
CircMGA Depresses Myoblast Proliferation and Promotes Myotube Formation through miR-144-5p/FAP Signal

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Primers, Vector Construction and RNA Oligonucleotides
2.2. Cell Culture and Transfection
2.3. RNA Exaction, cDNA Synthesis and Quantitative Real-Time PCR (qRT-PCR)
2.4. Flow Cytometry Analysis
2.5. EdU (5-Ethynyl-2′-Deoxyuridine) Assay
2.6. Immunofluorescence
2.7. Dual-Luciferase Reporter Assay
2.8. Biotin-Labeled miRNA Pulldown Assay
2.9. Statistical Analysis
3. Results
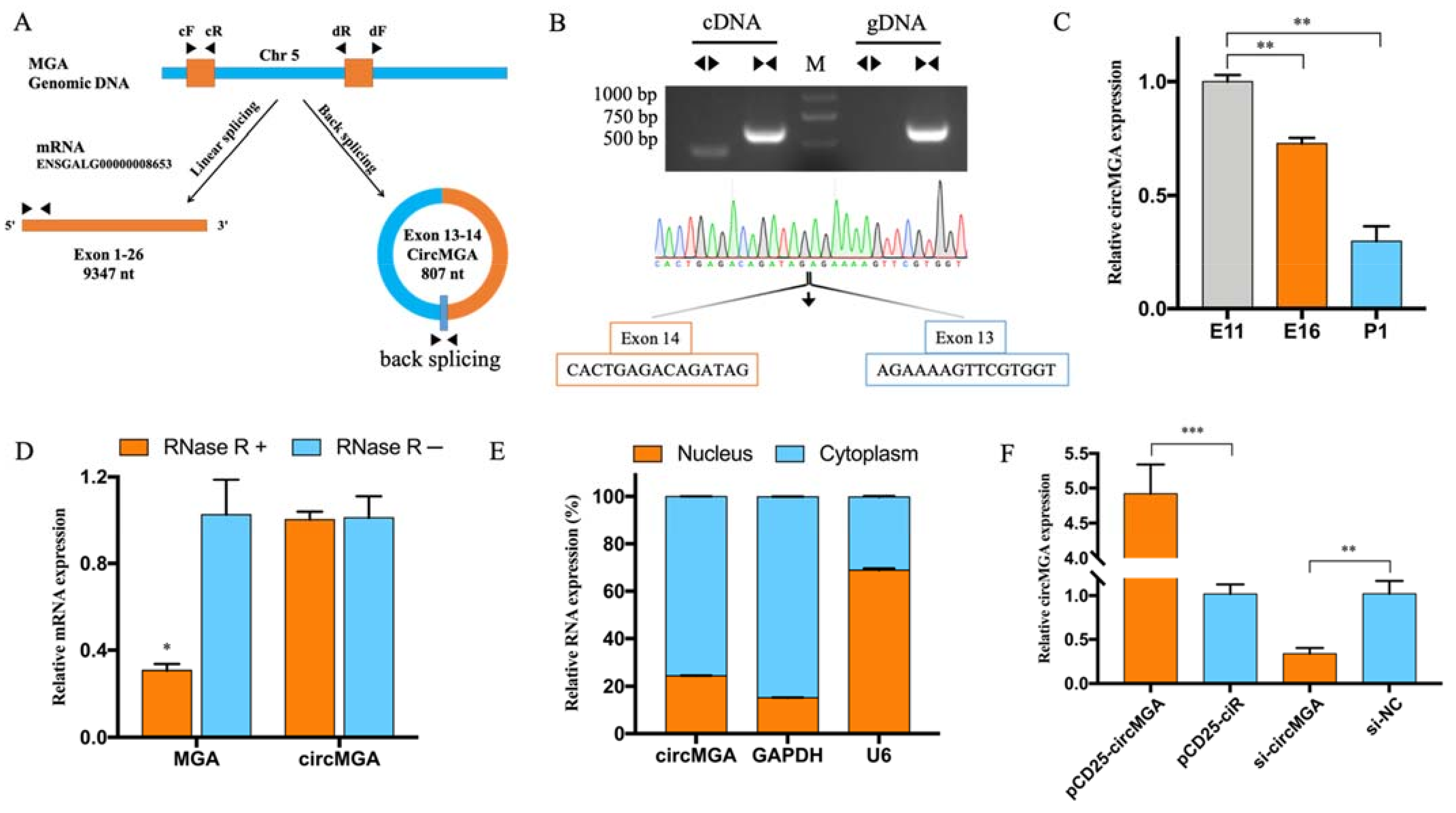
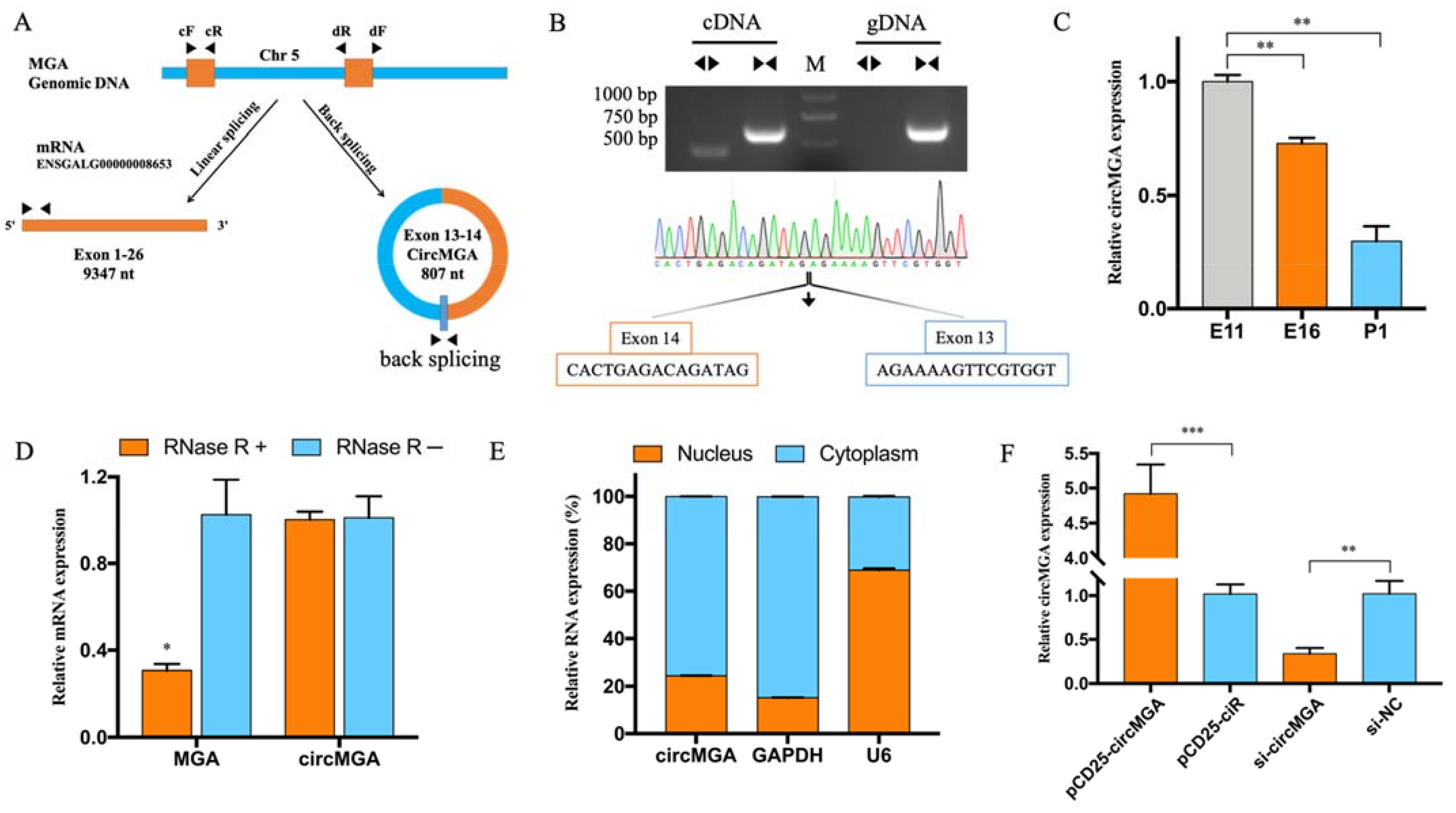
3.1. CircMGA Differentially Expressed during Embryonic Development
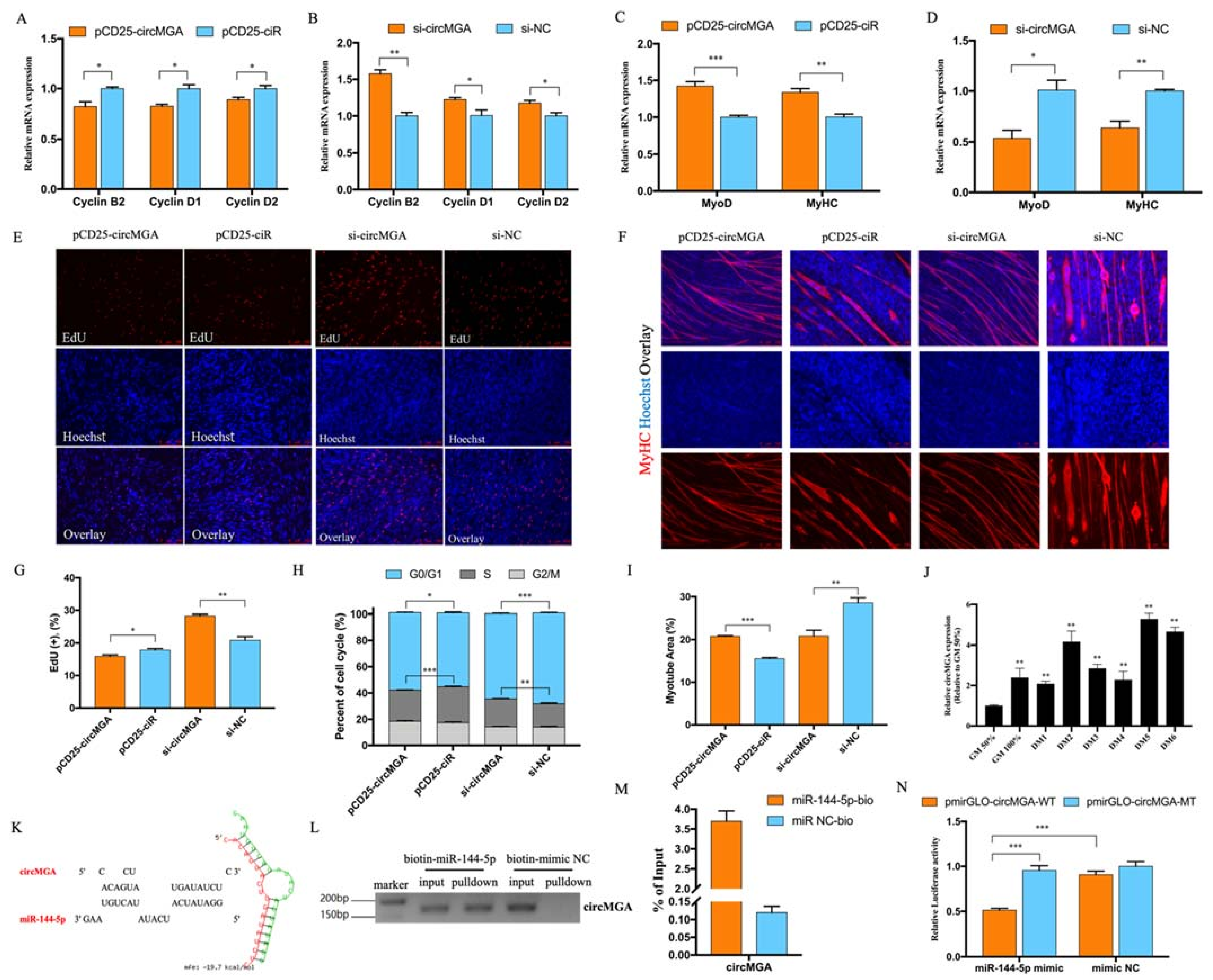
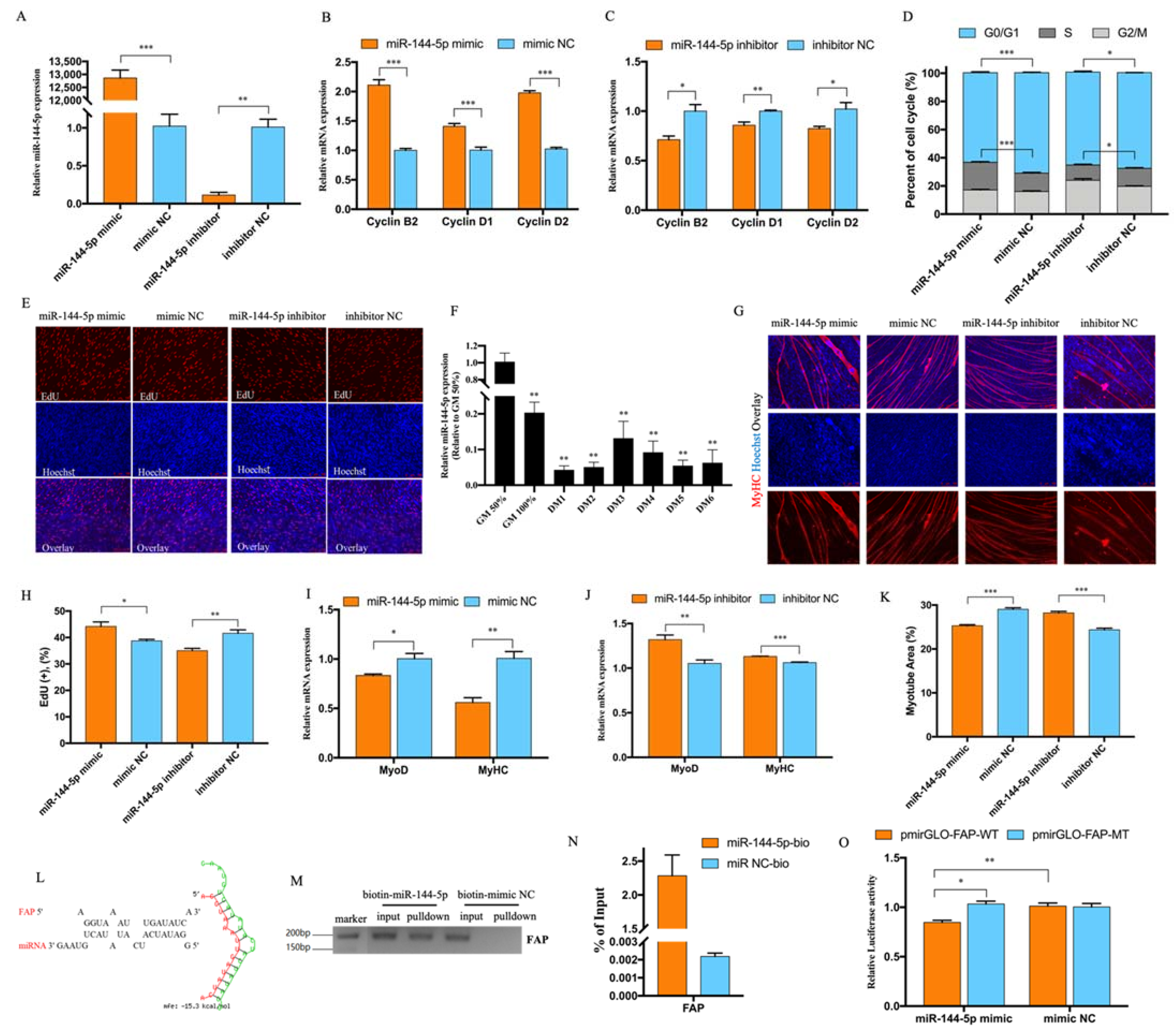
3.2. CircMGA Depressed Myoblast Cell Cycle Progress, Promoted Myotube Formation, and Sponged miR-144-5p
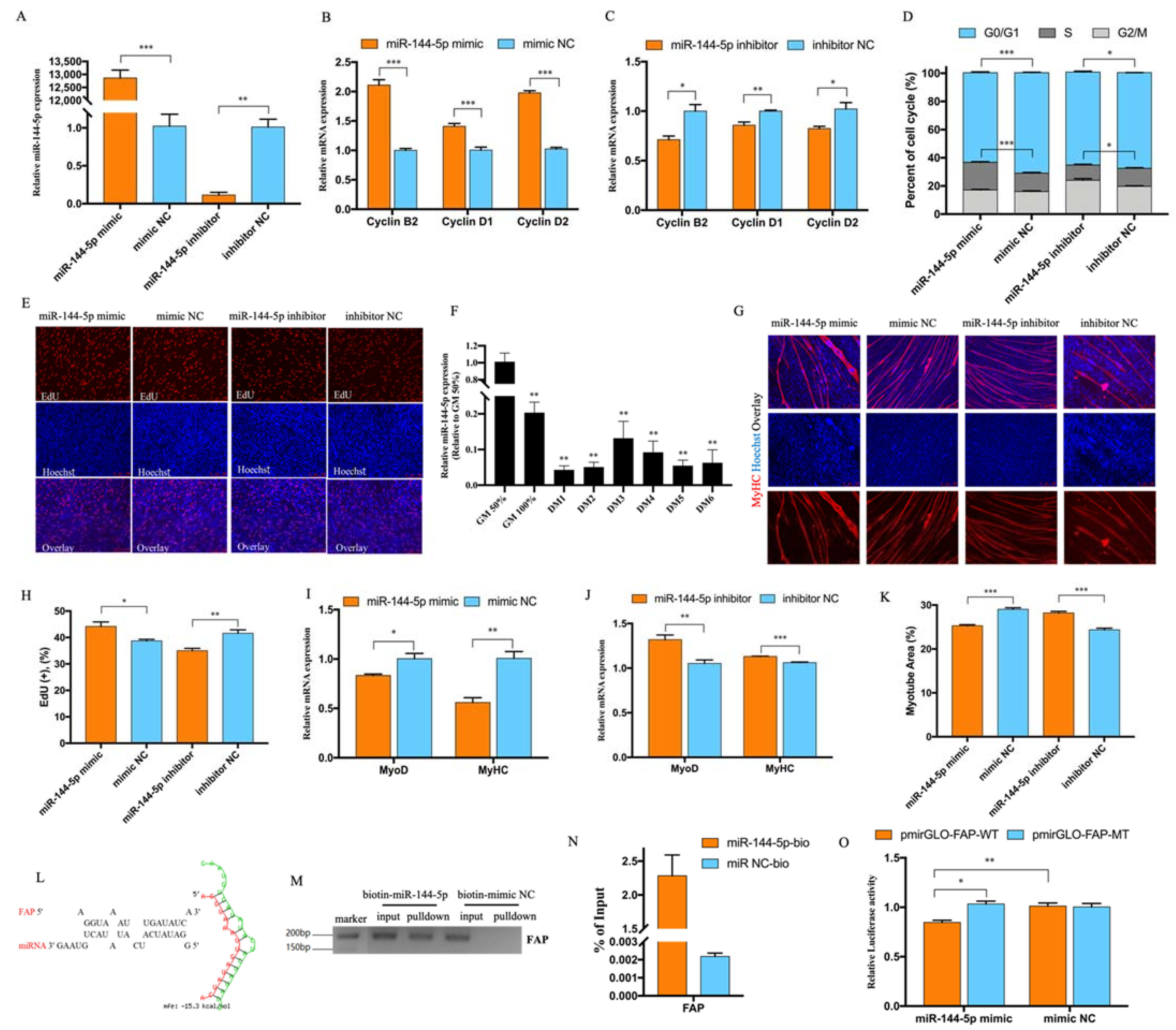
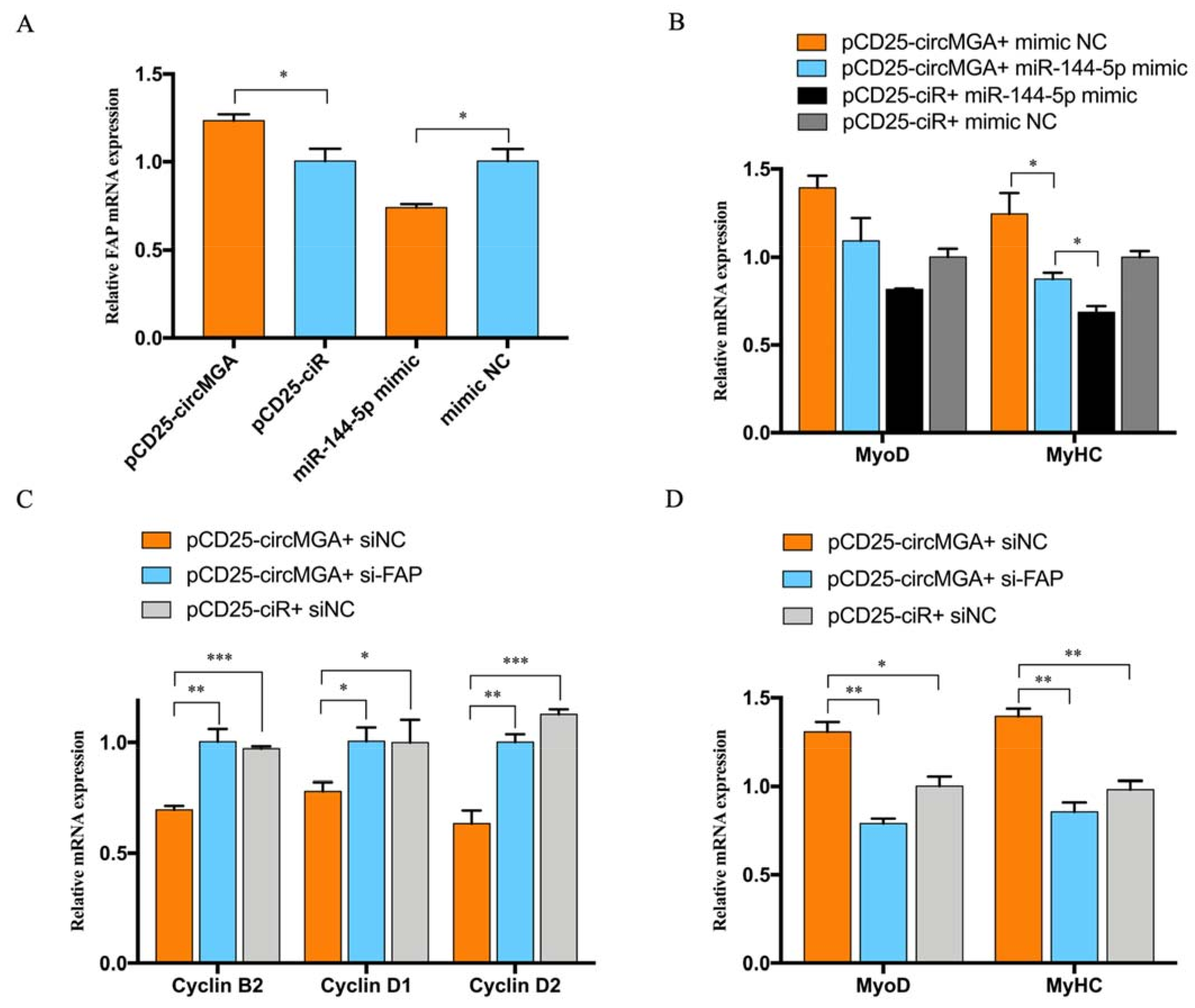
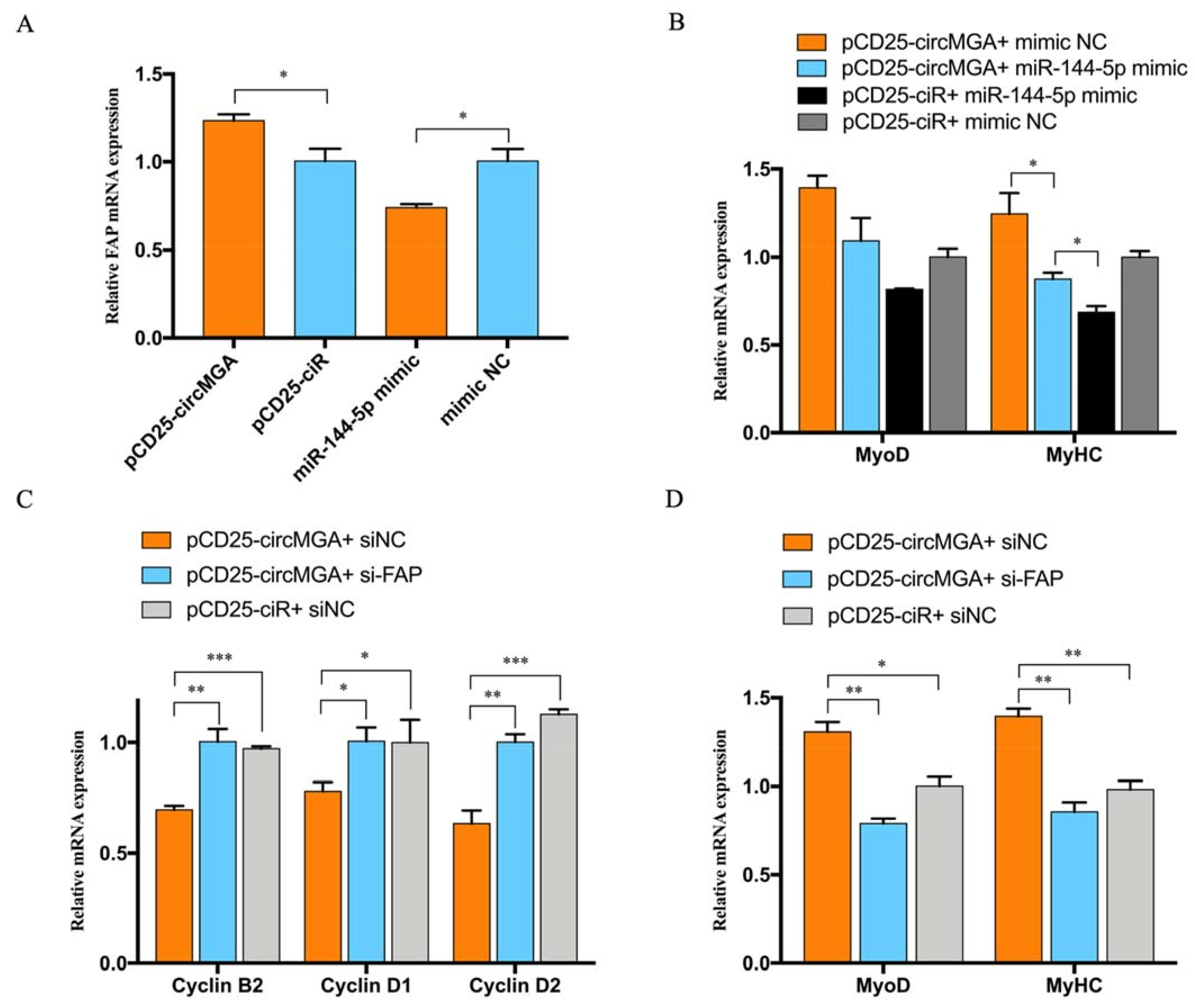
3.3. miR-144-5p Accelerated Myoblast Cell Cycle and Inhibited Myotube Formation by Targeting FAP
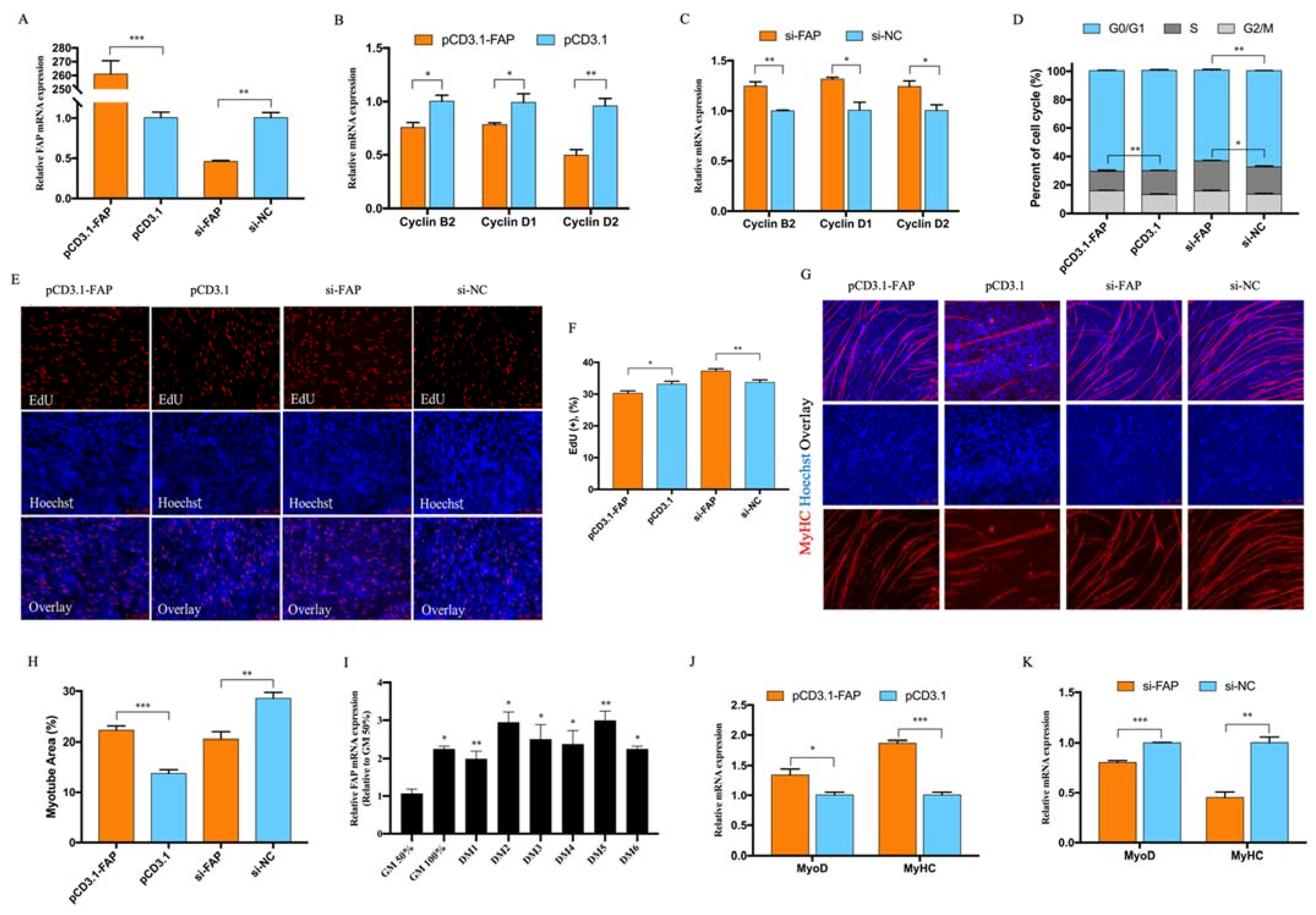
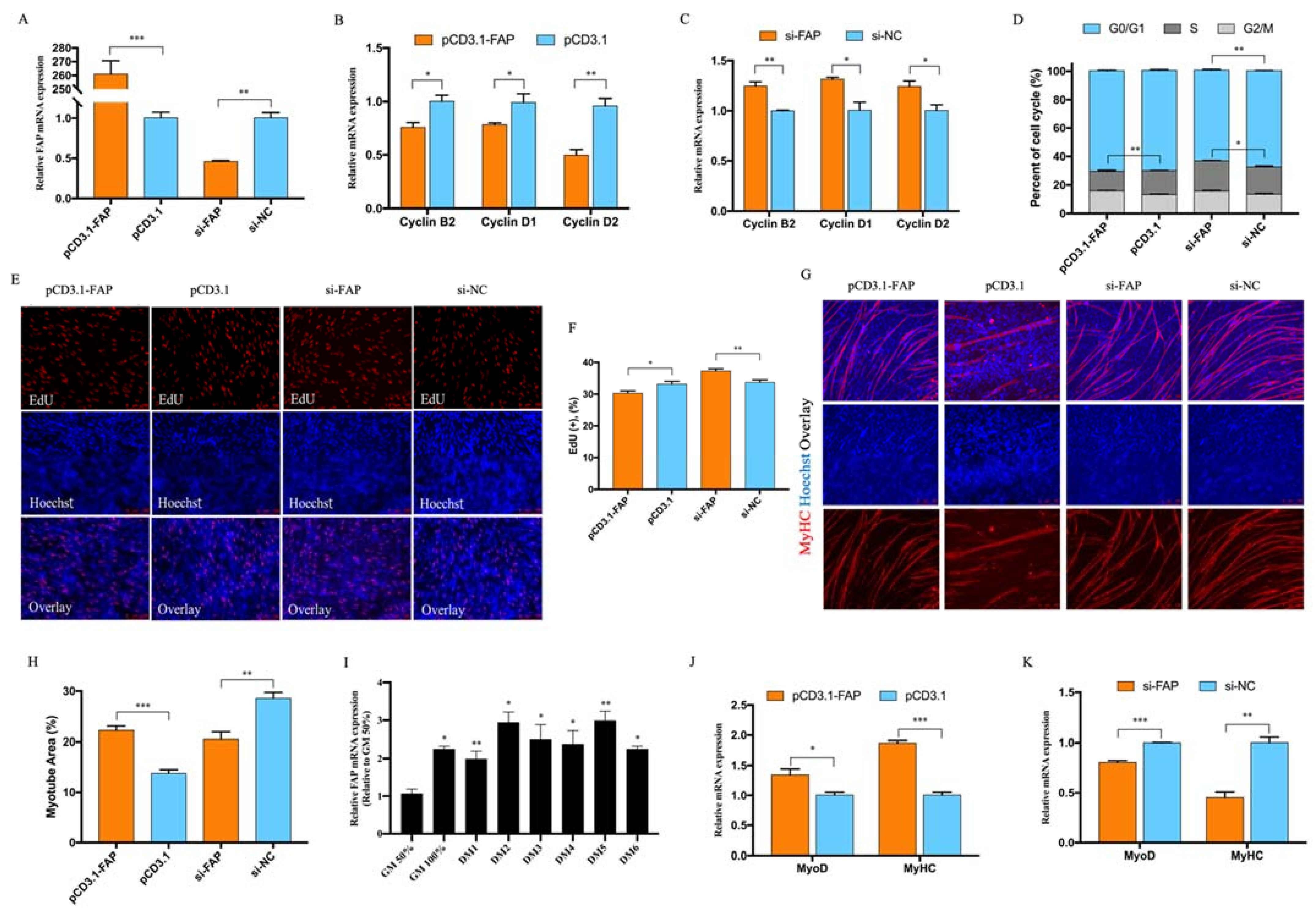
3.4. FAP Inhibited Myoblast Proliferation and Promote Myoblast Differentiation
3.5. CircMGA Promote Myotube Formation through miR-144-5p/FAP Signal
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regu-latory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef] [PubMed]
- Salzman, J.; Gawad, C.; Wang, P.L.; Lacayo, N.; Brown, P.O. Circular RNAs Are the Predominant Transcript Isoform from Hundreds of Human Genes in Diverse Cell Types. PLoS ONE 2012, 7, e30733. [Google Scholar] [CrossRef] [Green Version]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [Green Version]
- Westholm, J.O.; Miura, P.; Olson, S.; Shenker, S.; Joseph, B.; Sanfilippo, P.; Celniker, S.E.; Graveley, B.R.; Lai, E.C. Genome-wide Analysis of Drosophila Circular RNAs Reveals Their Structural and Sequence Properties and Age-Dependent Neural Accumulation. Cell Rep. 2014, 9, 1966–1980. [Google Scholar] [CrossRef] [Green Version]
- Huang, A.; Zheng, H.; Wu, Z.; Chen, M.; Huang, Y. Circular RNA-protein interactions: Functions, mechanisms, and identification. Theranostics 2020, 10, 3503–3517. [Google Scholar] [CrossRef]
- Du, W.W.; Yang, W.; Liu, E.; Yang, Z.; Dhaliwal, P.; Yang, B.B. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res. 2016, 44, 2846–2858. [Google Scholar] [CrossRef] [Green Version]
- Du, W.W.; Zhang, C.; Yang, W.; Yong, T.; Awan, F.M.; Yang, B.B. Identifying and Characterizing circRNA-Protein Interaction. Theranostics 2017, 7, 4183–4191. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, X.; Zhang, M.; Yan, S.; Sun, C.; Xiao, F.; Huang, N.; Yang, X.; Zhao, K.; Zhou, H.; et al. Novel Role of FBXW7 Circular RNA in Repressing Glioma Tumorigenesis. J. Natl. Cancer Inst. 2018, 110, 304–315. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Fan, X.; Mao, M.; Song, X.; Wu, P.; Zhang, Y.; Jin, Y.; Yang, Y.; Chen, L.-L.; Wang, Y.; et al. Extensive translation of circular RNAs driven by N6-methyladenosine. Cell Res. 2017, 27, 626–641. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Nie, Q.; Zhang, X. MicroRNAs Involved in Skeletal Muscle Differentiation. J. Genet. Genom. 2013, 40, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Sambasivan, R.; Tajbakhsh, S. Skeletal muscle stem cell birth and properties. Semin. Cell Dev. Biol. 2007, 18, 870–882. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, M.; Lian, D.; Li, Y.; Wang, J.; Deng, S.; Yu, K.; Lian, Z. Non-Coding RNA Regulates the Myogenesis of Skeletal Muscle Satellite Cells, Injury Repair and Diseases. Cells 2019, 8, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Chao, Z.; Zhang, R.; Ding, R.; Wang, Y.; Wu, W.; Han, Q.; Li, C.; Xu, H.; Wang, L.; et al. Circular RNA Regulation of Myogenesis. Cells 2019, 8, 885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.-L.; Song, C.-H.; Wang, P.; Dai, L.-P.; Zhang, J.-Y.; Wang, K.-J. Competing endogenous RNA networks and gastric cancer. World J. Gastroenterol. 2015, 21, 11680–11687. [Google Scholar] [CrossRef]
- Fan, C.; Tang, Y.; Wang, J.; Xiong, F.; Guo, C.; Wang, Y.; Zhang, S.; Gong, Z.; Wei, X.; Zhaoyang, Z.; et al. Role of long non-coding RNAs in glucose metabolism in cancer. Mol. Cancer 2017, 16, 130. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Cheng, Z.; Pang, Y.; Cui, L.; Qian, T.; Quan, L.; Zhao, H.; Shi, J.; Ke, X.; Fu, L. Role of microRNAs, circRNAs and long noncoding RNAs in acute myeloid leukemia. J. Hematol. Oncol. 2019, 12, 51. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Du, Y.; Yang, X.; Mo, Y.; Fan, C.; Xiong, F.; Ren, D.; Ye, X.; Li, C.; Wang, Y.; et al. Circular RNAs function as ceRNAs to regulate and control human cancer progression. Mol. Cancer 2018, 17, 79. [Google Scholar] [CrossRef]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef]
- Baek, D.; Villén, J.; Shin, C.; Camargo, F.D.; Gygi, S.P.; Bartel, D.P. The impact of microRNAs on protein output. Nature 2008, 455, 64–71. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-F.; Mandel, E.M.; Thomson, J.M.; Wu, Q.; E Callis, T.; Hammond, S.M.; Conlon, F.L.; Wang, D.-Z. The Role of MicroRNA-1 and MicroRNA-133 in Skeletal Muscle Proliferation and Differentiation. Nat. Genet. 2006, 38, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ouyang, H.; Wang, Z.; Chen, B.; Nie, Q. A Novel Circular RNA Generated by FGFR2 Gene Promotes Myoblast Proliferation and Differentiation by Sponging miR-133a-5p and miR-29b-1-5p. Cells 2018, 7, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.-D.; Shen, C.-H.; Tao, Y.-F.; Zhang, X.-F.; Zhang, Q.-B.; Ma, Z.-Y.; Wang, Z.-X. MicroRNA-144 suppresses the expression of cytokines through targeting RANKL in the matured immune cells. Cytokine 2018, 108, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.; Wang, H.; Wang, H.; Wu, X.; Yang, L.; Wang, C.; Li, X.; Jin, Y.; Li, M.; Wang, L.; et al. Circular RNA Circ_0006282 Promotes Cell Proliferation and Metastasis in Gastric Cancer by Regulating MicroRNA-144-5p/Tyrosine 3-Monooxygenase/Tryptophan 5-Monooxygenase Activation Protein β Axis. Cancer Manag. Res. 2021, 13, 815–827. [Google Scholar] [CrossRef]
- Ouyang, H.; Chen, X.; Wang, Z.; Yu, J.; Jia, X.; Li, Z.; Luo, W.; Abdalla, B.A.; Jebessa, E.; Nie, Q.; et al. Circular RNAs are abundant and dynamically expressed during embryonic muscle development in chickens. DNA Res. 2018, 25, 71–86. [Google Scholar] [CrossRef]
- Hamson, E.J.; Keane, F.M.; Tholen, S.; Schilling, O.; Gorrell, M.D. Understanding fibroblast activation protein (FAP): Substrates, activities, expression and targeting for cancer therapy. Proteom. Clin. Appl. 2014, 8, 454–463. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Piwecka, M.; Glažar, P.; Hernandez-Miranda, L.R.; Memczak, S.; Wolf, S.A.; Rybak-Wolf, A.; Filipchyk, A.; Klironomos, F.; Cerda Jara, C.A.; Fenske, P.; et al. Loss of a mammalian circular RNA locus causes miRNA deregulation and affects brain function. Science 2017, 357, 6357. [Google Scholar] [CrossRef] [Green Version]
- Cooper, D.A.; Cortés-López, M.; Miura, P. Genome-Wide circRNA Profiling from RNA-seq Data. Methods Mol. Biol. 2018, 1724, 27–41. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Chen, C.-Y.; Mai, T.-L.; Chuang, C.-F.; Chen, Y.-C.; Gupta, S.K.; Yen, L.; Wang, Y.-D.; Chuang, T.-J. Genome-wide, integrative analysis of circular RNA dysregulation and the corresponding circular RNA-microRNA-mRNA regulatory axes in autism. Genome Res. 2020, 30, 375–391. [Google Scholar] [CrossRef] [Green Version]
- Llabata, P.; Mitsuishi, Y.; Choi, P.; Cai, D.; Francis, J.M.; Torres-Diz, M.; Udeshi, N.D.; Golomb, L.; Wu, Z.; Zhou, J.; et al. Multi-Omics Analysis Identifies MGA as a Negative Regulator of the MYC Pathway in Lung Adenocarcinoma. Mol. Cancer Res. 2020, 18, 574–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferri, P.; Barbieri, E.; Burattini, S.; Guescini, M.; D’Emilio, A.; Biagiotti, L.; Del Grande, P.; De Luca, A.; Stocchi, V.; Falcieri, E. Expression and subcellular localization of myogenic regulatory factors during the differentiation of skeletal muscle C2C12 myoblasts. J. Cell. Biochem. 2009, 108, 1302–1317. [Google Scholar] [CrossRef] [PubMed]
- Sartorelli, V.; Caretti, G. Mechanisms underlying the transcriptional regulation of skeletal myogenesis. Curr. Opin. Genet. Dev. 2005, 15, 528–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkes, C.A.; Tapscott, S.J. MyoD and the transcriptional control of myogenesis. Semin. Cell Dev. Biol. 2005, 16, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Tajsharghi, H.; Oldfors, A. Myosinopathies: Pathology and mechanisms. Acta Neuropathol. 2012, 125, 3–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Nucleotide Sequences (5′-3′) | Application |
|---|---|---|
| d-circMGA | F: TCAGTACTGTGATCTCCAAGG | Divergent primer for circMGA |
| R: ATGGCACTGAAATTTTCACTC | ||
| c-MGA | F: CCTCATTATTGAATTGGCTTC | Convergent primer for MGA |
| R: GTCTCAGTGGTGGGATAAGAG | ||
| q-circMGA | F: TCCAAGGTAGCATCCAGTGC | qRT-PCR for circMGA |
| R: TTCATTCCAGCTGCAGTCAGAG | ||
| c-circMGA | F1: AGAAAAGTTCGTGGTCTTCCC | cloning full length primer of circMGA |
| R1: CTATCTGTCTCAGTGGTGGGAT | ||
| c-circMGA | F2: GGAATTCTAATACTTTCAGAGAAAAGTTCGTGGTCTTCCC | cloning full length primer of circMGA |
| R2: CGGGATCCAGTTGTTCTTACCTATCTGTCTCAGTGGTGGGAT | ||
| c-FAP | F: CGGGATCCATGAAGACCCGGCTAAAAGTA | cloning full CDS length primer of FAP |
| R: GCTCTAGACTATTCTGACAAAGAAAAACATT | ||
| q-FAP | F: GCTTCCGTCCAAAGTTGTCA | qRT-PCR for FAP |
| R: TGTGCTGTTGGTCATGATGGTA | ||
| Cyclin B2 | F: CAGTAAAGGCTACGAAAG | qRT-PCR for Cyclin B2 |
| R: ACATCCATAGGGACAGG | ||
| Cyclin D1 | F: CAGAAGTGCGAAGAGGAAGT | qRT-PCR for Cyclin D1 |
| R: CTGATGGAGTTGTCGGTGTA | ||
| Cyclin D2 | F: AACTTGCTCTACGACGACC | qRT-PCR for Cyclin D2 |
| R: TTCACAGACCTCCAACATC | ||
| MyoD | F: GCTACTACACGGAATCACCAAAT | qRT-PCR for MyoD |
| R: CTGGGCTCCACTGTCACTCA | ||
| MyHC | F: CTCCTCACGCTTTGGTAA | qRT-PCR for MyHC |
| R: TGATAGTCGTATGGGTTGGT | ||
| GAPDH | F: TCCTCCACCTTTGATGCG | qRT-PCR for GAPDH |
| R: GTGCCTGGCTCACTCCTT | ||
| β-actin | F: TTGTTGACAATGGCTCCGGT | qRT-PCR for β-actin |
| R: AACCATCACACCCTGATGTCT | ||
| U6 | F: CAAGGACCCATCGTTCCACA | qRT-PCR for U6 |
| R: CCATTGGACACGCAGAATGC |
| Name | Nucleotide Sequences (5′-3′) |
|---|---|
| si-circMGA | AGAUAGAAAAGUUCGUGTT |
| si-FAP | GCTCTTACACTGGATGATT |
| miR-144-5p mimic | GGAUAUCAUCAUAUACUGUAAG |
| miR-144-5p inhibitor | GAAUGUCAUAUACUACUAUAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Zhang, M.; Li, K.; Chen, Y.; Cai, D.; Chen, B.; Nie, Q. CircMGA Depresses Myoblast Proliferation and Promotes Myotube Formation through miR-144-5p/FAP Signal. Animals 2022, 12, 873. https://doi.org/10.3390/ani12070873
Wang Z, Zhang M, Li K, Chen Y, Cai D, Chen B, Nie Q. CircMGA Depresses Myoblast Proliferation and Promotes Myotube Formation through miR-144-5p/FAP Signal. Animals. 2022; 12(7):873. https://doi.org/10.3390/ani12070873
Chicago/Turabian StyleWang, Zhijun, Min Zhang, Kan Li, Yangfeng Chen, Danfeng Cai, Biao Chen, and Qinghua Nie. 2022. "CircMGA Depresses Myoblast Proliferation and Promotes Myotube Formation through miR-144-5p/FAP Signal" Animals 12, no. 7: 873. https://doi.org/10.3390/ani12070873
APA StyleWang, Z., Zhang, M., Li, K., Chen, Y., Cai, D., Chen, B., & Nie, Q. (2022). CircMGA Depresses Myoblast Proliferation and Promotes Myotube Formation through miR-144-5p/FAP Signal. Animals, 12(7), 873. https://doi.org/10.3390/ani12070873