich 1889); 55 mya [47] | UM | 115547 | |
| Eurohippus messelensis (1), (Haupt 1925) 56–33.9 mya [48] | SMF | SMF ME 11034 |
| Species (n) and Geological Period | Institute (n = 8) | Collection Catalogue No. | Size of Convexity |
|---|---|---|---|
| Hyracotherium (1) | YHM | Mounted replica | Large |
| Hyracotherium (1) 55 mya | AMNH | 15428 | Large |
| Hyracotherium grangeri (1) 55 mya [47] | UM | 115547 | Large |
| Eurohippus messelensis (1) 56–33.9 mya [48] | SMF | SMF ME 11034 | * |
| Mesohippus (cast) (1) | OUMNH | Mounted Replica | Large |
| Mesohippus bairdii (1) 37–32 mya | AMNH | 1477 | Large |
| Mesohippus (1) | YHM | Mounted replica | Large |
| Miohippus doliquideus (1) 32–25 mya | AMNH | 39115 | Large |
| Merychippus isonesus (1) 15–5 mya | AMNH | ESP. 5263638 | Large |
| Neohipparion whitneyi (1) 13.6–4.9 mya [46] | AMNH | 9815 | Large ^ |
| Pliohippus mirabilis (1) 12–6 mya | AMNH | 60810 | Large |
| Pliohippus pernix (1) 12–6 mya | AMNH | 60803 | Large |
| Pliohippus (1) | YHM | Mounted replica | Large |
| Pliohippus (1) 12–6 mya | LACM | (CIT) 210 | Large |
| Equus simplicidens (15) 5.3–1.8 mya | USNM | USNM 13791 | Small |
| USNM 12573 | Small | ||
| USNM PAL 785552 | Small | ||
| USNM V 12575 | Small | ||
| USNM PAL 785561 | Small | ||
| USNM 785560 | Small | ||
| USNM PAL 785553 | Medium | ||
| USNM 785559 | Medium | ||
| USNM 12155 | Medium | ||
| USNM 12580 | Small | ||
| USNM PAL 785557 | Medium | ||
| USNM PAL 785556 | Small | ||
| USNM PAL 785559 | Small | ||
| USNM PAL 785555 | Medium | ||
| USNM PAL 785554 | Small | ||
| Equus occidentalis (20) 10–40 kya | TPMRLB | 2-T22, 8’9 | Small |
| E-2, 9 | Small | ||
| 3 E2, 16 | Small | ||
| 3, E-1, 17.5 | Medium | ||
| 3, E-2, 12 | Medium | ||
| 3, F4, 12 | Medium | ||
| 3, F-4, 12 | Small | ||
| 4 B2 M-19 | Small | ||
| 4 B5, 20 | Small | ||
| 4 C-2, 17–19 | Small | ||
| 4 C-4 13–15 | Small | ||
| 4 D2-4, 8, 17 | Small | ||
| 4 D2–99 | Small | ||
| 4, C5, 17 246 | Small | ||
| 60 D-12, 14–16 | Small | ||
| 4, G + H-2 + 3,8 | Small | ||
| 67 H-9, 15–18.5 | Small | ||
| 77 E-9, 11 | Medium | ||
| Circle 61 | Medium | ||
| GIZ, 14.5–16.5 | Small | ||
| Equus mosbachensis (4) 80–600 kya | MMHBE | GO 374 3’ 8 P 197 | Small |
| Scho 13 II 4 95 690 22 14 | Small | ||
| Scho 13 II 4 95 690 18 45 | Small | ||
| Scho 13 II 4 96 692 27 | Small | ||
| Equus sp. 10–40 kya (1) | Ko 100–108 | Small | |
| Equus sp. (4) | NHMB | MB. Ma. 24107 | Small |
| MB. Ma. 24410 | Medium | ||
| MB. Ma. 24463 | Medium | ||
| MB. Ma. 45354 | Medium | ||
| Equus sp. (6) | LACM | (CIT) 138 120 | Small |
| (CIT) 138 138 | Small | ||
| (CIT) 138 699 | Medium | ||
| 192 L8 | Medium | ||
| LACM San Josecito | (CIT) 192 | Small | |
| LACM Row XIX | (CIT) 192 (duplicate) | Small | |
| Equus sp. (7) | UF | Leisey 1A UF 82283 | Small |
| Leisey 1A UF 151092 | Medium | ||
| Leisey 1A 151120 | Medium | ||
| UF 242520 | Small | ||
| UF 242521 | Small | ||
| Leisey 1A UF 151094 | Medium | ||
| UF 242522 | Medium |
| Species (n) | Institute | Collection/Catalogue No. | Adult or Subadult |
|---|---|---|---|
| Equus africanus (1) | NBC | N.A.M. 28.-11-1972 | Small |
| Equus hermionus (1) | No. 509 | ||
| Equus sp. (1) | Equus sp. mounted skeleton | ||
| Equus asinus (3) | ACEPT | Donald | Small |
| Sebastian | |||
| Kam | |||
| Equus przewalskii (5) | ES | Heteni | |
| Rideg | Small | ||
| YHM | YHM—Live Pony Park | Small | |
| NHMB | MB. Ma. 16464 | ||
| MB. Ma. 45373 | |||
| Equus quagga boehmi (1) | ES | Kimberley |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
May-Davis, S.; Hunter, R.; White, R. Morphology of the Ventral Process of the Sixth Cervical Vertebra in Extinct and Extant Equus: Functional Implications. Animals 2023, 13, 1672. https://doi.org/10.3390/ani13101672
May-Davis S, Hunter R, White R. Morphology of the Ventral Process of the Sixth Cervical Vertebra in Extinct and Extant Equus: Functional Implications. Animals. 2023; 13(10):1672. https://doi.org/10.3390/ani13101672
Chicago/Turabian StyleMay-Davis, Sharon, Robert Hunter, and Richard White. 2023. "Morphology of the Ventral Process of the Sixth Cervical Vertebra in Extinct and Extant Equus: Functional Implications" Animals 13, no. 10: 1672. https://doi.org/10.3390/ani13101672
APA StyleMay-Davis, S., Hunter, R., & White, R. (2023). Morphology of the Ventral Process of the Sixth Cervical Vertebra in Extinct and Extant Equus: Functional Implications. Animals, 13(10), 1672. https://doi.org/10.3390/ani13101672