
eDNA Metabarcoding Benchmarked towards Conventional Survey Methods in Amphibian Monitoring



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
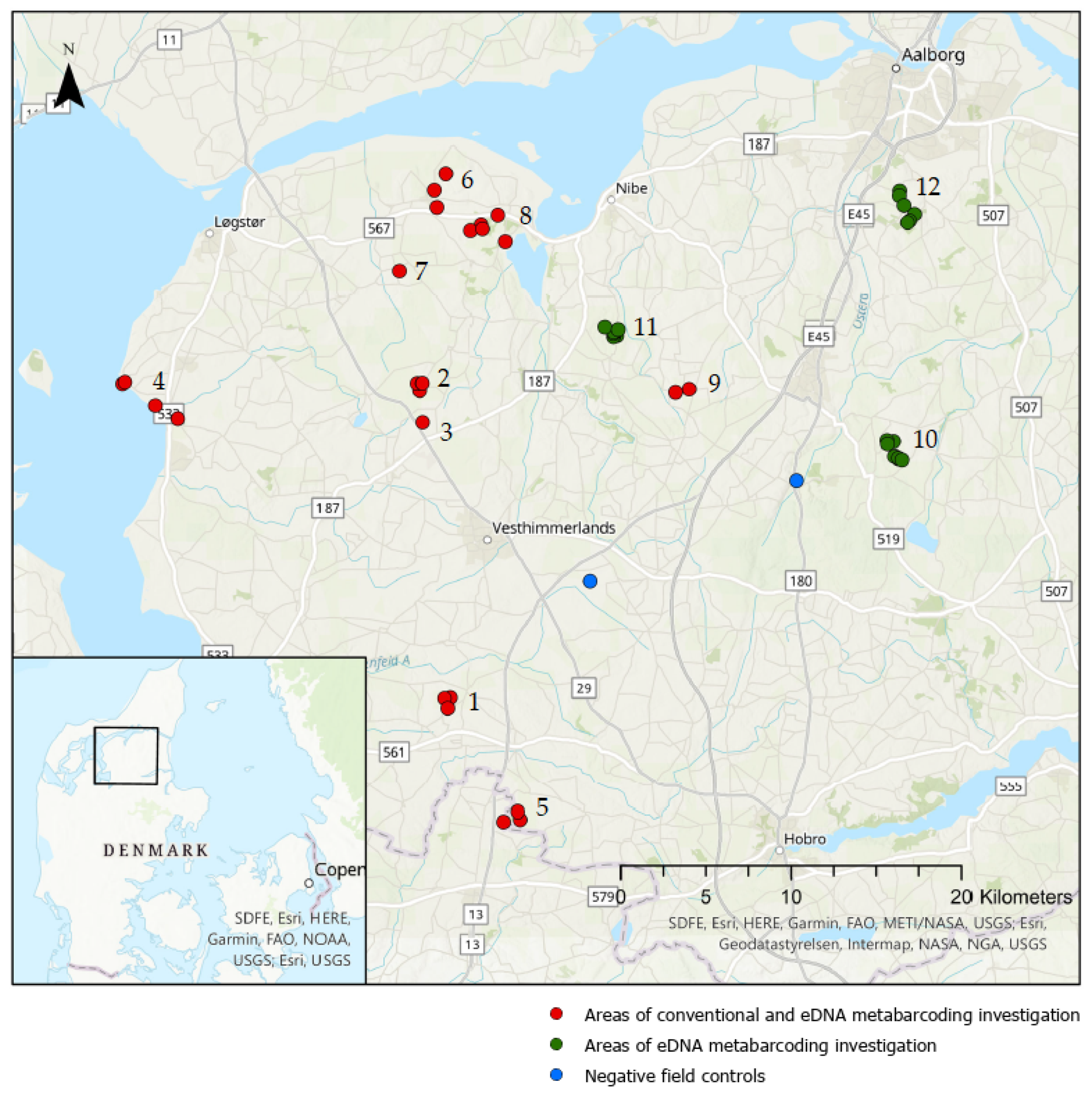
2.1. Areas of Investigation
2.2. Conventional Survey Methods
2.3. eDNA Sampling
2.4. Metabarcoding Laboratory and Library Analysis Procedure
2.5. Bioinformatics
2.6. Statistical Analysis
3. Results
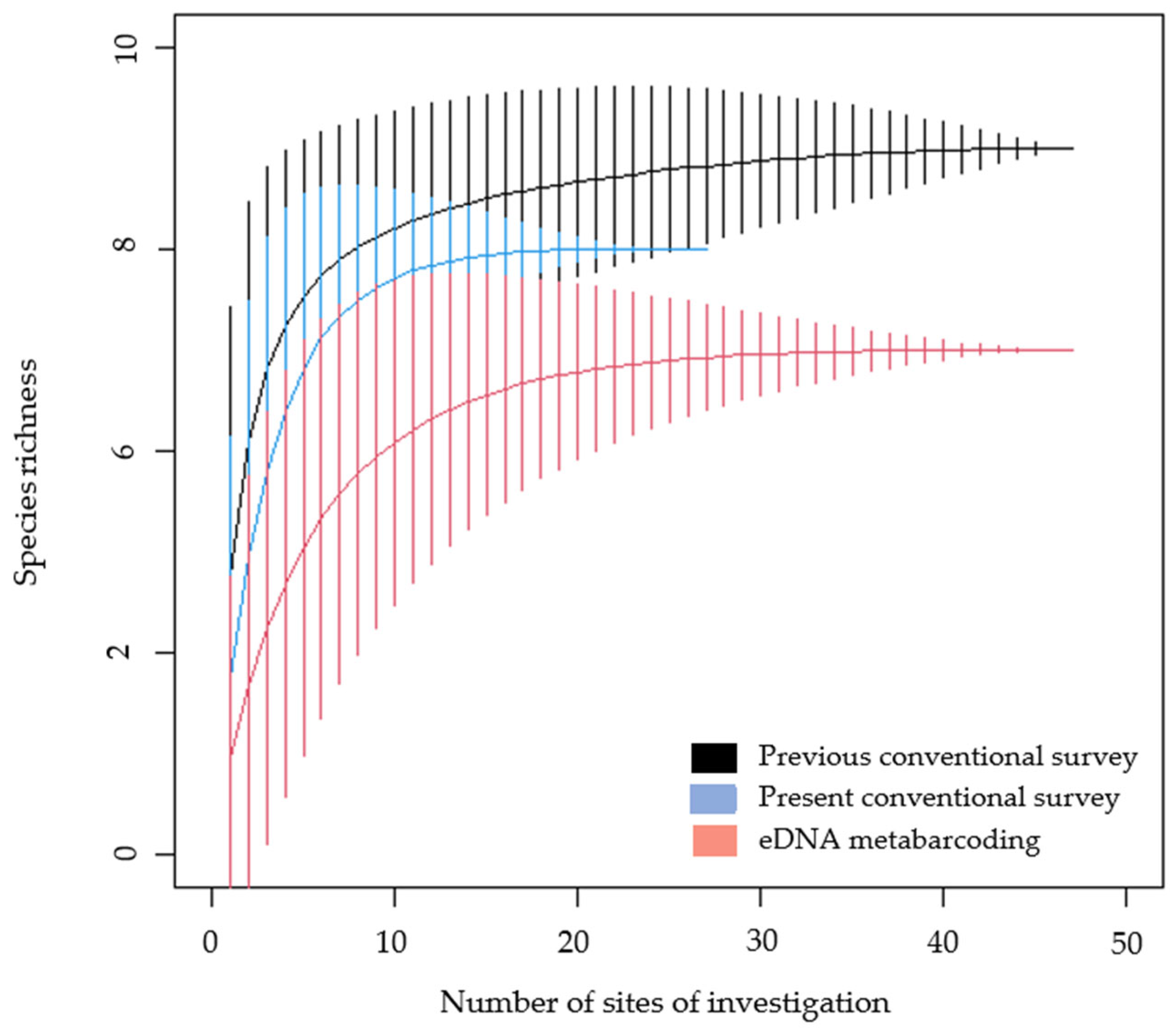
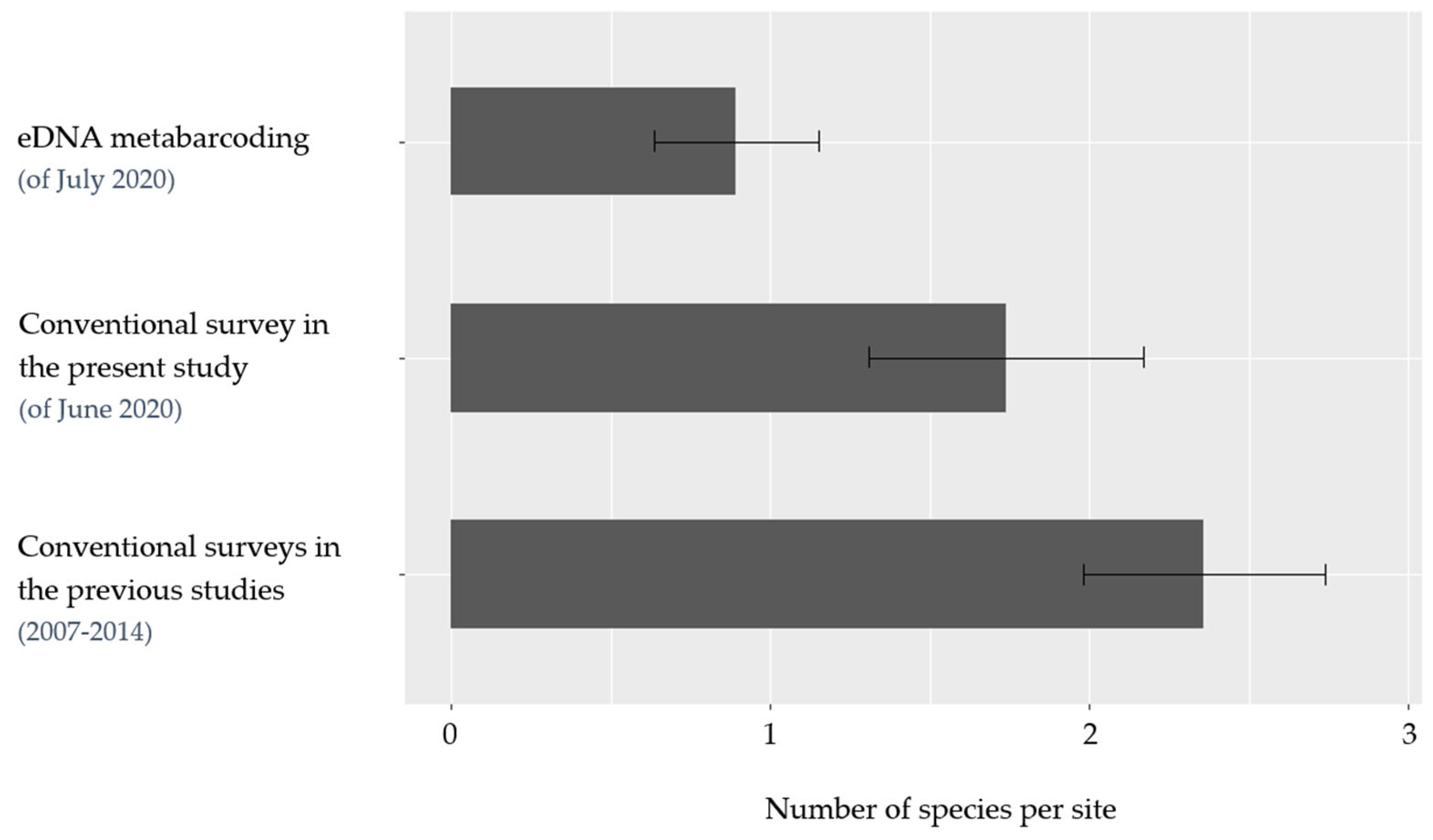
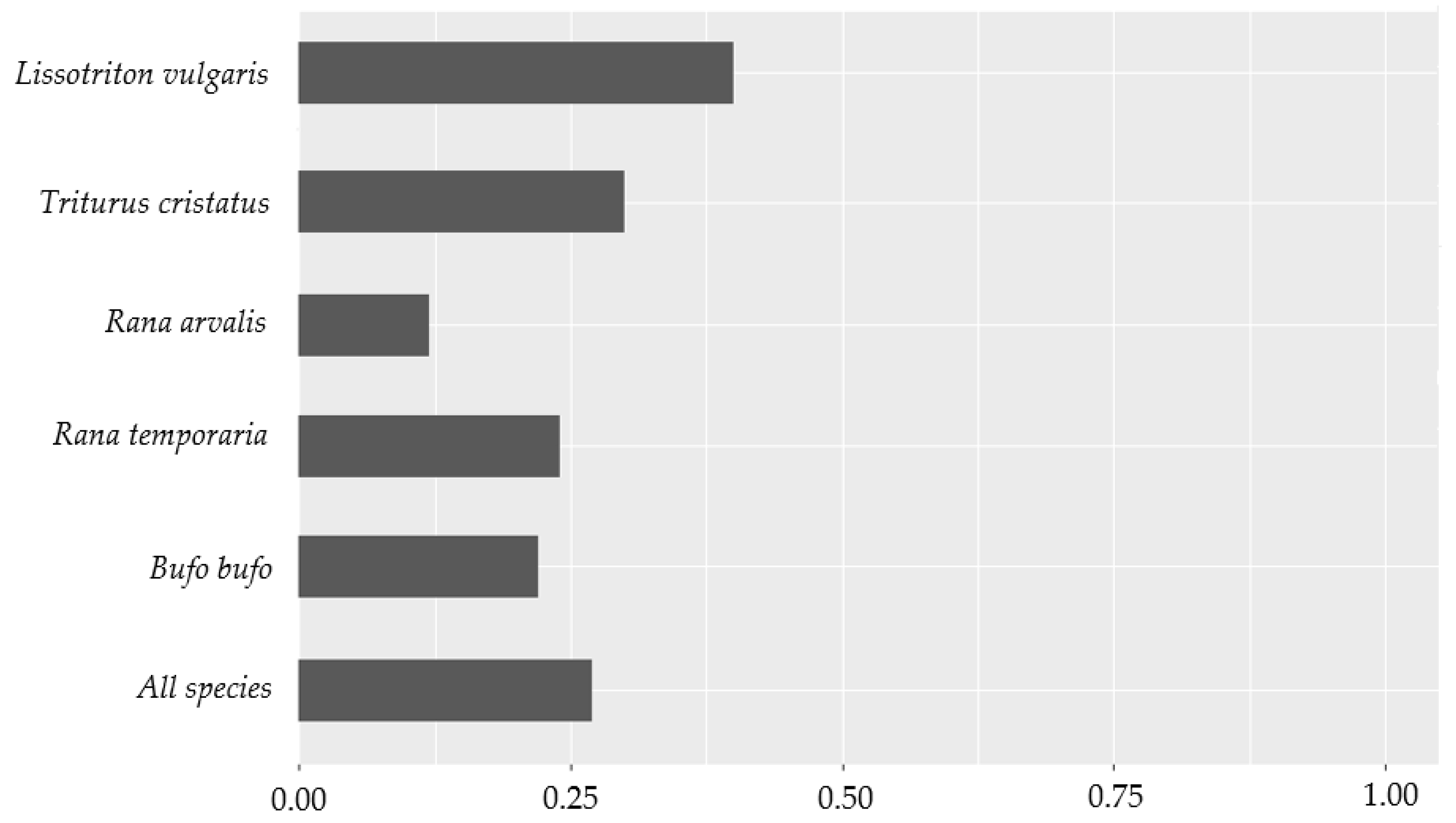
3.1. Species Richness and Average Detection
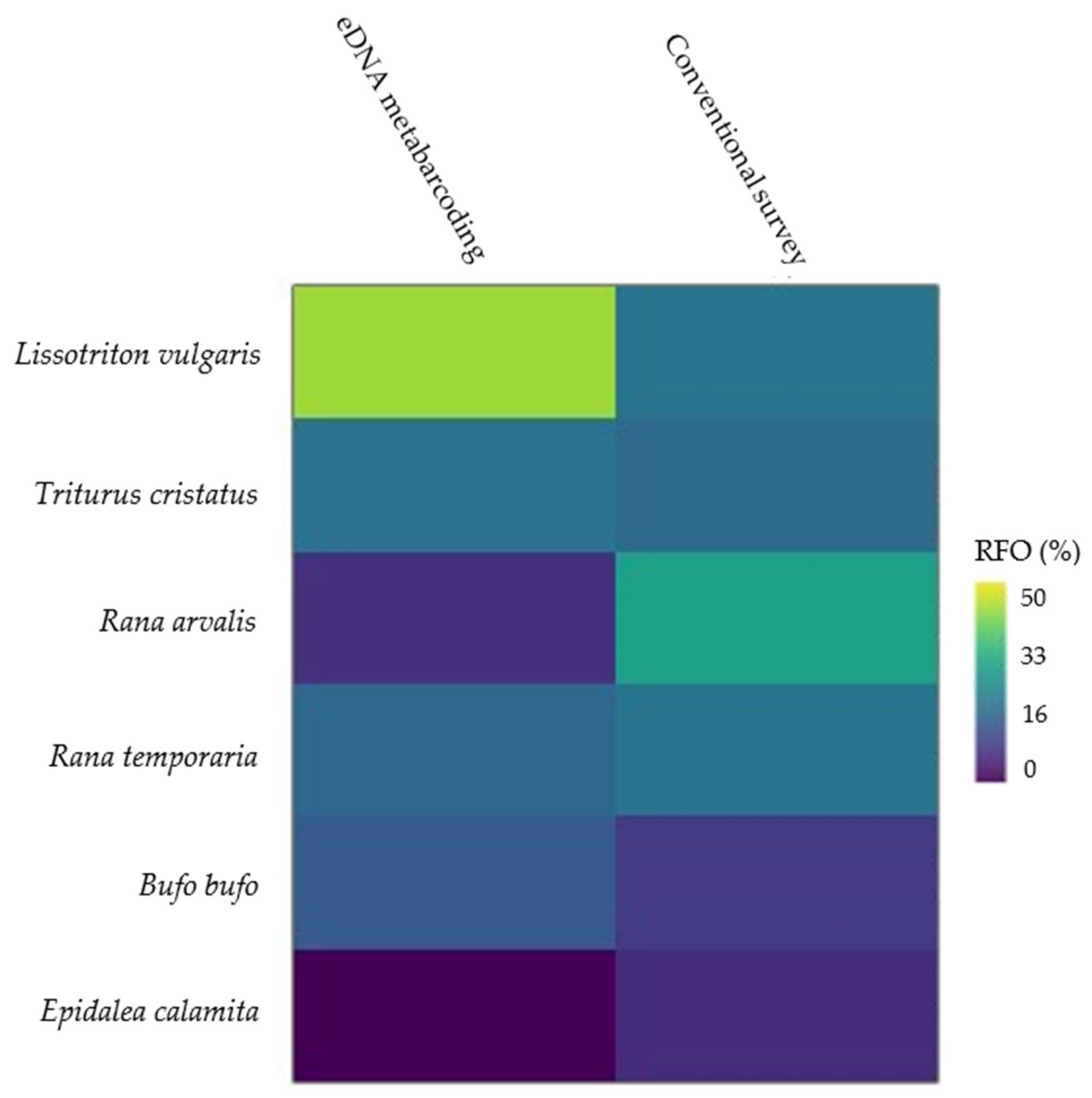
3.2. Species-Specific Detection
3.3. Similarity of Observations
4. Discussion
4.1. Assessment of Efficiency
4.2. A comparison with Historical Data
4.3. Similarity of Observations
4.4. Survey Methods in the Practice of Nature Monitoring
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collins, J.P.; Crump, M.L.; Lovejoy, T.E., III. Extinction in Our Times: Global Amphibian Decline; Oxford University Press, Inc.: Oxford, UK, 2009. [Google Scholar]
- Catenazzi, A. State of the World’s Amphibians. Annu. Rev. Environ. Resour. 2015, 40, 91–119. [Google Scholar] [CrossRef]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-1. 2021. Available online: https://www.iucnredlist.org (accessed on 18 May 2021).
- Søgaard, B.; Adrados, L.C.; Fog, K. Teknisk Anvisning Til Ekstensiv Overvågning; Aarhus Universitet. DCE—Nationalt Center for Miljø og Energi: Aarhus, Denmark, 2018; Volume A17, pp. 1–18.
- Pilliod, D.S.; Goldberg, C.S.; Arkle, R.S.; Waits, L.P. Estimating occupancy and abundance of stream amphibians using environmental DNA from filtered water samples. Can. J. Fish. Aquat. Sci. 2013, 70, 1123–1130. [Google Scholar] [CrossRef]
- Dejean, T.; Valentini, A.; Miquel, C.; Taberlet, P.; Bellemain, E.; Miaud, C. Improved detection of an alien invasive species through environmental DNA barcoding: The example of the American bullfrog Lithobates catesbeianus: Alien invasive species detection using eDNA. J. Appl. Ecol. 2012, 49, 953–959. [Google Scholar] [CrossRef]
- Eiler, A.; Löfgren, A.; Hjerne, O.; Nordén, S.; Saetre, P. Environmental DNA (eDNA) detects the pool frog (Pelophylax lessonae) at times when traditional monitoring methods are insensitive. Sci. Rep. 2018, 8, 5452. [Google Scholar] [CrossRef]
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163. [Google Scholar] [CrossRef]
- Bohmann, K.; Evans, A.; Gilbert, M.T.; Carvalho, G.R.; Creer, S.; Knapp, M.; Douglas, W.Y.; De Bruyn, M. Environmental DNA for wildlife biology and biodiversity monitoring. Trends Ecol. Evol. 2014, 29, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, C.S.; Turner, C.R.; Deiner, K.; Klymus, K.E.; Thomsen, P.F.; Murphy, M.A.; Spear, S.F.; McKee, A.; Oyler-McCance, S.J.; Cornman, R.S.; et al. Critical considerations for the application of environmental DNA methods to detect aquatic species. Methods Ecol. Evol. 2016, 7, 1299–1307. [Google Scholar] [CrossRef]
- Fog, K.; Schmedes, A.; Rosenørn, D. Nordens Padder og Krybdyr, 1st ed.; Gads Forlag: Copenhagen, Denmark, 1997. [Google Scholar]
- Holman, L.E.; Chng, Y.; Rius, M. How does eDNA decay affect metabarcoding experiments? Environ. DNA 2021, 4, 1–9. [Google Scholar] [CrossRef]
- Taberlet, P.; Bonin, A.; Zinger, L.; Coissac, E. Environmental DNA—For Biodiversity Research and Monitoring; Oxford University Press, Inc.: Oxford, UK, 2018. [Google Scholar]
- Harper, L.R.; Lawson Handley, L.; Hahn, C.; Boonham, N.; Rees, H.C.; Lewis, E.; Adams, I.P.; Brotherton, P.; Phillips, S.; Hänfling, B. Generating and testing ecological hypotheses at the pondscape with environmental DNA metabarcoding: A case study on a threatened amphibian. Environ. DNA 2020, 2, 184–199. [Google Scholar] [CrossRef] [Green Version]
- Valentini, A.; Taberlet, P.; Miaud, C.; Civade, R.; Herder, J.; Thomsen, P.F.; Bellemain, E.; Besnard, A.; Coissac, E.; Boyer, F.; et al. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol. Ecol. 2016, 25, 929–942. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, P.F.; Kielgast, J.; Iversen, L.L.; Wiuf, C.; Rasmussen, M.; Gilbert, M.T.P.; Orlando, L.; Willerslev, E. Monitoring endangered freshwater biodiversity using environmental DNA. Mol. Ecol. 2012, 21, 2565–2573. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.W.; Therkildsen, O.R. Overvågning af Bilag II- og IV-Arter Baseret på eDNA—Muligheder og Begrænsninger; Aarhus Universitet, DCE—Nationalt Center for Miljø og Energi: Aarhus, Denmark, 2020.
- Winding, A.; Bang-Andreasen, T.; Hansen, L.H.; Panitz, F.; Krogh, P.H.; Krause-Jensen, D.; Stæhr, P.; Nicolaisen, M.; Hendriksen, N.B.; Sapkota, R.; et al. eDNA in Environmental Monitoring; Technical Report 133; Aarhus Universitet, DCE—Nationalt Center for Miljø og Energi: Aarhus, Denmark, 2019.
- Broadhurst, H.A.; Gregory, L.M.; Bleakley, E.K.; Perkins, J.C.; Lavin, J.V.; Bolton, P.; Browett, S.S.; Howe, C.V.; Singleton, N.; Tansley, D.; et al. Mapping differences in mammalian distributions and diversity using environmental DNA from rivers. Sci. Total Environ. 2021, 801, 149724. [Google Scholar] [CrossRef] [PubMed]
- Senapati, D.; Bhattacharya, M.; Kar, A.; Chini, D.S.; Das, B.K.; Patra, B.C. Environmental DNA (eDNA): A Promising Biological Survey Tool for Aquatic Species Detection. Proc. Zool. Soc. 2019, 72, 211–228. [Google Scholar] [CrossRef]
- Coble, A.A.; Flinders, C.A.; Homyack, J.A.; Penaluna, B.E.; Cronn, R.C.; Weitemier, K. eDNA as a tool for identifying freshwater species in sustainable forestry: A critical review and potential future applications. Sci. Total Environ. 2019, 649, 14. [Google Scholar] [CrossRef]
- Li, W.; Hou, X.; Xu, C.; Qin, M.; Wang, S.; Wei, L.; Wang, Y.; Liu, X.; Li, Y. Validating eDNA measurements of the richness and abundance of anurans at a large scale. J. Anim. Ecol. 2021, 90, 1466–1479. [Google Scholar] [CrossRef]
- Browett, S.S.; O’Meara, D.B.; McDevitt, A.D. Genetic tools in the management of invasive mammals: Recent trends and future perspectives. Mammal. Rev. 2020, 50, 200–210. [Google Scholar] [CrossRef]
- Andersen, L.W.; Søgaard, B.; Johansson, L.S.; Wiberg-Larsen, P. Anvendelse af eDNA-Metoder i NOVANA-Artsovervågningen—Muligheder og Begrænsninger; Aarhus Universitet, DCE—Nationalt Center for Miljø og Energi: Aarhus, Denmark, 2012.
- Neergaard, R.S. Afrapportering af Vandhulsundersøgelser i Rebild Kommune i 2012; Amphi Consult: Slagelse, Denmark, 2013. [Google Scholar]
- Danmarks Miljøportal, Danmarks Naturdata. Available online: https://naturdata.miljoeportal.dk/ (accessed on 25 March 2021).
- Machida, R.; Leray, M.; Knowlton, N. MIDORI Reference Database Based on the Genback v241. 2020. Available online: http://www.reference-midori.info/index.html (accessed on 27 May 2021).
- NatureMetrics. Aquatic eDNA Sampling Instructions with NatureMetrics. Available online: https://www.naturemetrics.co.uk/wildlife-services/edna-from-water/#mbc-amphibians (accessed on 1 May 2021).
- Miya, M.; Sato, Y.; Fukunaga, T.; Sado, T.; Poulsen, J.Y.; Sato, K.; Minamoto, T.; Yamamoto, S.; Yamanaka, H.; Araki, H.; et al. MiFish, a set of universal PCR primers for metabarcoding environmental DNA from fishes: Detection of more than 230 subtropical marine species. R. Soc. Open Sci. 2015, 2, 150088. [Google Scholar] [CrossRef] [Green Version]
- Riaz, T.; Shehzad, W.; Viari, A.; Pompanon, F.; Taberlet, P.; Coissac, E. ecoPrimers: Inference of new DNA barcode markers from whole genome sequence analysis. Nucleic Acids Res. 2011, 39, e145. [Google Scholar] [CrossRef]
- Kelly, R.P.; Port, J.A.; Yamahara, K.M.; Crowder, L.B. Using Environmental DNA to Census Marine Fishes in a Large Mesocosm. PLoS ONE 2014, 9, e86175. [Google Scholar] [CrossRef] [Green Version]
- Yashiro, E. AmpProc 5.1.0.Beta2.12.1. 2021. Available online: https://github.com/eyashiro/AmpProc (accessed on 27 May 2021).
- Edgar, R.C. UNOISE2: Improved error-correction for Illumina 16S and ITS amplicon sequencing. Bioinformatics 2016. preprint. [Google Scholar] [CrossRef] [Green Version]
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M. ampvis2: An R Package to analyse and visualise 16S rRNA amplicon data. Bioinformatics 2018. preprint. [Google Scholar] [CrossRef] [Green Version]
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 13 April 2021).
- Jari Oksanen, F.; Guillaume, B.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P. Package ‘Vegan’—Community Ecology Package. 2020. Available online: https://cran.r-project.org/ (accessed on 30 April 2021).
- Wickham, H. ggplot2. Elegant Graphics for Data Analysis; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; Volume 2. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Sørensen, T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analysis of the vegetation on danish commons. Biol. Skr. Bind V Det K. Dan. Vidensk. Selsk. 1948, 5, 1–34. [Google Scholar]
- Danmarks Meteorologiske Institut. Året 2020; Danmarks Meteorologiske Institut: København Ø, Denmark, 2021.
- Evans, N.T.; Olds, B.P.; Renshaw, M.A.; Turner, C.R.; Li, Y.; Jerde, C.L.; Mahon, A.R.; Pfrender, M.E.; Lamberti, G.A.; Lodge, D.M. Quantification of mesocosm fish and amphibian species diversity via environmental DNA metabarcoding. Mol. Ecol. Resour. 2016, 16, 29–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blom, L.B. The Use of Environmental DNA (eDNA) in Biological Monitoring of the Natterjack Toad (Epidalea calamita); Ekoll AB and Lunds University, Department of Biology: Lund, Sweden, 2018. [Google Scholar]
- Reyne, M.; Naaum, A.M.; Marnell, F.; Reid, N.; Helyar, S.J. Development and validation of a quantitative qPCR assay for detecting Natterjack toad (Epidalea calamita) eDNA samples. Conserv. Genet. Resour. 2021, 13, 319–322. [Google Scholar] [CrossRef]
- Moeslund, J.E.; Nygaard, B.; Ejrnæs, R.; Bell, N.; Bruun, L.D.; Bygebjerg, R.; Carl, H.; Damgaard, J.; Dylmer, E.; Elmeros, M.; et al. Den Danske Rødliste; Aarhus Universitet, DCE—Nationalt Center for Miljø og Energi: Aarhus, Denmark, 2019. Available online: www.redlist.au.dk (accessed on 23 March 2021).
- European Environment Agency. State of Nature in the EU: Results from Reporting Under the Nature Directives 2013–2018. Publications Office. 2020. Available online: https://data.europa.eu/doi/10.2800/705440 (accessed on 1 May 2021).
- Temple, H.J.; Cox, N.A. European Red List of Amphibians; IUCN; Office for Official Publications of the European Communities: Gland, Switzerland, 2009. [Google Scholar]
- Søgaard, B.; Asferg, T.; Søgaard, B. Håndbog om Dyrearter på Habitatdirektivets Bilag IV: Til Brug i Administration og Planlægning; Danmarks Miljøundersøgelser: Aarhus, Denmark, 2007.
- Turner, C.R.; Barnes, M.; Xu, C.C.Y.; Jones, S.E.; Jerde, C.L.; Lodge, D.M. Particle size distribution and optimal capture of aqueous macrobial eDNA. Methods Ecol. Evol. 2014, 5, 676–684. [Google Scholar] [CrossRef] [Green Version]
- Mize, E.; Erickson, R.; Merkes, C.M.; Berndt, N.; Bockrath, K.; Credico, J.; Grueneis, N.; Merry, J.; Mosel, K.; Tuttle-Lau, M.; et al. Refinement ofeDNAas an early monitoring tool at the landscape-level: Study design considerations. Ecol. Appl. 2019, 29, e01951. [Google Scholar] [CrossRef]
- Roussel, J.-M.; Paillisson, J.-M.; Treguier, A.; Petit, E. The downside of eDNA as a survey tool in water bodies. J. Appl. Ecol. 2015, 52, 823–826. [Google Scholar] [CrossRef]
- Biggs, J.; Ewald, N.; Valentini, A.; Gaboriaud, C.; Dejean, T.; Griffiths, R.A.; Foster, J.; Wilkinson, J.W.; Arnell, A.; Brotherton, P.; et al. Using eDNA to develop a national citizen science-based monitoring programme for the great crested newt (Triturus cristatus). Biol. Conserv. 2015, 183, 19–28. [Google Scholar] [CrossRef]
- Ray, A.M.; Hossack, B.R.; Gould, W.R.; Patla, D.A.; Spear, S.F.; Klaver, R.W.; Bartelt, P.E.; Thoma, D.P.; Legg, K.L.; Daley, R.; et al. Multi-species amphibian monitoring across a protected landscape: Critical reflections on 15 years of wetland monitoring in Grand Teton and Yellowstone national parks. Ecol. Indic. 2022, 135, 108519. [Google Scholar] [CrossRef]
- Smith, M.M.; Goldberg, C.S. Occupancy in dynamic systems: Accounting for multiple scales and false positives using environmental DNA to inform monitoring. Ecography 2020, 43, 376–386. [Google Scholar] [CrossRef]
- Lynggaard, C.; Nielsen, L.D.S.B.; Santos-Bay, L.; Gastauer, M.; Oliveira, G.; Bohmann, K. Vertebrate diversity revealed by metabarcoding of bulk arthropod samples from tropical forests. Environ. DNA 2019, 1, 329–341. [Google Scholar] [CrossRef]
- Valsecchi, E.; Bylemans, J.; Goodman, S.J.; Lombardi, R.; Carr, I.; Castellano, L.; Galimberti, A.; Galli, P. Novel universal primers for metabarcoding environmental DNA surveys of marine mammals and other marine vertebrates. Environ. DNA 2020, 2, 460–476. [Google Scholar] [CrossRef]
- Harper, L.R.; Handley, L.L.; Hahn, C.; Boonham, N.; Rees, H.C.; Lewis, E.; Adams, I.P.; Brotherton, P.; Phillips, S.; Hänfling, B. Ground-truthing environmental DNA (eDNA) metabarcoding for ecological hypothesis testing at the pondscape. bioRxiv 2018, 278309. [Google Scholar] [CrossRef] [Green Version]
- Hansen, L.S.; Svenningsen, A.K. Betydningen af Hydrologisk Genopretningfor Bilag IV-Padder på Skagen Odde; Department of Chemistry and Bioscience, Aalborg University: Aalborg, Denmark, 2018. [Google Scholar]
- Boyce, M.; Rushton, S.; Lynam, T. Does modelling have a role in conservation? In Key Topics in Conservation Biology; Wiley: Hoboken, NJ, USA, 2007; pp. 134–144. [Google Scholar]
- Préau, C.; Bertrand, R.; Trochet, A.; Isselin-Nondedeu, F. Modeling potential distributions of three European amphibian species comparing ENFA and MaxEnt. Herpetol. Conserv. Biol. 2018, 13, 15. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Svenningsen, A.K.N.; Pertoldi, C.; Bruhn, D. eDNA Metabarcoding Benchmarked towards Conventional Survey Methods in Amphibian Monitoring. Animals 2022, 12, 763. https://doi.org/10.3390/ani12060763
Svenningsen AKN, Pertoldi C, Bruhn D. eDNA Metabarcoding Benchmarked towards Conventional Survey Methods in Amphibian Monitoring. Animals. 2022; 12(6):763. https://doi.org/10.3390/ani12060763
Chicago/Turabian StyleSvenningsen, Anne Katrine Nørgaard, Cino Pertoldi, and Dan Bruhn. 2022. "eDNA Metabarcoding Benchmarked towards Conventional Survey Methods in Amphibian Monitoring" Animals 12, no. 6: 763. https://doi.org/10.3390/ani12060763
APA StyleSvenningsen, A. K. N., Pertoldi, C., & Bruhn, D. (2022). eDNA Metabarcoding Benchmarked towards Conventional Survey Methods in Amphibian Monitoring. Animals, 12(6), 763. https://doi.org/10.3390/ani12060763