
Vascular Distribution and Expression Patterns of Angiogenic Factors in Caruncle during the Early Stage of Pregnancy in Goats (Capra hircus)


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Caruncle Collection
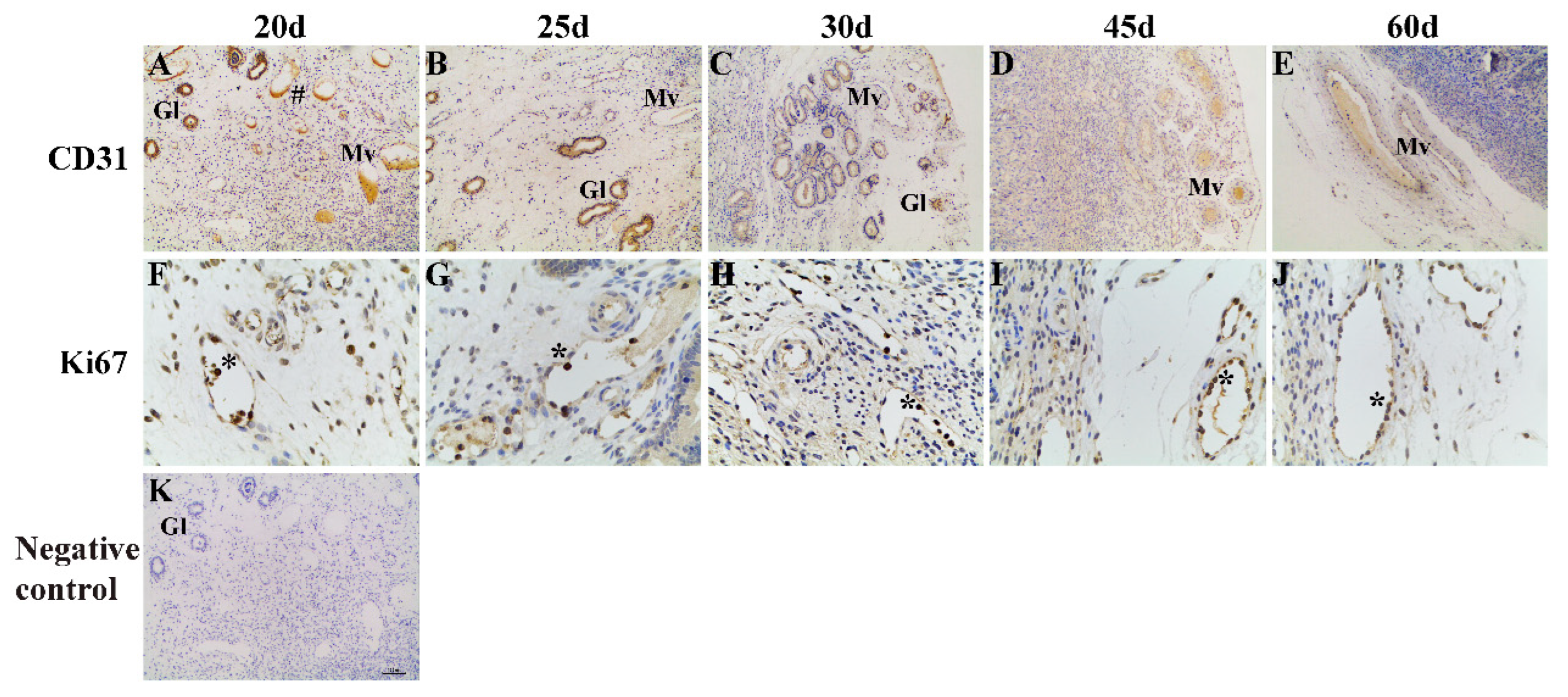
2.2. Immunohistochemistry and Image Analysis
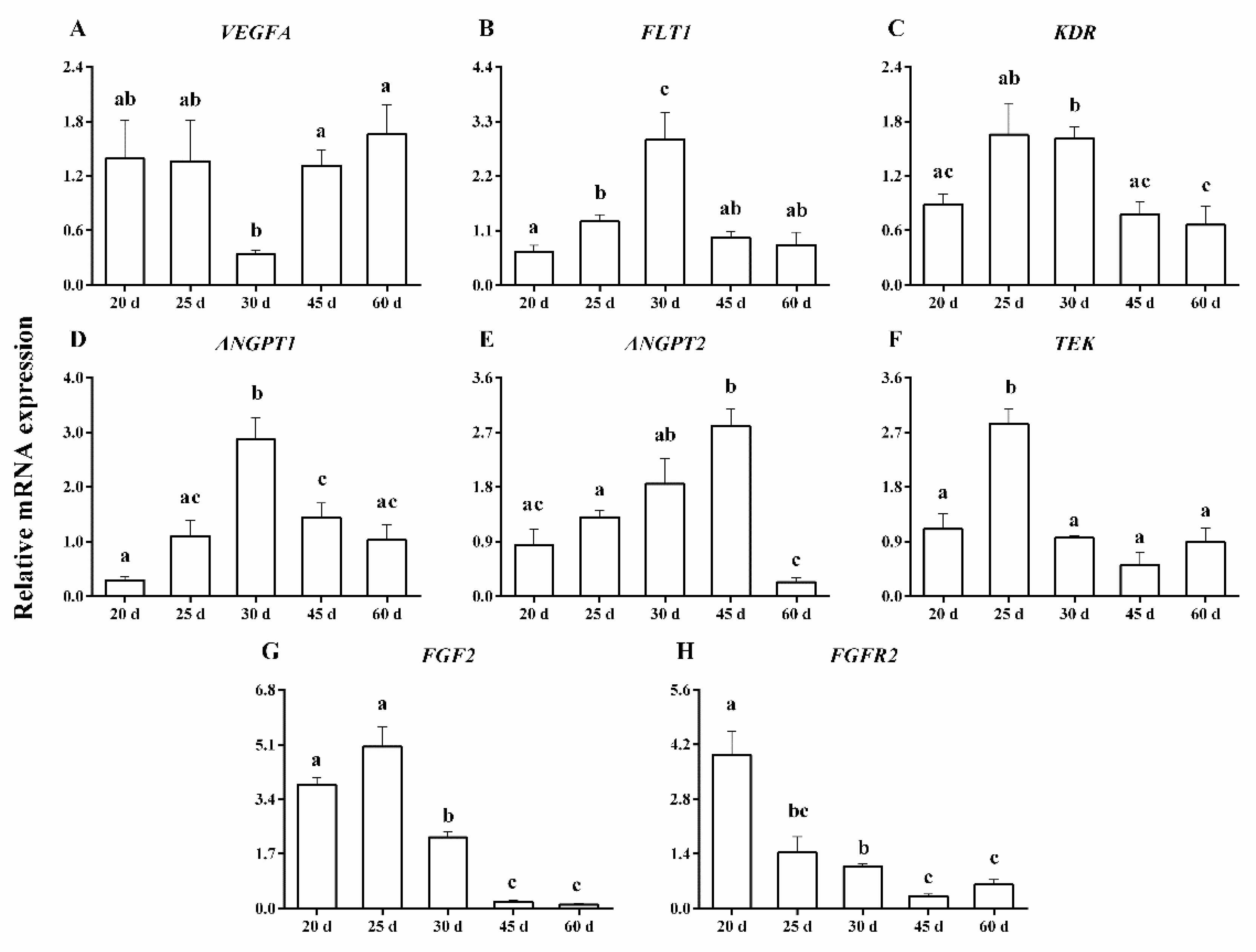
2.3. RNA Isolation and Quantitative Real-Time RT-PCR for mRNA Expression
2.4. ANGPT2 Methylation Assay
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Borowicz, P.P.; Arnold, D.R.; Johnson, M.L. Grazul-Bilska AT, Redmer DA, Reynolds LP. Placental Growth Throughout the Last Two Thirds of Pregnancy in Sheep, Vascular Development and Angiogenic Factor Expression. Biol. Reprod. 2007, 76, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Cui, Y.; Yang, B.; Fan, J.; Zhao, Z.; Yu, S. Morphometric Analysis of Yak Placentomes During Gestation. Anat. Rec. 2010, 293, 1873–1879. [Google Scholar] [CrossRef] [PubMed]
- Osol, G.; Moore, L.G. Maternal Uterine Vascular Remodeling During Pregnancy. Microcirculation 2014, 21, 38–47. [Google Scholar] [CrossRef]
- Demir, R.; Seval, Y.; Huppertz, B. Vasculogenesis and angiogenesis in the early human placenta. Acta Histochem. 2007, 109, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Grazul-Bilska, A.T.; Borowicz, P.P.; Johnson, M.L.; Minten, M.A.; Bilski, J.J.; Wroblewski, R.; Redmer, D.A.; Reynolds, L.P. Placental development during early pregnancy in sheep, Vascular growth and expression of angiogenic factors in maternal placenta. Reproduction 2010, 140, 165–174. [Google Scholar] [CrossRef]
- Goodger, A.M.; Rogers, P.A. Blood vessel growth and endothelial cell density in rat endometrium. J. Reprod. Fertil. 1995, 105, 259–261. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bairagi, S.; Quinn, K.E.; Crane, A.R.; Ashley, R.L.; Borowicz, P.P.; Caton, J.S.; Redden, R.R.; Grazul-Bilska, A.T.; Reynolds, L.P. Maternal environment and placental vascularization in small ruminants. Theriogenology 2016, 86, 288–305. [Google Scholar] [CrossRef]
- Gale, N.W.; Thurston, G.; Hackett, S.F.; Renard, R.; Wang, Q.; McClain, J.; Martin, C.; Witte, C.; Witte, M.H.; Jackson, D.; et al. Angiopoietin-2 is required for postnatal angiogenesis and lymphatic patterning, and only the latter role is rescued by angiopoietin-1. Dev. Cell. 2002, 3, 411–423. [Google Scholar] [CrossRef]
- Augustin, H.G.; Koh, G.Y.; Thurston, G.; Alitalo, K. Control of vascular morphogenesis and homeostasis through the angiopoietin-Tie system. Nat. Rev. Mol. Cell Biol. 2009, 10, 165–177. [Google Scholar] [CrossRef]
- Fearnley, G.W.; Smith, G.A.; Odell, A.F.; Latham, A.M.; Wheatcroft, S.B.; Harrison, M.A.; Tomlinson, D.C.; Ponnambalam, S. Vascular endothelial growth factor A-stimulated signaling from endosomes in primary endothelial cells. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2014; pp. 265–292. [Google Scholar]
- Wang, Q.; Lash, G.E. Angiopoietin 2 in placentation and tumor biology, The yin and yang of vascular biology. Placenta 2017, 56, 73–78. [Google Scholar] [CrossRef]
- Yancopoulos, G.D.; Davis, S.; Gale, N.W.; Rudge, J.S.; Wiegand, S.J.; Holash, J. Vascular-specific growth factors and blood vessel formation. Nature 2000, 407, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Bhat, A.; Woodnutt, G.; Lappe, R. Targeting the ANGPT-TIE2 pathway in malignancy. Nat. Rev. Cancer 2010, 10, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Saharinen, P.; Leppänen, V.M.; Alitalo, K. SnapShot, Angiopoietins and Their Functions. Cell 2017, 171, 724–724.e1. [Google Scholar] [CrossRef] [PubMed]
- Moss, A. The angiopoietin, Tie 2 interaction, a potential target for future therapies in human vascular disease. Cytokine Growth Factor Rev. 2013, 24, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Mata-Greenwood, E.; Liao, W.X.; Zheng, J.; Chen, D.B. Differential activation of multiple signaling pathways dictates eNOS upregulation by FGF2 but not VEGF in placental artery endothelial cells. Placenta 2008, 29, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wen, Y.; Song, Y.; Wang, K.; Chen, D.B.; Magness, R.R. Activation of multiple signaling pathways is critical for fibroblast growth factor 2- and vascular endothelial growth factor-stimulated ovine fetoplacental endothelial cell proliferation. Biol. Reprod. 2008, 78, 143–150. [Google Scholar] [CrossRef]
- Feng, L.; Liao, W.X.; Luo, Q.; Zhang, H.H.; Wang, W.; Zheng, J.; Chen, D.B. Caveolin-1 orchestrates fibroblast growth factor 2 signaling control of angiogenesis in placental artery endothelial cell caveolae. J. Cell Physiol. 2012, 227, 2480–2491. [Google Scholar] [CrossRef]
- Liu, Y.X.; Gao, F.; Wei, P.; Chen, X.L.; Gao, H.J.; Zou, R.J.; Siao, L.J.; Xu, F.H.; Feng, Q.; Liu, K.; et al. Involvement of molecules related to angiogenesis, proteolysis and apoptosis in implantation in rhesus monkey and mouse. Contraception 2005, 71, 249–262. [Google Scholar] [CrossRef]
- Grazul-Bilska, A.T.; Johnson, M.L.; Borowicz, P.P.; Minten, M.; Bilski, J.J.; Wroblewski, R.; Velimirovich, M.; Coupe, L.R.; Redmer, D.A.; Reynolds, L.P. Placental development during early pregnancy in sheep, cell proliferation, global methylation, and angiogenesis in the fetal placenta. Reproduction 2011, 141, 529–540. [Google Scholar] [CrossRef]
- Januar, V.; Desoye, G.; Novakovic, B.; Cvitic, S.; Saffery, R. Epigenetic regulation of human placental function and pregnancy outcome, considerations for causal inference. Am. J. Obstet. Gynecol. 2015, 213, S182–S196. [Google Scholar] [CrossRef]
- Han, Y.; Ouyang, H.; Che, D.; Chen, X.; Huang, Y.; Elfaki elhag, I.A.; Pang, D.; Li, Z. Elevated expression of vascular endothelial growth factor (VEGF) 120 in parthenogenetic porcine placentas. Biotechnol. Lett. 2014, 36, 913–917. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Liu, Z.; Han, Y.; Gao, W.; Hao, J.; Zhong, X.; Wang, D.; Li, Z. DNA methylation-mediated silencing of FLT1 in parthenogenetic porcine placentas. Placenta 2017, 58, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Correia-da-Silva, G.; Bell, S.C.; Pringle, J.H.; Teixeira, N.A. Patterns of Uterine Cellular Proliferation and Apoptosis in the Implantation Site of the Rat during Pregnancy. Placenta 2004, 25, 538–547. [Google Scholar] [CrossRef]
- Rowe, A.J.; Wulff, C.; Fraser, H.M. Angiogenesis and microvascular development in the marmoset (Callithrix jacchus) endometrium during early pregnancy. Reproduction 2004, 128, 107–116. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kar, M.; Ghosh, D.; Sengupta, J. Histochemical and morphological examination of proliferation and apoptosis in human first trimester villous trophoblast. Hum. Reprod. 2007, 22, 2814–2823. [Google Scholar] [CrossRef] [PubMed]
- Sanchis, E.G.; Cristofolini, A.L.; Fiorimanti, M.R.; Barbeito, C.G.; Merkis, C.I. Apoptosis and cell proliferation in porcine placental vascularization. Anim. Reprod. Sci. 2017, 184, 20–28. [Google Scholar] [CrossRef]
- Tang, Y.; Chen, Q.; Luo, N.; Tan, X.; Xu, H.; Zhao, Y. Mitochondrial related genes expression in placental tissues of different gestational stages in Dazu black goat. Chin. J. Vet. Sci. 2018, 38, 2193–2200. (In Chinese) [Google Scholar]
- Otrock, Z.K.; Makarem, J.A.; Shamseddine, A.I. Vascular endothelial growth factor family of ligands and receptors: Review. Blood Cells Mol. Dis. 2007, 38, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Vrachnis, N.; Kalampokas, E.; Sifakis, S.; Vitoratos, N.; Kalampokas, T.; Botsis, D.; Iliodromiti, Z. Placental growth factor (PlGF), a key to optimizing fetal growth. J. Matern. Fetal Neonatal Med. 2013, 26, 995–1002. [Google Scholar] [CrossRef]
- Kappou, D.; Sifakis, S.; Konstantinidou, A.; Papantoniou, N.; Spandidos, D.A. Role of the angiopoietin/Tie system in pregnancy (Review). Exp. Ther. Med. 2015, 9, 1091–1096. [Google Scholar] [CrossRef] [PubMed]
- Edwards, A.K.; van den Heuvel, M.J.; Wessels, J.M.; Lamarre, J.; Croy, B.A.; Tayade, C. Expression of angiogenic basic fibroblast growth factor, platelet derived growth factor, thrombospondin-1 and their receptors at the porcine maternal-fetal interface. Reprod. Biol. Endocrinol. 2011, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Fiorimanti, M.R.; Rabaglino, M.B.; Cristofolini, A.L.; Merkis, C.I. Immunohistochemical determination of Ang-1, Ang-2 and Tie-2 in placentas of sows at 30, 60 and 114 days of gestation and validation through a bioinformatic approach. Anim. Reprod. Sci. 2018, 195, 242–250. [Google Scholar] [CrossRef] [PubMed]
- Fagiani, E.; Christofori, G. Angiopoietins in angiogenesis. Cancer Lett. 2013, 328, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Welter, H.; Wollenhaupt, K.; Einspanier, R. Developmental and hormonal regulated gene expression of fibroblast growth factor 2 (FGF-2) and its receptors in porcine endometrium. J. Steroid. Biochem. Mol. Biol. 2004, 88, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Presta, M.; Dell’Era, P.; Mitola, S.; Moroni, E.; Ronca, R.; Rusnati, M. Fibroblast growth factor/fibroblast growth factor receptor system in angiogenesis. Cytokine Growth Factor Rev. 2005, 16, 159–178. [Google Scholar] [CrossRef]
- Pfarrer, C.; Weise, S.; Berisha, B.; Schams, D.; Leiser, R.; Hoffmann, B.; Schuler, G. Fibroblast Growth Factor (FGF)-1, FGF2, FGF7 and FGF Receptors are Uniformly Expressed in Trophoblast Giant Cells during Restricted Trophoblast Invasion in Cows. Placenta 2006, 27, 758–770. [Google Scholar] [CrossRef]
- Ozawa, M.; Yang, Q.E.; Ealy, A.D. The expression of fibroblast growth factor receptors during early bovine conceptus development and pharmacological analysis of their actions on trophoblast growth in vitro. Reproduction 2013, 145, 191–201. [Google Scholar] [CrossRef]
- Kunath, T.; Yamanaka, Y.; Detmar, J.; MacPhee, D.; Caniggia, I.; Rossant, J.; Jurisicova, A. Developmental differences in the expression of FGF receptors between human and mouse embryos. Placenta 2014, 35, 1079–1088. [Google Scholar] [CrossRef]
- Lins, R.J.; Mitchell, M.D. Novel Insights into the Control of Human Pregnancy: Potential Role(s) for Epigenetic Regulation. Reprod. Biol. Insights 2008, 2008, 3–8. [Google Scholar]
- Sundrani, D.P.; Reddy, U.S.; Joshi, A.A.; Mehendale, S.S.; Chavan-Gautam, P.M.; Hardikar, A.A.; Chandak, G.R.; Joshi, S.R. Differential placental methylation and expression of VEGF, FLT-1 and KDR genes in human term and preterm preeclampsia. Clin. Epigenetics 2013, 5, 6. [Google Scholar] [CrossRef]
- Martinelli, S.; Kanduri, M.; Maffei, R.; Fiorcari, S.; Bulgarelli, J.; Marasca, R.; Rosenquist, R. ANGPT2 promoter methylation is strongly associated with gene expression and prognosis in chronic lymphocytic leukemia. Epigenetics 2013, 8, 720–729. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. | Primer Sequence (5′→3′) | Product Length (bp) |
|---|---|---|---|
| VEGFA | XM_018038497.1 | F: GTGACCCAGCACAGTTCCTCTT | 74 |
| R: TTCCGGGCTCGGTGATTTAG | |||
| FLT1 | XM_018056600.1 | F: TGCCAGCAAGTGGGAGTTT | 102 |
| R: TTGATGCCGAATGCCGATG | |||
| KDR | XM_005681629.3 | F: TCAACAGGATGGCAAAGACT | 96 |
| R: GAAACAGGTGAGGTAGGCAGA | |||
| ANGPT1 | XM_ 005689163.3 | F: CTGGTGGTTTGATGCCTGTG | 182 |
| R: GCTCTTGATTGCTATTGTGCC | |||
| ANGPT2 | XM_013975359.2 | F: GACTGGTCAGAAACGCTACG | 168 |
| R: GGCTGGCTTATGCTGCTTATT | |||
| TEK | XM_018051890.1 | F: GGGGAAAGGTATGAGTGCTAA | 117 |
| R: AGGATGGCTTCTACAGGTGAG | |||
| FGF2 | XM_018061205.1 | F: GCAAACCGTTACCTTGCTAT | 153 |
| R: TCGTTTCAGTGCCACATACC | |||
| FGFR2 | XM_018041441.1 | F: ACTGTGGTTGGAGGCGATGT | 105 |
| R: CCCGTATTTACTGCCGTTCTT | |||
| β-actin | NM_001314342.1 | F: TGATATTGCTGCGCTCGTGGT | 189 |
| R: GTCAGGATGCCTCTCTTGCTC |
| Chromosomal Region | Chr27:39007363 to 39007851 |
|---|---|
| Target sequence (3′→5′) | ggagtaagtatttcaggttcctgggccacaC[1]GggctctgtcacagctgctC[2]GactctgcC[3]GttgtagtgtagaagccatcaagagaaaaccctC[4]GaggatgggaggtggctgtgtgccaaC[5]GaagcC[6]GtattC[7]GttgacactgaaatttgaacttatcataatttccaC [8]GtcatgaaatattctctggatttttttcagtcttttaaaaatgtaggaaccattcttagctcactggcC[9]GttcC[10]GC[11]GgggacC[12]GcaggccctggcctgC[13]GggctggagggggttgC[14]GggggggctgtgacaC[15]GgtctccaggcctctcC[16]GggC[17]GcaggcagtgtaagagggaacctccaggagccttccaC[18]GcctcagagcagC[19]GccC[20]GttctcccaaggaC[21]GagaactgtgagacaggcC[22]GggggaaagggctcattgaagggaC[23]GggcacattC[24]GaccaaaaaggaaagtgcC[25]Gcttggaagctggatctaaactcca |
| location | 5′UTR; N-shore |
| Primers (5′→3′) | Forward: aggaagagagGGAGTAAGTATTTTAGGTTTTTGGGT Reverse: cagtaatacgactcactatagggagaaggctTAAAATTTAAATCCAACTTCCAAAC |
| Amplicon length (bp) | 489 |
| Angiogenic Factor | Ki67 | CAD | CSD | CND | APC | |
|---|---|---|---|---|---|---|
| VEGFA | r2 | 0.260 | 0.138 | −0.167 | −0.131 | 0.117 |
| p | 0.350 | 0.624 | 0.552 | 0.641 | 0.678 | |
| FLT1 | r2 | −0.174 | −0.222 | 0.396 | 0.378 | −0.398 |
| p | 0.535 | 0.427 | 0.144 | 0.165 | 0.142 | |
| KDR | r2 | −0.274 | −0.474 | −0.074 | 0.258 | −0.505 |
| p | 0.322 | 0.074 | 0.794 | 0.352 | 0.055 | |
| ANGPT1 | r2 | 0.063 | −0.092 | 0.469 | 0.437 | −0.265 |
| p | 0.823 | 0.744 | 0.077 | 0.103 | 0.339 | |
| ANGPT2 | r2 | 0.004 | −0.404 | 0.308 | 0.641 | −0.589 |
| p | 0.988 | 0.135 | 0.265 | 0.010 | 0.021 | |
| TEK | r2 | −0.231 | −0.377 | −0.625 | −0.288 | −0.077 |
| p | 0.408 | 0.165 | 0.013 | 0.299 | 0.784 | |
| FGF2 | r2 | −0.707 | −0.716 | −0.648 | −0.189 | −0.399 |
| p | 0.003 | 0.003 | 0.009 | 0.501 | 0.140 | |
| FGFR2 | r2 | −0.741 | −0.505 | −0.484 | −0.238 | −0.193 |
| p | 0.002 | 0.055 | 0.067 | 0.393 | 0.492 | |
| CpG Sites | CAD | CSD | CND | APC | ANGPT2 | |
|---|---|---|---|---|---|---|
| CpG 2 | r2 | −0.361 | −0.654 | −0.464 | −0.071 | −0.325 |
| p | 0.205 | 0.011 | 0.095 | 0.809 | 0.256 | |
| CpG 4/18 | r2 | −0.505 | −0.155 | 0.189 | −0.508 | 0.231 |
| p | 0.066 | 0.598 | 0.517 | 0.064 | 0.427 | |
| CpG 6 | r2 | −0.565 | −0.342 | −0.023 | −0.421 | 0.191 |
| p | 0.035 | 0.232 | 0.937 | 0.134 | 0.513 | |
| CpG 7 | r2 | −0.545 | −0.481 | −0.095 | −0.413 | −0.187 |
| p | 0.044 | 0.082 | 0.746 | 0.142 | 0.522 | |
| CpG 8/12/24 | r2 | −0.036 | −0.068 | 0.011 | 0.040 | −0.087 |
| p | 0.902 | 0.816 | 0.970 | 0.892 | 0.768 | |
| CpG 9.10.11 | r2 | −0.517 | −0.495 | −0.233 | −0.310 | 0.038 |
| p | 0.059 | 0.072 | 0.424 | 0.280 | 0.897 | |
| CpG 13 | r2 | −0.523 | −0.393 | −0.036 | −0.443 | 0.108 |
| p | 0.055 | 0.165 | 0.902 | 0.113 | 0.713 | |
| CpG 14 | r2 | −0.532 | −0.388 | −0.088 | −0.440 | 0.108 |
| p | 0.050 | 0.171 | 0.765 | 0.115 | 0.713 | |
| CpG 15 | r2 | −0.572 | 0.184 | 0.678 | −0.873 | 0.594 |
| p | 0.033 | 0.529 | 0.008 | 0.000 | 0.025 | |
| CpG 16.17 | r2 | −0.250 | −0.676 | −0.417 | 0.008 | −0.520 |
| p | 0.388 | 0.008 | 0.138 | 0.978 | 0.057 | |
| CpG 22 | r2 | −0.417 | −0.255 | 0.014 | −0.427 | 0.161 |
| p | 0.138 | 0.379 | 0.963 | 0.128 | 0.582 | |
| CpG 23 | r2 | 0.248 | −0.051 | −0.133 | 0.422 | −0.534 |
| p | 0.393 | 0.863 | 0.650 | 0.133 | 0.049 | |
| CpG 25 | r2 | −0.059 | −0.484 | −0.439 | 0.107 | −0.485 |
| p | 0.842 | 0.079 | 0.117 | 0.717 | 0.079 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, P.; Luo, N.; Zhao, L.; Zhao, Y. Vascular Distribution and Expression Patterns of Angiogenic Factors in Caruncle during the Early Stage of Pregnancy in Goats (Capra hircus). Animals 2023, 13, 99. https://doi.org/10.3390/ani13010099
Wang P, Luo N, Zhao L, Zhao Y. Vascular Distribution and Expression Patterns of Angiogenic Factors in Caruncle during the Early Stage of Pregnancy in Goats (Capra hircus). Animals. 2023; 13(1):99. https://doi.org/10.3390/ani13010099
Chicago/Turabian StyleWang, Pan, Nanjian Luo, Le Zhao, and Yongju Zhao. 2023. "Vascular Distribution and Expression Patterns of Angiogenic Factors in Caruncle during the Early Stage of Pregnancy in Goats (Capra hircus)" Animals 13, no. 1: 99. https://doi.org/10.3390/ani13010099
APA StyleWang, P., Luo, N., Zhao, L., & Zhao, Y. (2023). Vascular Distribution and Expression Patterns of Angiogenic Factors in Caruncle during the Early Stage of Pregnancy in Goats (Capra hircus). Animals, 13(1), 99. https://doi.org/10.3390/ani13010099