
Successful Laparoscopic Oviductal Artificial Insemination in the Endangered Tsushima Leopard Cat (Prionailurus bengalensis euptilurus)

 , , , and
, , , and
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Animals and Husbandry
2.2. Behavioral Observations
2.3. Fecal Hormone Analysis
2.4. Induction of Ovarian Activity
2.5. Semen Collection and Processing
2.6. Vaginal Smear and Ovarian Assessment
2.7. Laparoscopic AI
2.8. Pregnancy Diagnosis and Observation of Delivery
2.9. Genetic Testing
3. Results
3.1. Behavioral Observation
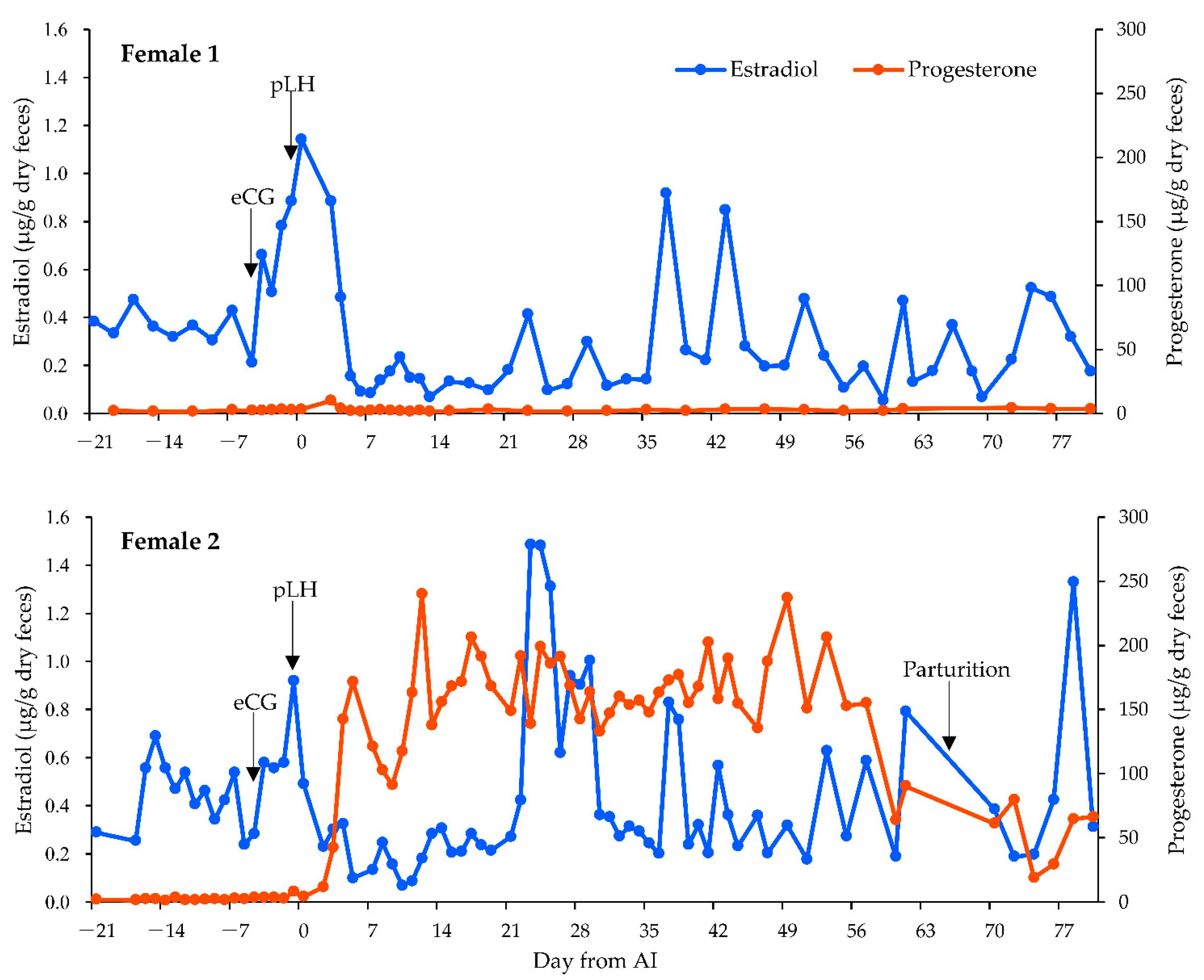
3.2. Fecal Hormone Analysis
3.3. Semen Characteristics
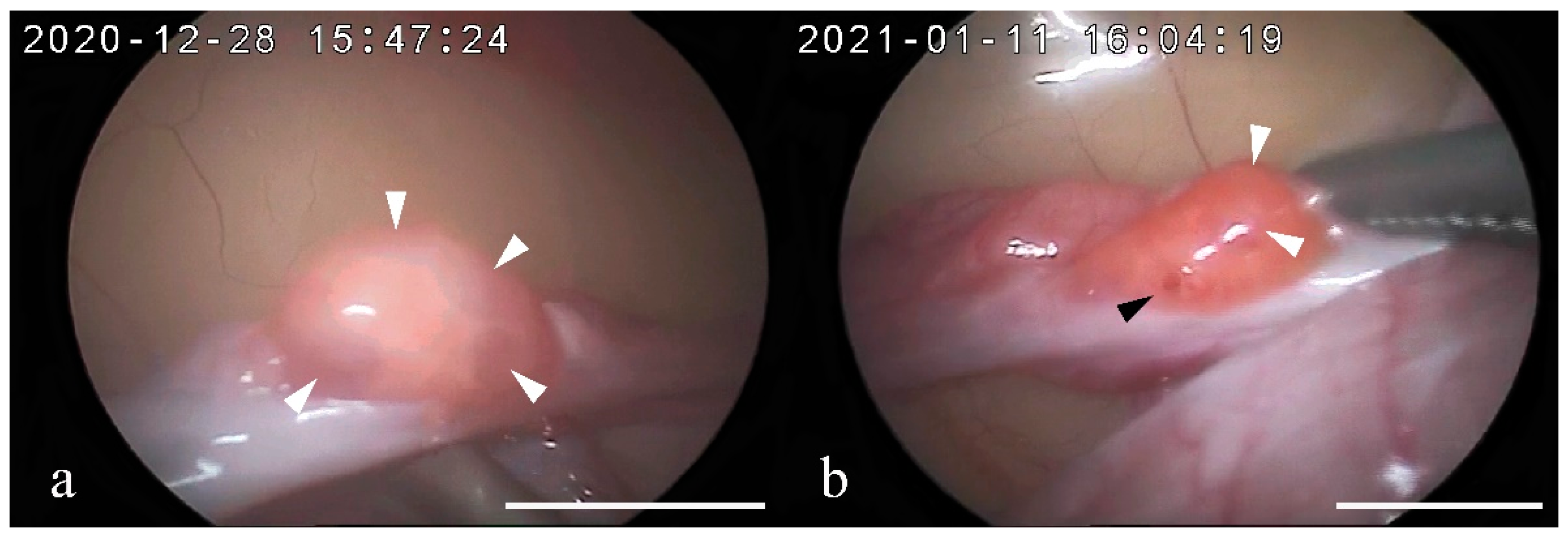
3.4. Vaginal Smear and Ovarian Assessment
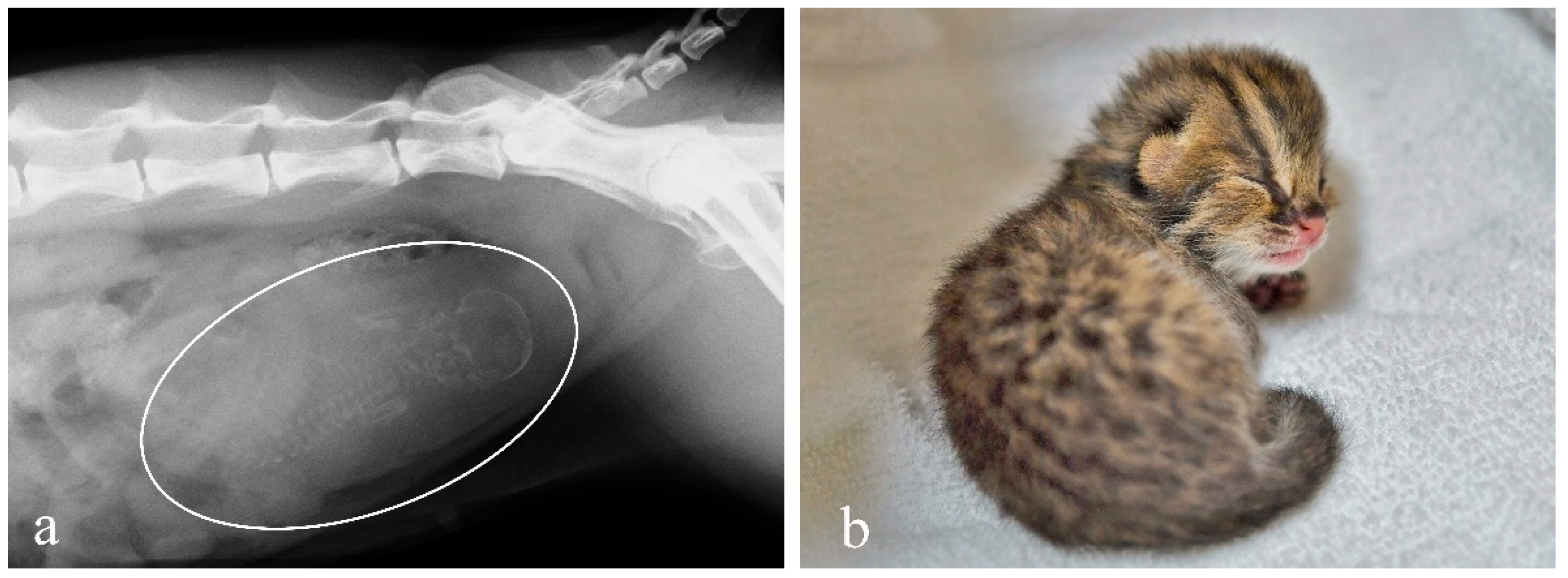
3.5. Pregnancy Diagnosis and Observation of Delivery
3.6. Genetic Testing
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Masuda, R.; Yoshida, M.C. Two Japanese Wildcats, the Tsushima Cat and the Iriomote Cat, Show the Same Mitochondrial DNA Lineage as the Leopard Cat Felis bengalensis. Zool. Sci. 1995, 12, 655–659. [Google Scholar] [CrossRef] [PubMed]
- Izawa, M.; Doi, T.; Nakanishi, N.; Teranishi, A. Ecology and Conservation of Two Endangered Subspecies of the Leopard Cat (Prionailurus bengalensis) on Japanese Islands. Biol. Conserv. 2009, 142, 1884–1890. [Google Scholar] [CrossRef]
- Machii, J.; Monji, K.; Imada, R.; Tanaka, S.; Nagao, E.; Saito, Y.; Hamachi, M. Husbandry and Breeding of the Tsushima Leopard Cat, Felis bengalensis euptilura. J Jpn. Assoc. Zool. Aquat. 2002, 43, 94–103. (In Japanese) [Google Scholar]
- Chatdarong, K.; Axnér, E.; Manee-In, S.; Thuwanut, P.; Linde-Forsberg, C. Pregnancy in the Domestic Cat After Vaginal or Transcervical Insemination with Fresh and Frozen Semen. Theriogenology 2007, 68, 1326–1333. [Google Scholar] [CrossRef]
- Dresser, B.L.; Kramer, L.; Reece, B.; Russell, P.T. Induction of ovulation and successful artificial insemination in a Persian leopard (Panthera pardus saxicolor). Zoo Biol. 1982, 1, 55–57. [Google Scholar] [CrossRef]
- Howard, J.; Donoghue, A.M.; Barone, M.A.; Goodrowe, K.L.; Blumer, E.S.; Snodgrass, K.; Starnes, D.; Tucker, M.; Bush, M.; Wildt, D.E. Successful induction of ovarian activity and laparoscopic intrauterine artificial insemination in the cheetah (Acinonyx jubatus). J. Zoo Wildl. Med. 1992, 23, 288–300. [Google Scholar]
- Donoghue, A.M.; Johnston, L.A.; Armstrong, D.L.; Simmons, L.G.; Wildt, D.E. Birth of a Siberian tiger cub (Panthera tigris altaica) following laparoscopic intrauterine artificial insemination. J. Zoo Wildl. Med. 1993, 23, 185–189. [Google Scholar]
- Barone, M.A.; Wildt, D.E.; Byers, A.P.; Roelke, M.E.; Glass, C.M.; Howard, J.G. Gonadotrophin dose and timing of anaesthesia for laparoscopic artificial insemination in the puma (Felis concolor). Reproduction 1994, 101, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Swanson, W.F.; Howard, J.; Roth, T.L.; Brown, J.L.; Alvarado, T.; Burton, M.; Starnes, D.; Wildt, D.E. Responsiveness of ovaries to exogenous gonadotrophins and laparoscopic artificial insemination with frozen–thawed spermatozoa in ocelots (Felis pardalis). Reproduction 1996, 106, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, A.M.; Byers, A.P.; Johnston, L.A.; Armstrong, D.L.; Wildt, D.E. Timing of ovulation after gonadotrophin induction and its importance to successful intrauterine insemination in the tiger (Panthera tigris). Reproduction 1996, 107, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Howard, J.; Byers, A.P.; Brown, J.L.; Barrett, S.J.; Evans, M.Z.; Schwartz, R.J.; Wildt, D.E. Successful ovulation induction and laparoscopic intrauterine artificial insemination in the clouded leopard (Neofelis nebulosa). Zoo Biol. 1996, 15, 55–69. [Google Scholar] [CrossRef]
- Roth, T.L.; Armstrong, D.L.; Barrie, M.T.; Wildt, D.E. Seasonal effects on ovarian responsiveness to exogenous gonadotrophins and successful artificial insemination in the snow leopard (Uncia uncia). Reprod. Fertil. Dev. 1997, 9, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Howard, J.; Roth, T.L.; Byers, A.P.; Swanson, W.F.; Wildt, D.E. Sensitivity to exogenous gonadotropins for ovulation induction and laparoscopic artificial insemination in the cheetah and clouded leopard. Biol. Reprod. 1997, 56, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Howard, J.G.; Wildt, D.E. Approaches and Efficacy of Artificial Insemination in Felids and Mustelids. Theriogenology 2009, 71, 130–148. [Google Scholar] [CrossRef]
- Lueders, I.; Ludwig, C.; Schroeder, M.; Mueller, K.; Zahmel, J.; Dehnhard, M. Successful Nonsurgical Artificial Insemination and Hormonal Monitoring in an Asiatic Golden Cat (Catopuma temmincki). J. Zoo Wildl. Med. 2014, 45, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Callealta, I.; Ganswindt, A.; Malan, M.; Lueders, I. Non-Surgical Artificial Insemination Using a GnRH Analogue for Ovulation Induction During Natural Oestrus in African Lions (Panthera leo). Theriogenology 2019, 139, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Conforti, V.A.; Bateman, H.L.; Schook, M.W.; Newsom, J.; Lyons, L.A.; Grahn, R.A.; Deddens, J.A.; Swanson, W.F. Laparoscopic oviductal artificial insemination improves pregnancy success in exogenous gonadotropin-treated domestic cats as a model for endangered felids. Biol. Reprod. 2013, 89, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lambo, C.A.; Bateman, H.L.; Swanson, W.F. 3 Application of Laparoscopic Oviductal Artificial Insemination for Conservation Management of Brazilian Ocelots and Amur Tigers. Reprod. Fertil. Dev. 2014, 26, 116. [Google Scholar] [CrossRef]
- Swanson, W.F.; Bateman, H.L.; Herrick, J.R.; Lyons, L.A.; Vansandt, L.M. 121 Fixed-Time Laparoscopic Oviductal Artificial Insemination (LO-AI) with Frozen–Thawed Semen in Pallas’ Cats (Otocolobus manul). Reprod. Fertil. Dev. 2016, 28, 190–191. [Google Scholar] [CrossRef]
- Swanson, W.F. Practical Application of Laparoscopic Oviductal Artificial Insemination for the Propagation of Domestic Cats and Wild Felids. Reprod. Fertil. Dev. 2018, 31, 27–39. [Google Scholar] [CrossRef]
- Adachi, I.; Kusuda, S.; Nagao, E.; Taira, Y.; Asano, M.; Tsubota, T.; Doi, O. Fecal steroid metabolites and reproductive monitoring in a female Tsushima leopard cat (Prionailurus bengalensis euptilurus). Theriogenology 2010, 74, 1499–1503. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.H.; Swanson, W.F.; Brown, J.L. Chorionic gonadotropin administration in domestic cats causes an abnormal endocrine environment that disrupts oviductal embryo transport. Theriogenology 2000, 54, 1117–1131. [Google Scholar] [CrossRef]
- Howard, J.G.; Brown, J.L.; Bush, M.; Wildt, D.E. Teratospermic and Normospermic Domestic Cats: Ejaculate Traits, Pituitary-Gonadal Hormones, and Improvement of Spermatozoal Motility and Morphology After Swim-Up Processing. J. Androl. 1990, 11, 204–215. [Google Scholar] [PubMed]
- Zambelli, D.; Prati, F.; Cunto, M.; Iacono, E.; Merlo, B. Quality and In Vitro Fertilizing Ability of Cryopreserved Cat Spermatozoa Obtained by Urethral Catheterization After Medetomidine Administration. Theriogenology 2008, 69, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Björndahl, L.; Söderlund, I.; Kvist, U. Evaluation of the One-Step Eosin-Nigrosin Staining Technique for Human Sperm Vitality Assessment. Hum. Reprod. 2003, 18, 813–816. [Google Scholar] [CrossRef]
- Herrick, J.R.; Bond, J.B.; Magarey, G.M.; Bateman, H.L.; Krisher, R.L.; Dunford, S.A.; Swanson, W.F. Toward a Feline-Optimized Culture Medium: Impact of Ions, Carbohydrates, Essential Amino Acids, Vitamins, and Serum on Development and Metabolism of In Vitro Fertilization-Derived Feline Embryos Relative to Embryos Grown In Vivo. Biol. Reprod. 2007, 76, 858–870. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ito, H.; Inoue-Murayama, M. The Tsushima Leopard Cat Exhibits Extremely Low Genetic Diversity Compared with the Korean Amur Leopard Cat: Implications for Conservation. PeerJ 2019, 7, e7297. [Google Scholar] [CrossRef]
- Babraham Institute. Trim Galore, Version 0.6.5; Babraham Institute: Cambridge, UK, 2019; Available online: https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 18 September 2021).
- Buckley, R.M.; Davis, B.W.; Brashear, W.A.; Farias, F.H.G.; Kuroki, K.; Graves, T.; Hillier, L.W.; Kremitzki, M.; Li, G.; Middleton, R.P.; et al. A New Domestic Cat Genome Assembly Based on Long Sequence Reads Empowers Feline Genomic Medicine and Identifies a Novel Gene for Dwarfism. PLoS Genet. 2020, 16, e1008926. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-Generation PLINK: Rising to the Challenge of Larger and Richer Datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef] [PubMed]
- Manichaikul, A.; Mychaleckyj, J.C.; Rich, S.S.; Daly, K.; Sale, M.; Chen, W.M. Robust Relationship Inference in Genome-Wide Association Studies. Bioinformatics 2010, 26, 2867–2873. [Google Scholar] [CrossRef] [PubMed]
- Swanson, W.F. Application of Assisted Reproduction for Population Management in Felids: The Potential and Reality for Conservation of Small Cats. Theriogenology 2006, 66, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Lermen, D.; Blömeke, B.; Browne, R.; Clarke, A.N.N.; Dyce, P.W.; Fixemer, T.; Fuhr, G.R.; Holt, W.V.; Jewgenow, K.; Lloyd, R.E.; et al. Cryobanking of Viable Biomaterials: Implementation of New Strategies for Conservation Purposes. Mol. Ecol. 2009, 18, 1030–1033. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, T.; Sakai, Y.; Matsui, Y.; Sato, M.; Yamane, I.; Murao, I.; Stabenfeldt, G.H. Induced Ovulation in Cats Using Porcine Pituitary Gland Preparation During the Non-Breeding Season. Jpn. J. Vet. Sci. 1989, 51, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Tajima, H.; Yoshizawa, M.; Sasaki, S.; Yamamoto, F.; Narushima, E.; Tsutsui, T.; Funahashi, T.; Kusuda, S.; Doi, O.; Tateyama, Y.; et al. Intrauterine Insemination with Fresh Semen in Amur Leopard Cat (Pionailurus bengalensis eutilura) During Non-Breeding Season. J. Vet. Med. Sci. 2017, 79, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L.; Wildt, D.E.; Graham, L.H.; Byers, A.P.; Collins, L.; Barrett, S.; Howard, J.G. Natural Versus Chorionic Gonadotropin-Induced Ovarian Responses in the Clouded Leopard (Neofelis nebulosa) Assessed by Fecal Steroid Analysis. Biol. Reprod. 1995, 53, 93–102. [Google Scholar] [CrossRef]
- Tipkantha, W.; Thuwanut, P.; Maikeaw, U.; Thongphakdee, A.; Yapila, S.; Kamolnorranath, S.; Siriaroonrat, B.; Comizzoli, P.; Chatdarong, K. Successful Laparoscopic Oviductal Artificial Insemination in the Clouded Leopard (Neofelis nebulosa) in Thailand. J. Zoo Wildl. Med. 2017, 48, 804–812. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.H.; Kim, J.H.; Na, K.J. Characterization and Cryopreservation of Amur Leopard Cats (Prionailurus bengalensis euptilurus) Semen Collected by Urethral Catheterization. Theriogenology 2018, 119, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Takagi, Y.; Nakagawa, K.; Fujimoto, Y.; Hori, T.; Tsutsui, T. Artificial Intravaginal Insemination Using Fresh Semen in Cats. J. Vet. Med. Sci. 2000, 62, 1163–1167. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, T.; Tanaka, A.; Takagi, Y.; Nakagawa, K.; Fujimoto, Y.; Murai, M.; Anzai, M.; Hori, T. Unilateral Intrauterine Horn Insemination of Fresh Semen in Cats. J. Vet. Med. Sci. 2000, 62, 1241–1245. [Google Scholar] [CrossRef] [PubMed]
- De Haas van Dorsser, F.J.; Strick, J.A. Semen Characteristics and Sperm Morphology in the Arabian Leopard (Panthera pardus nimr) and How These Vary with Age and Season. Reprod. Fertil. Dev. 2005, 17, 675–682. [Google Scholar] [CrossRef]
- Brown, J.L.; Graham, L.H.; Wu, J.M.; Collins, D.; Swanson, W.F. Reproductive Endocrine Responses to Photoperiod and Exogenous Gonadotropins in the Pallas’ Cat (Otocolobus manul). Zoo Biol. 2002, 21, 347–364. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Concentration (mM) |
|---|---|
| NaCl | 100.0 |
| KCl | 8.0 |
| KH2PO4 | 1.0 |
| CaCl2-2H2O | 2.0 |
| MgSO4-7H2O | 1.0 |
| Glucose | 3.0 |
| L-Lactate | 6.0 |
| Pyruvate | 0.1 |
| NaHCO3 | 5.0 |
| HEPES | 20.0 |
| Glutamine | 1.0 |
| Taurine | 0.1 |
| NEAA | ×1 |
| BSA 1 | 4.0 mg/mL |
| Parameter | Date of Semen Collection | |||
|---|---|---|---|---|
| 28 December 2020 | 11 January 2021 | |||
| Male 1 | Male 2 | Male 1 | Male 2 | |
| Volume (μL) 1 | 8 + 60 | 7 + 50 | 10 + 25 | 10 + 35 |
| Concentration (×10⁶/mL) | 45 | 77 | 84 | 129 |
| Total sperm (×10⁶) | 2.7 | 3.9 | 2.9 | 5.8 |
| Motility (%) | 30 | 40 | 15 | 50 |
| Progressive motility (%) | 15 | 20 | 5 | 30 |
| Viability (%) | – 2 | – | – | 61 |
| Abnormality (%) | – | – | – | 47 |
| ID1 | ID2 | IBS0 | Kinship |
|---|---|---|---|
| Kitten | Female 2 | 0.01 | 0.22 |
| Kitten | Male 1 | 0.11 | −0.09 |
| Kitten | Male 2 | 0.01 | 0.23 |
| Male 1 | Female 2 | 0.09 | 0.04 |
| Male 2 | Female 2 | 0.05 | 0.13 |
| Male 2 | Male 1 | 0.08 | 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azumano, A.; Ueda, M.; Nomura, M.; Usui, M.; Ichinose, M.; Yanagawa, Y.; Kusuda, S.; Matsumoto, Y.; Murata, K. Successful Laparoscopic Oviductal Artificial Insemination in the Endangered Tsushima Leopard Cat (Prionailurus bengalensis euptilurus). Animals 2022, 12, 777. https://doi.org/10.3390/ani12060777
Azumano A, Ueda M, Nomura M, Usui M, Ichinose M, Yanagawa Y, Kusuda S, Matsumoto Y, Murata K. Successful Laparoscopic Oviductal Artificial Insemination in the Endangered Tsushima Leopard Cat (Prionailurus bengalensis euptilurus). Animals. 2022; 12(6):777. https://doi.org/10.3390/ani12060777
Chicago/Turabian StyleAzumano, Akinori, Miya Ueda, Mika Nomura, Masashi Usui, Midori Ichinose, Yojiro Yanagawa, Satoshi Kusuda, Yuki Matsumoto, and Koichi Murata. 2022. "Successful Laparoscopic Oviductal Artificial Insemination in the Endangered Tsushima Leopard Cat (Prionailurus bengalensis euptilurus)" Animals 12, no. 6: 777. https://doi.org/10.3390/ani12060777
APA StyleAzumano, A., Ueda, M., Nomura, M., Usui, M., Ichinose, M., Yanagawa, Y., Kusuda, S., Matsumoto, Y., & Murata, K. (2022). Successful Laparoscopic Oviductal Artificial Insemination in the Endangered Tsushima Leopard Cat (Prionailurus bengalensis euptilurus). Animals, 12(6), 777. https://doi.org/10.3390/ani12060777