
Integrated Metabolomic and Transcriptomic Analysis Reveals Potential Gut-Liver Crosstalks in the Lipogenesis of Chicken
 ,
, 
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sample Preparation
2.3. Detection of Lipogenic Indicators
2.4. Metabolomics Analysis
2.5. Hepatic Transcriptome Analysis
2.6. Correlation Analysis
3. Results
3.1. HFD Induced Enhanced Lipogenesis and Excessive Fat Deposition
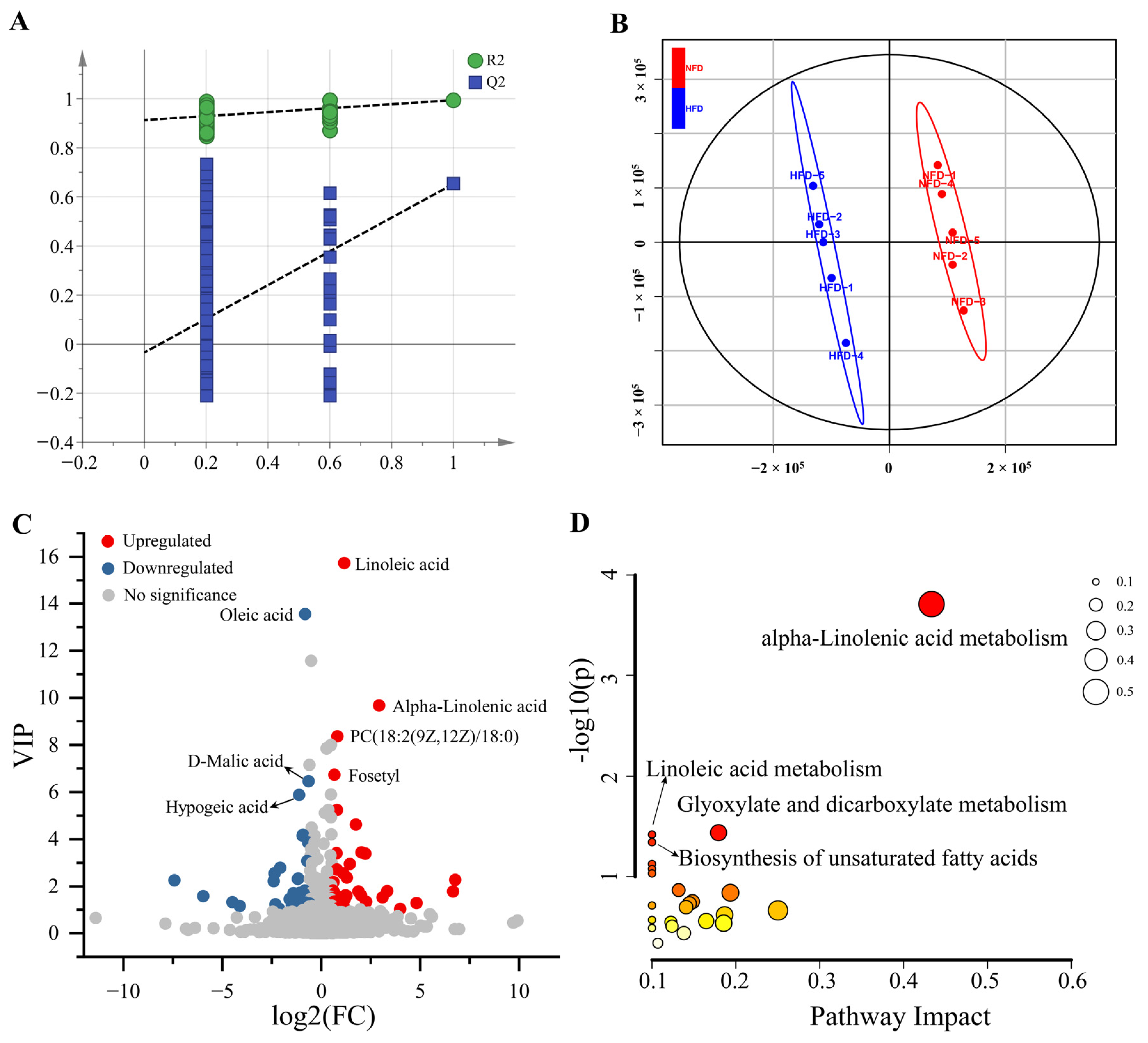
3.2. Effects of HFD on Cecal Metabolomics
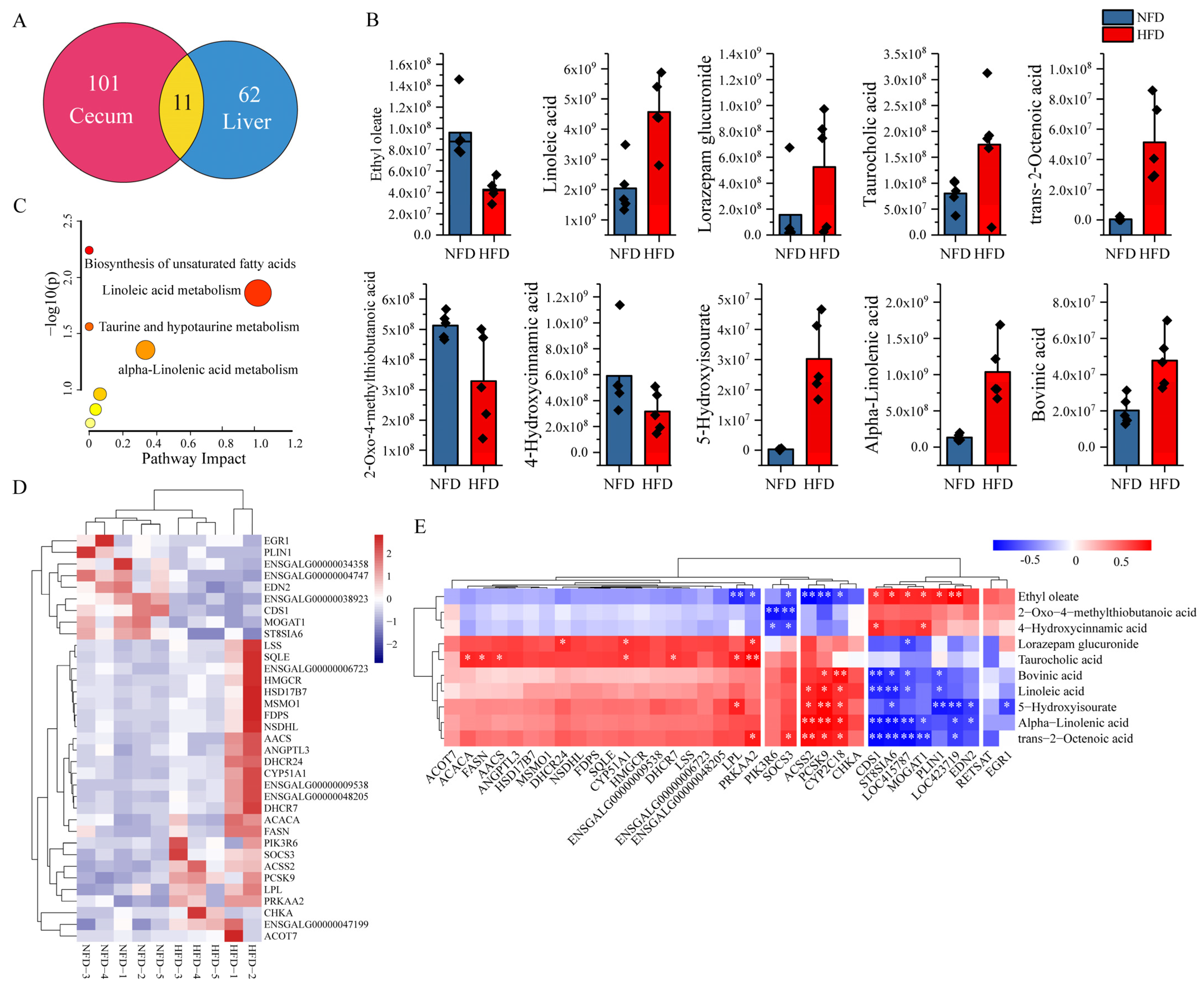
3.3. Effects of HFD on Hepatic Metabolomics
3.4. Effects of HFD on Hepatic Transcriptome
3.5. Identification of Potential Lipogenesis-Related Gut–Liver Crosstalks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, T.; Lu, H.; Wang, L.; Yin, M.; Yang, L. Specific expression pattern of IMP metabolism related-genes in chicken muscle between cage and free range conditions. PLoS ONE 2018, 13, e0201736. [Google Scholar] [CrossRef]
- Tian, W.; Hao, X.; Nie, R.; Ling, Y.; Zhang, B.; Zhang, H.; Wu, C. Integrative analysis of miRNA and mRNA profiles reveals that gga-miR-106-5p inhibits adipogenesis by targeting the KLF15 gene in chickens. J. Anim. Sci. Biotechnol. 2022, 13, 81. [Google Scholar] [CrossRef]
- Nematbakhsh, S.; Pei Pei, C.; Selamat, J.; Nordin, N.; Idris, L.H.; Abdull Razis, A.F. Molecular Regulation of Lipogenesis, Adipogenesis and Fat Deposition in Chicken. Genes 2021, 12, 414. [Google Scholar] [CrossRef]
- Ma, Z.; Li, H.; Zheng, H.; Jiang, K.; Yan, F.; Tian, Y.; Kang, X.; Wang, Y.; Liu, X. Hepatic ELOVL6 mRNA is regulated by the gga-miR-22-3p in egg-laying hen. Gene 2017, 623, 72–79. [Google Scholar] [CrossRef]
- Cui, H.; Zheng, M.; Zhao, G.; Liu, R.; Wen, J. Identification of differentially expressed genes and pathways for intramuscular fat metabolism between breast and thigh tissues of chickens. BMC Genomics 2018, 19, 55. [Google Scholar] [CrossRef]
- Ding, J.H.; Jin, Z.; Yang, X.X.; Lou, J.; Shan, W.X.; Hu, Y.X.; Du, Q.; Liao, Q.S.; Xie, R.; Xu, J.Y. Role of gut microbiota via the gut-liver-brain axis in digestive diseases. World J. Gastroenterol. 2020, 26, 6141–6162. [Google Scholar] [CrossRef]
- Wahlström, A.; Sayin, S.I.; Marschall, H.U.; Bäckhed, F. Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab. 2016, 24, 41–50. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef]
- Gu, T.; Duan, M.; Zhang, R.; Zeng, T.; Xu, W.; Feng, W.; Jiang, C.; Tian, Y.; Chen, L.; Lu, L. Probiotic Fermented Feed Alleviates Liver Fat Deposition in Shaoxing Ducks via Modulating Gut Microbiota. Front. Microbiol. 2022, 13, 928670. [Google Scholar] [CrossRef]
- Saeedi, B.J.; Liu, K.H.; Owens, J.A.; Hunter-Chang, S.; Camacho, M.C.; Eboka, R.U.; Chandrasekharan, B.; Baker, N.F.; Darby, T.M.; Robinson, B.S.; et al. Gut-Resident Lactobacilli Activate Hepatic Nrf2 and Protect against Oxidative Liver Injury. Cell Metab. 2020, 31, 956–968.e5. [Google Scholar] [CrossRef]
- Yang, H.; Pan, R.; Wang, J.; Zheng, L.; Li, Z.; Guo, Q.; Wang, C. Modulation of the Gut Microbiota and Liver Transcriptome by Red Yeast Rice and Monascus Pigment Fermented by Purple Monascus SHM1105 in Rats Fed with a High-Fat Diet. Front. Pharmacol. 2020, 11, 599760. [Google Scholar] [CrossRef] [PubMed]
- Mayneris-Perxachs, J.; Cardellini, M.; Hoyles, L.; Latorre, J.; Davato, F.; Moreno-Navarrete, J.M.; Arnoriaga-Rodríguez, M.; Serino, M.; Abbott, J.; Barton, R.H.; et al. Iron status influences non-alcoholic fatty liver disease in obesity through the gut microbiome. Microbiome 2021, 9, 104. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Adolph, T.E.; Trauner, M. Gut-liver axis: Pathophysiological concepts and clinical implications. Cell Metab. 2022, 34, 1700–1718. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Ding, H.; Chen, L.; Lin, Y.; Gong, Y.; Pan, Z.; Zhang, G.; Xie, K.; Dai, G.; Wang, J. Antibiotic-Induced Dysbiosis of Microbiota Promotes Chicken Lipogenesis by Altering Metabolomics in the Cecum. Metabolites 2021, 11, 487. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.; Sun, Q.; Ma, Z.; Peng, J.; Zhang, M.; Wang, C.; Zhang, X.; Yan, X.; Chang, L.; Hou, X.; et al. Seabuckthorn polysaccharide ameliorates high-fat diet-induced obesity by gut microbiota-SCFAs-liver axis. Food Funct. 2022, 13, 2925–2937. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Guo, R.; Pei, L.; Lai, S.; Li, J.; Yin, Y.; Xu, T.; Yang, W.; Song, Q.; Han, Q.; et al. N-Acetylcysteine alleviates high fat diet-induced hepatic steatosis and liver injury via regulating the intestinal microecology in mice. Food Funct. 2022, 13, 3368–3380. [Google Scholar] [CrossRef]
- Rao, Y.; Kuang, Z.; Li, C.; Guo, S.; Xu, Y.; Zhao, D.; Hu, Y.; Song, B.; Jiang, Z.; Ge, Z.; et al. Gut Akkermansia muciniphila ameliorates metabolic dysfunction-associated fatty liver disease by regulating the metabolism of L-aspartate via gut-liver axis. Gut Microbes 2021, 13, 1927633. [Google Scholar] [CrossRef]
- Daniel, N.; Genua, F.; Jenab, M.; Mayén, A.L.; Chrysovalantou Chatziioannou, A.; Keski-Rahkonen, P.; Hughes, D.J. The role of the gut microbiome in the development of hepatobiliary cancers. Hepatology 2023. [Google Scholar] [CrossRef]
- Xu, H.; Luo, J.; Ma, G.; Zhang, X.; Yao, D.; Li, M.; Loor, J.J. Acyl-CoA synthetase short-chain family member 2 (ACSS2) is regulated by SREBP-1 and plays a role in fatty acid synthesis in caprine mammary epithelial cells. J. Cell. Physiol. 2018, 233, 1005–1016. [Google Scholar] [CrossRef]
- Huang, Z.; Zhang, M.; Plec, A.A.; Estill, S.J.; Cai, L.; Repa, J.J.; McKnight, S.L.; Tu, B.P. ACSS2 promotes systemic fat storage and utilization through selective regulation of genes involved in lipid metabolism. Proc. Natl. Acad. Sci. USA 2018, 115, E9499–E9506. [Google Scholar] [CrossRef]
- Momtazi-Borojeni, A.A.; Banach, M.; Ruscica, M.; Sahebkar, A. The role of PCSK9 in NAFLD/NASH and therapeutic implications of PCSK9 inhibition. Expert Rev. Clin. Pharmacol. 2022, 15, 1199–1208. [Google Scholar] [CrossRef] [PubMed]
- Lebeau, P.F.; Byun, J.H.; Platko, K.; MacDonald, M.E.; Poon, S.V.; Faiyaz, M.; Seidah, N.G.; Austin, R.C. Diet-induced hepatic steatosis abrogates cell-surface LDLR by inducing de novo PCSK9 expression in mice. J. Biol. Chem. 2019, 294, 9037–9047. [Google Scholar] [CrossRef] [PubMed]
- Tavori, H.; Giunzioni, I.; Predazzi, I.M.; Plubell, D.; Shivinsky, A.; Miles, J.; Devay, R.M.; Liang, H.; Rashid, S.; Linton, M.F.; et al. Human PCSK9 promotes hepatic lipogenesis and atherosclerosis development via apoE- and LDLR-mediated mechanisms. Cardiovasc. Res. 2016, 110, 268–278. [Google Scholar] [CrossRef]
- Herbert, B.; Patel, D.; Waddington, S.N.; Eden, E.R.; McAleenan, A.; Sun, X.M.; Soutar, A.K. Increased secretion of lipoproteins in transgenic mice expressing human D374Y PCSK9 under physiological genetic control. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1333–1339. [Google Scholar] [CrossRef]
- Sun, H.; Samarghandi, A.; Zhang, N.; Yao, Z.; Xiong, M.; Teng, B.B. Proprotein convertase subtilisin/kexin type 9 interacts with apolipoprotein B and prevents its intracellular degradation, irrespective of the low-density lipoprotein receptor. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1585–1595. [Google Scholar] [CrossRef]
- Watanabe, K.P.; Kawai, Y.K.; Nakayama, S.M.; Ikenaka, Y.; Mizukawa, H.; Takaesu, N.; Ito, M.; Ikushiro, S.I.; Sakaki, T.; Ishizuka, M. Partial cloning of CYP2C23a genes and hepatic protein expression in eight representative avian species. J. Vet. Pharmacol. Ther. 2015, 38, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Woodfint, R.M.; Lee, J.; Wu, H.; Ma, J.; Suh, Y.; Hwang, S.; Cressman, M.; Lee, K. Comparative identification, nutritional, and physiological regulation of chicken liver-enriched genes. Poult. Sci. 2019, 98, 3007–3013. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Mak, H.Y.; Lukmantara, I.; Li, Y.E.; Hoehn, K.L.; Huang, X.; Du, X.; Yang, H. CDP-DAG synthase 1 and 2 regulate lipid droplet growth through distinct mechanisms. J. Biol. Chem. 2019, 294, 16740–16755. [Google Scholar] [CrossRef]
- Qi, Y.; Kapterian, T.S.; Du, X.; Ma, Q.; Fei, W.; Zhang, Y.; Huang, X.; Dawes, I.W.; Yang, H. CDP-diacylglycerol synthases regulate the growth of lipid droplets and adipocyte development. J. Lipid Res. 2016, 57, 767–780. [Google Scholar] [CrossRef]
- Singer, J.M.; Shew, T.M.; Ferguson, D.; Renkemeyer, M.K.; Pietka, T.A.; Hall, A.M.; Finck, B.N.; Lutkewitte, A.J. Monoacylglycerol O-acyltransferase 1 lowers adipocyte differentiation capacity in vitro but does not affect adiposity in mice. Obesity 2022, 30, 2122–2133. [Google Scholar] [CrossRef]
- Lutkewitte, A.J.; Singer, J.M.; Shew, T.M.; Martino, M.R.; Hall, A.M.; He, M.; Finck, B.N. Multiple antisense oligonucleotides targeted against monoacylglycerol acyltransferase 1 (Mogat1) improve glucose metabolism independently of Mogat1. Mol. Metab. 2021, 49, 101204. [Google Scholar] [CrossRef] [PubMed]
- Zhai, G.; Pang, Y.; Zou, Y.; Wang, X.; Liu, J.; Zhang, Q.; Cao, Z.; Wang, N.; Li, H.; Wang, Y. Effects of PLIN1 Gene Knockout on the Proliferation, Apoptosis, Differentiation and Lipolysis of Chicken Preadipocytes. Animals 2022, 13, 92. [Google Scholar] [CrossRef] [PubMed]
- Lou, X.; Zhou, X.; Li, H.; Lu, X.; Bao, X.; Yang, K.; Liao, X.; Chen, H.; Fang, H.; Yang, Y.; et al. Biallelic Mutations in ACACA Cause a Disruption in Lipid Homeostasis That Is Associated with Global Developmental Delay, Microcephaly, and Dysmorphic Facial Features. Front. Cell Dev. Biol. 2021, 9, 618492. [Google Scholar] [CrossRef] [PubMed]
- Moioli, B.; Scatà, M.C.; De Matteis, G.; Annicchiarico, G.; Catillo, G.; Napolitano, F. The ACACA gene is a potential candidate gene for fat content in sheep milk. Anim. Genet. 2013, 44, 601–603. [Google Scholar] [CrossRef] [PubMed]
- Stachowiak, M.; Nowacka-Woszuk, J.; Szydlowski, M.; Switonski, M. The ACACA and SREBF1 genes are promising markers for pig carcass and performance traits, but not for fatty acid content in the longissimus dorsi muscle and adipose tissue. Meat Sci. 2013, 95, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Song, Y.; Shi, Q.; Fu, L. Research progress on FASN and MGLL in the regulation of abnormal lipid metabolism and the relationship between tumor invasion and metastasis. Front. Med. 2021, 15, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, S.; Ikeda, Y.; Yamasaki, M.; Fukui, T. The role of acetoacetyl-CoA synthetase, a ketone body-utilizing enzyme, in 3T3-L1 adipocyte differentiation. Biol. Pharm. Bull. 2012, 35, 1980–1985. [Google Scholar] [CrossRef] [PubMed]
- Tsai, M.T.; Chen, Y.J.; Chen, C.Y.; Tsai, M.H.; Han, C.L.; Chen, Y.J.; Mersmann, H.J.; Ding, S.T. Identification of Potential Plasma Biomarkers for Non-alcoholic Fatty Liver Disease by Integrating Transcriptomics and Proteomics in Laying Hens. J. Nutr. 2017, 147, 293–303. [Google Scholar] [CrossRef]
- Bartelt, A.; Weigelt, C.; Cherradi, M.L.; Niemeier, A.; Tödter, K.; Heeren, J.; Scheja, L. Effects of adipocyte lipoprotein lipase on de novo lipogenesis and white adipose tissue browning. Biochim. Biophys. Acta 2013, 1831, 934–942. [Google Scholar] [CrossRef]
- Gao, J.; Ding, G.; Li, Q.; Gong, L.; Huang, J.; Sang, Y. Tibet kefir milk decreases fat deposition by regulating the gut microbiota and gene expression of Lpl and Angptl4 in high fat diet-fed rats. Food Res. Int. 2019, 121, 278–287. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO ID | Description | Gene Number | Q Value |
|---|---|---|---|
| GO:0016125 | sterol metabolic process | 12 | 3.14 × 10−6 |
| GO:0008203 | cholesterol metabolic process | 10 | 4.35 × 10−5 |
| GO:0016126 | sterol biosynthetic process | 8 | 4.35 × 10−5 |
| GO:0008610 | lipid biosynthetic process | 23 | 4.47 × 10−5 |
| GO:1902652 | secondary alcohol metabolic process | 10 | 5.40 × 10−5 |
| GO:0006629 | lipid metabolic process | 35 | 2.98 × 10−4 |
| GO:0055114 | oxidation-reduction process | 32 | 2.98 × 10−4 |
| GO:0008202 | steroid metabolic process | 13 | 3.85 × 10−4 |
| GO:0006694 | steroid biosynthetic process | 10 | 4.77 × 10−4 |
| GO:0006695 | cholesterol biosynthetic process | 6 | 5.24 × 10−4 |
| GO:1902653 | secondary alcohol biosynthetic process | 6 | 1.04 × 10−3 |
| GO:0006066 | alcohol metabolic process | 15 | 1.67 × 10−3 |
| GO:0044283 | small molecule biosynthetic process | 19 | 2.37 × 10−3 |
| GO:1901615 | organic hydroxy compound metabolic process | 17 | 6.11 × 10−3 |
| GO:0044711 | single-organism biosynthetic process | 36 | 6.11 × 10−3 |
| GO:0042742 | defense response to bacterium | 9 | 9.74 × 10−3 |
| GO:0046165 | alcohol biosynthetic process | 9 | 2.36 × 10−2 |
| GO:0034128 | negative regulation of MyD88-independent Toll-like receptor signaling pathway | 2 | 4.62 × 10−2 |
| GO:0071615 | oxidative deethylation | 2 | 4.62 × 10−2 |
| Metabolites | Cecum | Liver | ||
|---|---|---|---|---|
| Fold Change | VIP | Fold Change | VIP | |
| 2-oxo-4-methylthiobutanoic acid | 5.632986445 ↓ | 1.34903 | 1.562114 ↓ | 3.84847 |
| 4-hydroxycinnamic acid | 4.022521132 ↓ | 1.31592 | 1.874103 ↓ | 4.17537 |
| 5-hydroxyisourate | 9.492578339 ↑ | 1.68904 | 101.3498 ↑ | 1.77908 |
| alpha-linolenic acid | 22.38871776 ↑ | 5.35261 | 7.669327 ↑ | 9.67412 |
| bovinic acid | 7.363542529 ↑ | 1.26022 | 2.359367 ↑ | 1.61691 |
| ethyl oleate | 1.581726323 ↓ | 1.64936 | 2.253551 ↓ | 2.29809 |
| linoleic acid | 4.777977301 ↑ | 11.6529 | 2.239847 ↑ | 15.7242 |
| lorazepam glucuronide | 13.1473488 ↑ | 5.31738 | 3.366851 ↑ | 4.61575 |
| taurocholic acid | 10.84230807 ↑ | 1.31562 | 2.177994 ↑ | 2.53774 |
| trans-2-octenoic acid | 718.8764339 ↑ | 2.81436 | 109.9878 ↑ | 2.26858 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Chen, W.; Ding, H.; Zhang, G.; Xie, K.; Zhang, T. Integrated Metabolomic and Transcriptomic Analysis Reveals Potential Gut-Liver Crosstalks in the Lipogenesis of Chicken. Animals 2023, 13, 1659. https://doi.org/10.3390/ani13101659
Chen C, Chen W, Ding H, Zhang G, Xie K, Zhang T. Integrated Metabolomic and Transcriptomic Analysis Reveals Potential Gut-Liver Crosstalks in the Lipogenesis of Chicken. Animals. 2023; 13(10):1659. https://doi.org/10.3390/ani13101659
Chicago/Turabian StyleChen, Can, Weilin Chen, Hao Ding, Genxi Zhang, Kaizhou Xie, and Tao Zhang. 2023. "Integrated Metabolomic and Transcriptomic Analysis Reveals Potential Gut-Liver Crosstalks in the Lipogenesis of Chicken" Animals 13, no. 10: 1659. https://doi.org/10.3390/ani13101659
APA StyleChen, C., Chen, W., Ding, H., Zhang, G., Xie, K., & Zhang, T. (2023). Integrated Metabolomic and Transcriptomic Analysis Reveals Potential Gut-Liver Crosstalks in the Lipogenesis of Chicken. Animals, 13(10), 1659. https://doi.org/10.3390/ani13101659