
Effect of Intramuscularly Administered Oxytetracycline or Enrofloxacin on Vancomycin-Resistant Enterococci, Extended Spectrum Beta-Lactamase- and Carbapenemase-Producing Enterobacteriaceae in Pigs

 ,
,  , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sampling
2.3. Bacterial Isolation and Identification
2.4. Statistical Analysis
2.5. Ethical Considerations
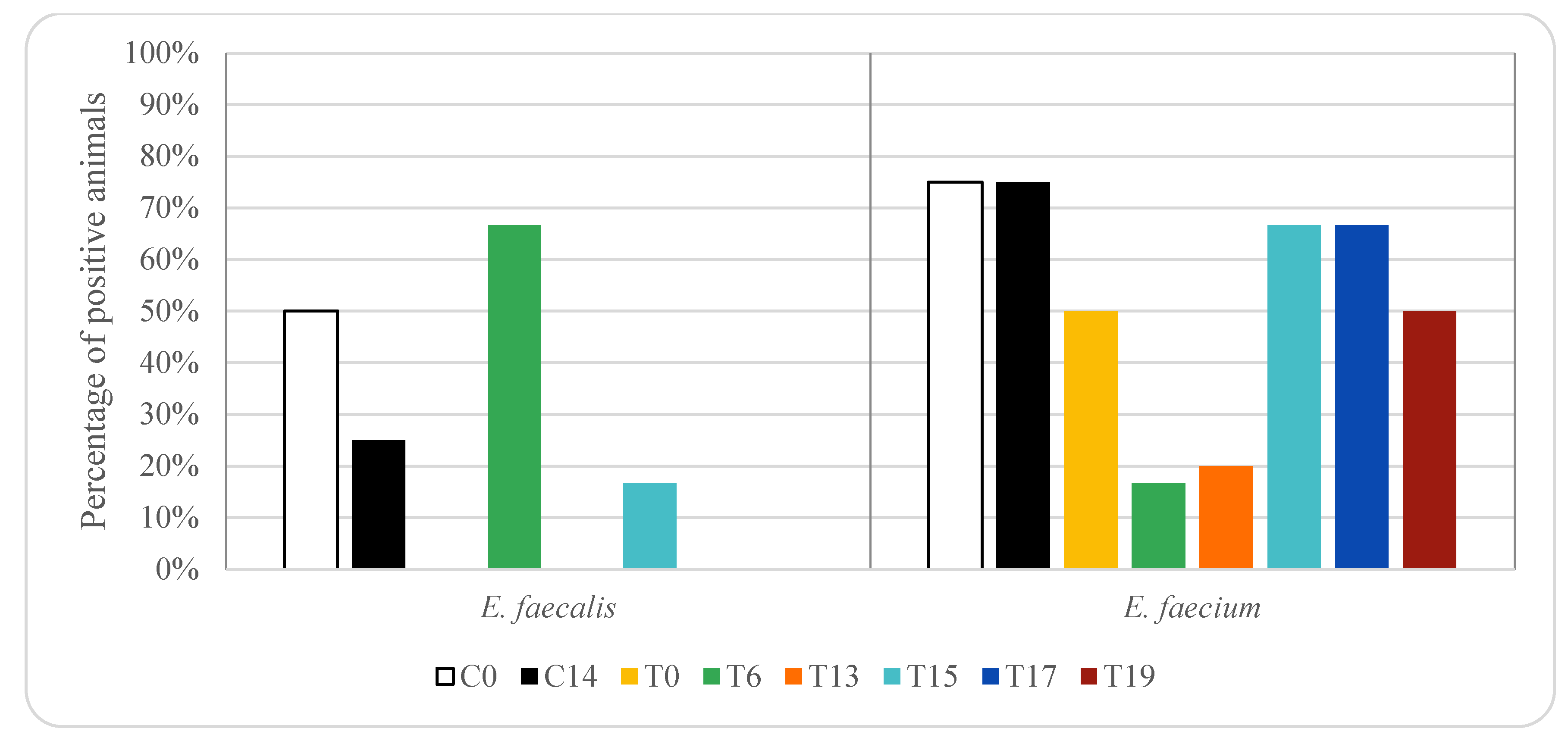
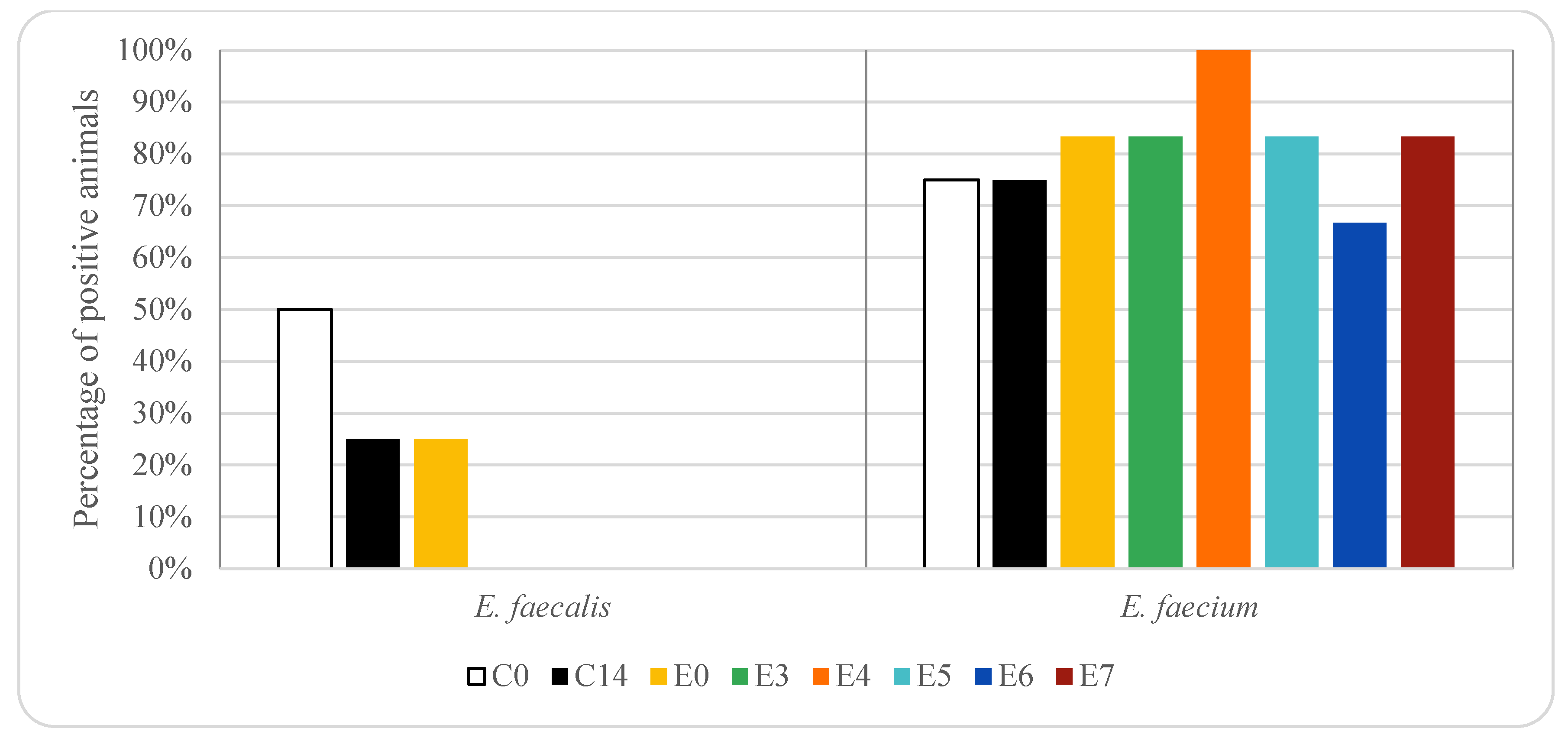
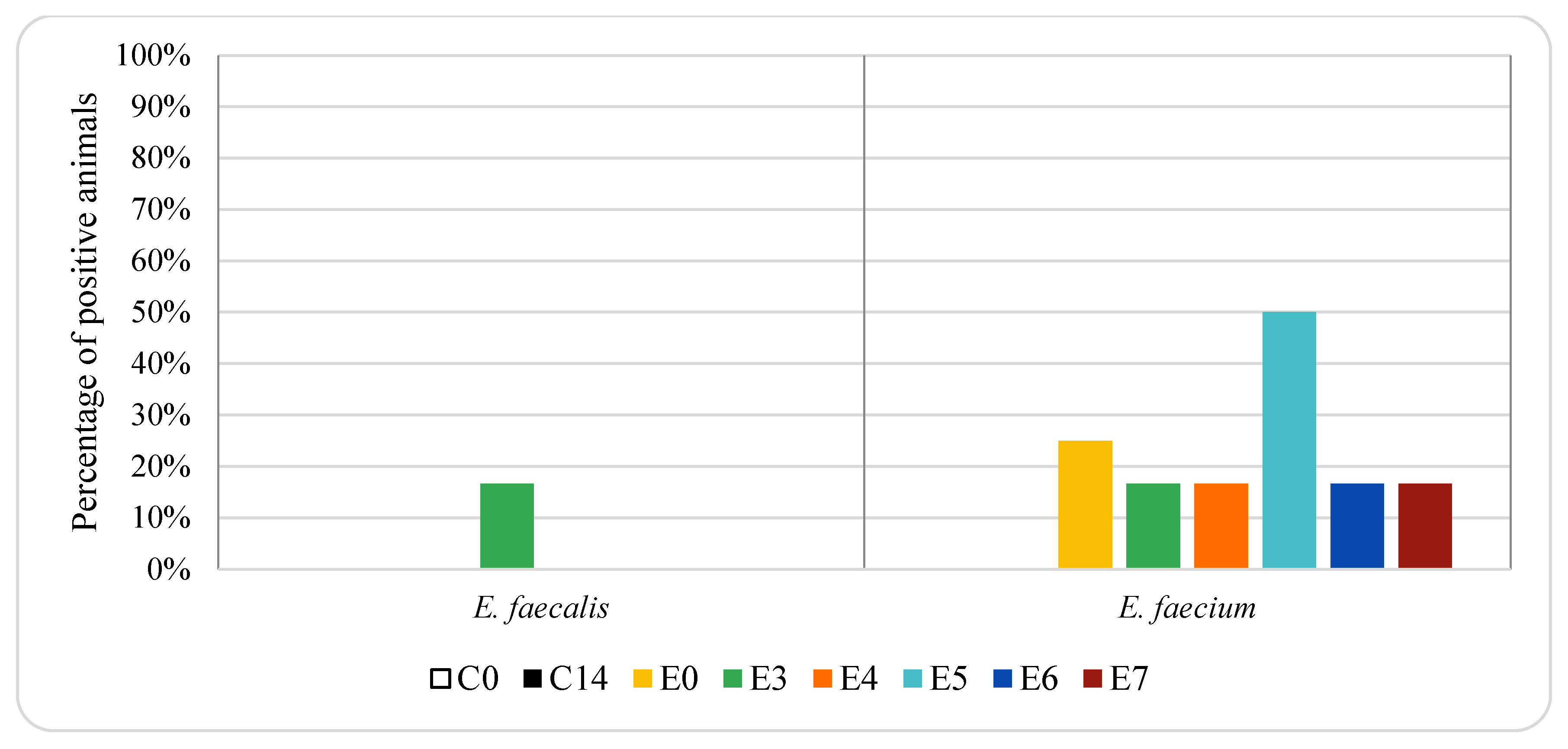
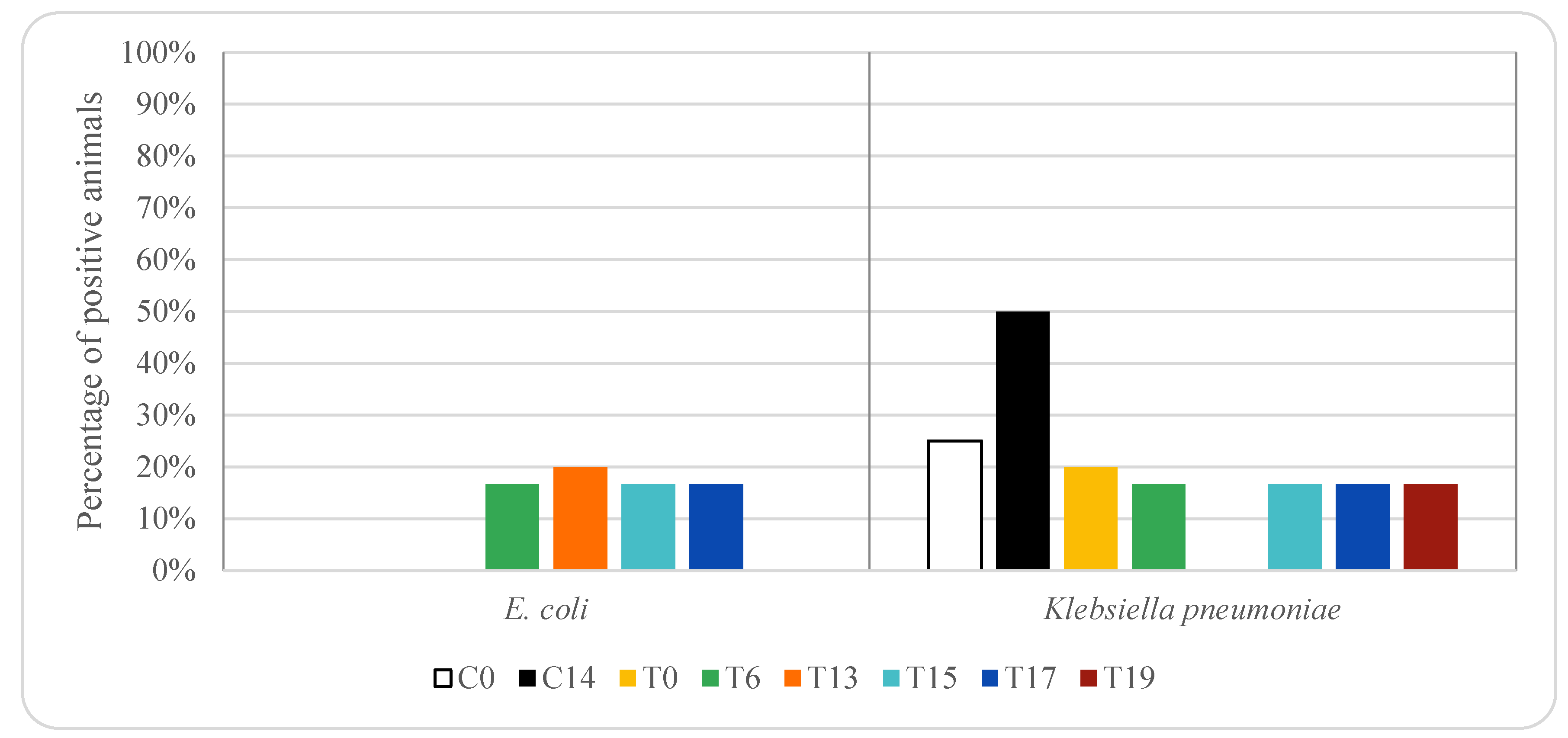
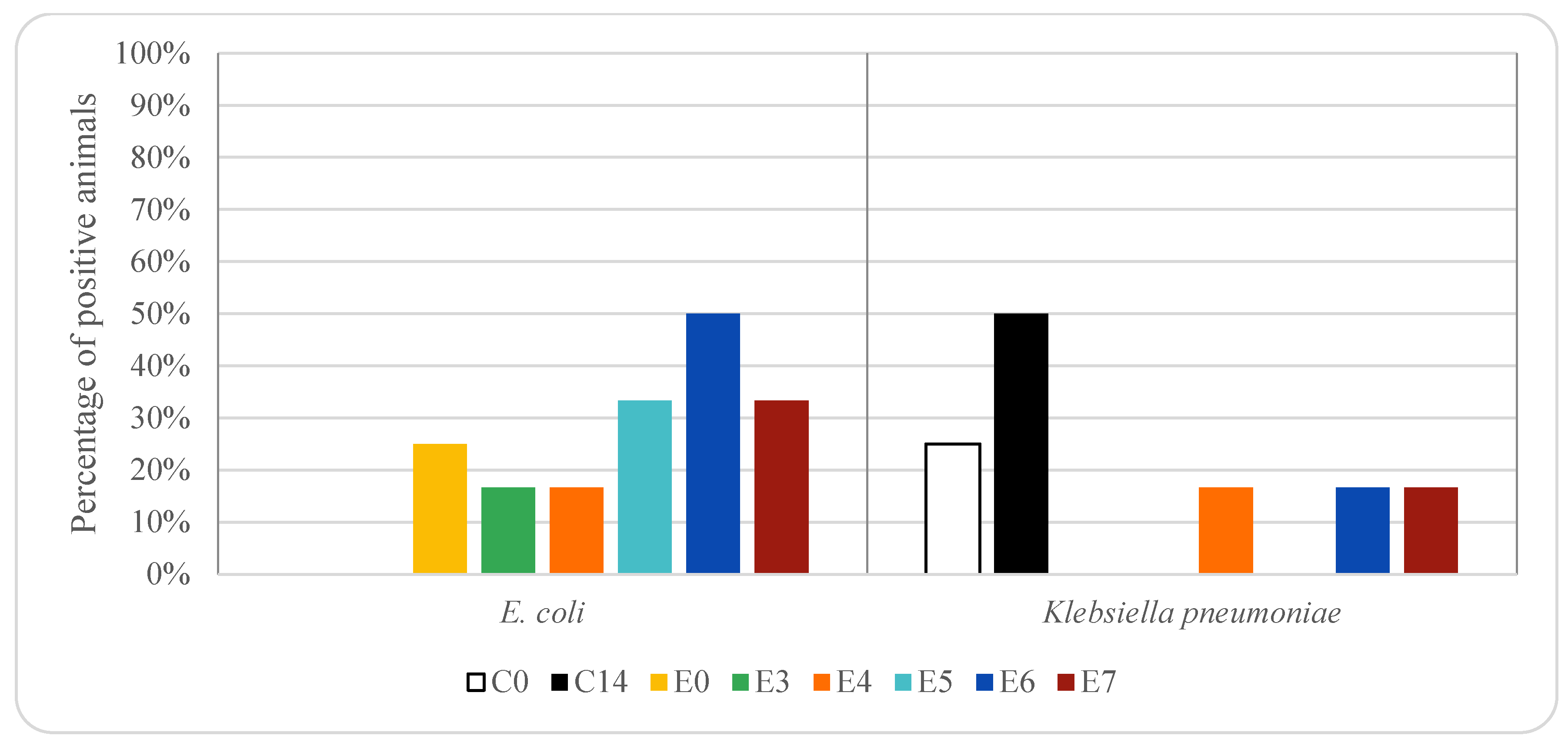
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Neill, J. Antimicrobial Resistance: Tackling a Crisis for the Health and Wealth of Nations. London: Review on Antimicrobial Resistance. 2014. Available online: https://amr-review.org/sites/default/files/AMR%20Review%20Paper%20-%20Tacklng%20a%20crisis%20for%20the%20health%20and%20wealth%20of%20nations_1.pdf (accessed on 2 November 2021).
- Wall, B.A.; Mateus, A.; Marshall, L.; Pfeiffer, D.; Lubroth, J.; Ormel, H.J.; Otto, P.; Patriarchi, A.; Food and Agriculture Organization of the United Nations. Drivers, Dynamics and Epidemiology of Antimicrobial Resistance in Animal Production; Food and Agriculture Organization (FAO): Rome, Italy, 2016. [Google Scholar]
- World Health Organization. Antibacterial Agents in Clinical Development: An Analysis of The Antibacterial Clinical Development Pipeline; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- World Health Organization. Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- World Organization for Animal Health (OIE): OIE List of Antimicrobials of Veterinary Importance. 2019. Available online: http://www.oie.int (accessed on 18 November 2021).
- Amusi, J.; Tamara, L.; Horton, R.; Winkler, A.S. Reconnecting for our future: The Lancet One Health commission. Lancet 2020, 395, 1469–1471. [Google Scholar] [CrossRef]
- Colligon, P.J.; McEwen, S.A. One Health. Its importance in helping to better control antimicrobial resistance. Trop. Med. Infect. Dis. 2019, 4, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergšpica, I.; Kaprou, G.; Alexa, E.A.; Prieto, M.; Alvarez-Ordóñez, A. Extended spectrum β-lactamase (ESBL) Escherichia coli in pigs and pork meat in European Union. Antibiotics 2020, 9, 678. [Google Scholar] [CrossRef]
- Fournier, C.; Nordmann, P.; Pittet, O.; Poirel, L. Does an antibiotic stewardship applied in a pig farm lead to low ESBL prevalence? Antibiotics 2021, 10, 574. [Google Scholar] [CrossRef] [PubMed]
- Dandachi, I.; Chabou, S.; Daoud, Z.; Rolain, J.M. Prevalence and emergence of extended-spectrum cephalosporin, carbapenem and colistin-resistant Gram negative bacteria of animal origin in the Mediterranean basin. Front. Microbiol. 2018, 9, 2299. [Google Scholar] [CrossRef] [PubMed]
- Unal, N.; Bal, E.; Kapagoz, A.; ALtun, B.; Kozag, N. Detection of vancomycin-resistant enterococci in samples from broiler flocks and houses. Acta Vet. Hung. 2020, 68, 117–122. [Google Scholar] [CrossRef]
- Fournier, S.; Brun-Buisson, C.; Jarlier, V. Twenty years of antimicrobial resistance control programme in a regional multi hospital institution, with focus on emerging bacteria (VRE and CPE). Antimicrob. Resist. Infect. Control 2012, 1, 9. [Google Scholar] [CrossRef] [Green Version]
- Adler, A.; Katz, D.E.; Marchaim, D. The continuing plague of extended-spectrum beta-lactamase-producing Enterobacteriaceae infections. Infect. Dis. Clin. North Am. 2016, 30, 347–375. [Google Scholar] [CrossRef]
- Madec, J.Y.; Haenni, M.; Nordmann, P.; Poirel, L. Extended-spectrum beta-lactamase/AmpC- and carbapenemase-producing Enterobacteriaceae in animals: A threat for humans? Clin. Microbiol. Infect. 2017, 23, 826–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2018/2019. EFSA J. 2021, 19, e06490. [Google Scholar]
- Novais, C.; Freitas, A.R.; Silveira, E.; Antunes, P.; Silva, R.; Coque, T.M.; Peixe, L. Spread of multidrug-resistant Enterococcus to animals and humans: An underestimated role for the pig farm environment. J. Antimicrob. Chemother. 2013, 68, 2746–2754. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O. Vancomycin resistant enterococci in farm animals. Occurrence and importance. Infect. Ecol. Epidemiol. 2012, 2, 16959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gastmeier, P.; Schröder, C.; Behnke, M.; Meyer, E.; Geffers, C. Dramatic increase in vancomycin-resistant enterococci in Germany. J. Antimicrob. Chemother. 2014, 69, 1660–1664. [Google Scholar] [CrossRef] [Green Version]
- Hammerum, A.M. Enterococci of animal origin and their significance for public health. Clin. Microbiol. Infect. 2012, 18, 619–625. [Google Scholar] [CrossRef] [PubMed]
- EMA (European Medicine Agency). Categorisation of Antibiotics for Use in Animals for Prudent and Responsible Use. 2020. Available online: https://www.ema.europa.eu/en/documents/report/infographic-categorisation-antibiotics-use-animals-prudent-responsible-use_en.pdf (accessed on 27 April 2021).
- Tyson, G.H.; Nyirabahizi, E.; Crarey, E.; Kabera, C.; Lam, C.; Rice-Trujillo, C.; McDermott, P.F.; Tate, H. Prevalence and antimicrobial resistance of enterococci isolated from world retail meats in the United States, 2002 to 2014. Appl. Environ. Microbiol. 2018, 84, e01902-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galler, H.; Luxner, J.; Petternel, C.; Reinthaler, F.F.; Habib, J.; Haas, D.; Kittinger, C.; Pless, P.; Feierl, G.; Zarfel, G. Multiresistant bacteria isolated from intestinal faeces of farm animals in Austria. Antibiotics 2021, 10, 466. [Google Scholar] [CrossRef]
- Doi, Y.; Iovleva, A.; Bonomo, R.A. The ecology of extended-spectrum B-lactamases (ESBLs) in the developed world. J. Travel Med. 2017, 24, S44–S51. [Google Scholar] [CrossRef]
- Tamta, S.; Vonodh-Kumar, O.R.; Pruthvishree, B.S.; Karthikeyan, R.; Ramkumar, R.R.; Chethan, G.E.; Dubal, Z.B.; Sinha, D.K.; Singh, B.R. Faecal carriage of extended spectrum beta-lactamase (ESBL) and New Delhi metallobeta-lactamase(NDM) producing Escherichia coli between piglets and pig farmworkers. Comp. Immunol. Microbiol. Infect. Dis. 2020, 73, 101564. [Google Scholar] [CrossRef]
- Cantas, L.; Suer, K.; Guler, E.; Imir, T. High emergence of ESBL producing E. coli cystitis: Time to get smarter in Cyprus. Front. Microbiol. 2015, 6, 1446. [Google Scholar] [CrossRef]
- Stefani, S.; Giovanelli, I.; Anacarso, I.; Condo, C.; Messi, P.; de Niederhausern, S.; Bondi, M.; Iseppi, R.; Sabia, C. Prevalence and characterization of extended-spectrum beta-lactamase-producing Enterobacteriaceae in food-producing animals in Northern Italy. New Microbiol. 2014, 37, 551–555. [Google Scholar]
- Nordmann, P. Carbapenemase-producing Enterobacteriaceae: Overview of a major public health challenge. Méd. Mal. Infect. 2014, 44, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Food Safety Authority. Scientific Opinion on Carbapenem resistance in food animal ecosystems. EFSA J. 2013, 11, 3501. [Google Scholar]
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Köck, R.; Daniels-Haardt, I.; Becker, K.; Mellmann, A.; Friedrich, A.W.; Mevius, D.; Schwarz, S.; Jurke, A. Carbapen Carbapenem-resistant Enterobacterales in wildlife, food-producing, and companion animals: A systematic review. Clin. Microbiol. Infect. 2018, 24, 1241–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.C.; Chong, C.W.; Teh, C.S.J.; Ooi, P.T.; Thong, K.L. Occurrence of virulent multidrug resistant Enterococcus faecalis and Enterococcus faecium in the pigs, farmers and farm environments in Malaysia. PeerJ 2018, 6, e5353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aasmäe, B.; Häkkinen, L.; Kaartt, T.; Kalmus, P. Antimicrobial resistance of Escherichia coli and Enterococcus spp. isolated from Estonian cattle and swine from 2010 to 2015. Acta Vet. Scand. 2019, 61, 5. [Google Scholar] [CrossRef] [PubMed]
- Gruel, G.; Sellin, A.; Riveiro, H.; Pot, M.; Breurec, S.; Guyomard-Rabenirina, S.; Talarmin, A.; Ferdinand, S. Antimicrobial use and resistance in Escherichia coli from healthy food-producing animals in Guadeloupe. BMC Vet. Res. 2021, 17, 116. [Google Scholar] [CrossRef]
- De Smet, J.; Boyen, F.; Croubels, S.; Rasschaert, G.; Haesebrouck, F.; Temmerman, R.; Rutjens, S.; De Backer, P.; Devreese, M. The impact of therapeutic-dose induced intestinal enrofloxacin concentrations in healthy pigs on fecal Escherichia coli populations. BMC Vet. Res. 2020, 16, 382. [Google Scholar] [CrossRef]
- Serrano, M.J.; Mitjana, O.; Bonastre, C.; Laborda, A.; Falceto, M.F.; García-Gonzalo, D.; Abilleira, E.; Elorduy, J.; Bousquet-Melou, A.; Mata, L.; et al. Is Blood a Good Indicator for Detecting Antimicrobials in Meat? Evidence for the Development of In Vivo Surveillance Methods. Antibiotics 2020, 9, 175. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Ohlsson, A.K.; Ullberg, M.; Özenci, V. Evaluation of species-specific PCR, Bruker MS, VITEK MS and the VITEK 2 system for the identification of clinical Enterococcus isolates. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 3073–3077. [Google Scholar] [CrossRef]
- Bilecen, K.; Yaman, I.G.; Ciftci, U.; Laleli, Y.R. Performances and reliability of Bruker Microflex LT and VITEK MS MALDI-TOF mass spectrometry systems for the identification of clinical microorganisms. BioMed Res. Int. 2015, 2015, 516410. [Google Scholar] [CrossRef] [Green Version]
- Real Decreto 53/2013, de 1 de febrero, por el que se establecen las normas básicas aplicables para la protección de los animales utilizados en experimentación y otros fines científicos, incluyendo la docencia. Boletín Of. Estado 2013, 34, 11370–11421.
- Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes. Off. J. Eur. Union 2010, 276, 33–79.
- Hershberger, E.; Oprea, S.F.; Donabedian, S.M.; Perri, M.; Bozigar, P.; Bartlett, P.; Zervos, M. Epidemiology of antimicrobial resistance in enterococci of animal origin. J. Antimicrob. Chemother. 2005, 55, 127–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Liu, K.; Lai, J.; Wu, C.; Shen, J.; Wang, Y. Prevalence and antimicrobial resistance of Enterococcus species of food animal origin from Beijing and Shandong Province, China. J. Appl. Microbiol. 2012, 114, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S.; Igrejas, G.; Capelo-Martinez, J.L.; Poeta, P. Antibiotic resistance and mechanisms implicated in fecal enterococci recovered from pigs, cattle and sheep in a Portuguese slaughterhouse. Ann. Microbiol. 2012, 62, 1485–1494. [Google Scholar] [CrossRef]
- Nowakiewicz, A.; Ziółkowska, G.; Troscianczyk, A.; Zięba, P.; Gnat, S. Determination of antimicrobial resistance of Enterococcus strains isolated from pigs and their genotypic characterization by method of amplification of DNA fragments surrounding rare restriction sites (ADSRRS fingerprinting). J. Med. Microbiol. 2017, 66, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Aarestrup, F.M.; Agerso, Y.; Gerner-Smidt, P.; Madsen, M.; Jensen, L.B. Comparison of antimicrobial resistance phenotypes and resistance genes in Enterococcus faecalis and Enterococcus faecium from humans in the community, broilers, and pigs in Denmark. Diagn. Microbiol. Infect. Dis. 2000, 37, 127–137. [Google Scholar] [CrossRef]
- Wist, V.; Morach, M.; Schneeberger, M.; Cernela, N.; Stevens, M.J.A.; Zurfluh, K.; Stephan, R.; Nüesch-Inderbinen, M. Phenotypic and genotypic traits of vancomycin-resistant enterococci from healthy food-producing animals. Microorganisms 2020, 8, 261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruksakorn, C.; Pimarn, C.; Boonsoongnern, A.; Narongsak, W. Detection and phenotypic characterization of vancomycin-resistant enterococci in pigs in Thailand. Agric. Nat. Resour. 2016, 50, 199–203. [Google Scholar]
- Bager, F.; Madsen, M.; Christensen, J.; Aarestrup, F.M. Avoparcin used as a growth promoter is associated with the occurrence of vancomycin-resistant Enterococcus faecium on Danish poultry and pig farms. Prev. Vet. Med. 1997, 31, 95–112. [Google Scholar] [CrossRef]
- Directive 97/6/EC. of 30 January amending Council Directive 70/524/EEC concerning additives in feeding stuffs. Off. J. Eur. Commun. 1997, 35, 11–13.
- Yoshimura, H.; Ishimaru, M.; Endoh, Y.S.; Suginaka, M.; Yamatani, S. Isolation of glycopeptide-resistant enterococci from chicken in Japan. Antimicrob. Agents Chemother. 1998, 42, 3333. [Google Scholar] [CrossRef] [Green Version]
- Lauderdale, T.L.; Shiau, Y.R.; Wang, H.Y.; Lai, J.F.; Huang, I.W.; Chen, P.C.; Chen, H.Y.; Lai, S.S.; Liu, Y.F.; Ho, M. Effect of banning vancomycin analogue avoparcin on vancomycin-resistant enterococci in chicken farms in Taiwan. Environ. Microbiol. 2007, 9, 819–823. [Google Scholar] [CrossRef]
- Hammerum, A.M.; Lester, C.H.; Heuer, O.E. Antimicrobial resistant enterococci in animals and meat: A human health hazard? Foodborne Pathog. Dis. 2010, 7, 1137–1146. [Google Scholar] [CrossRef] [PubMed]
- Frye, J.G.; Jackson, C.R. Genetic mechanisms of antimicrobial resistance identified in Salmonella enterica, Escherichia coli, and Enteroccocus spp. isolated from U.S. food animals. Front. Microbiol. 2013, 4, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrido, A.M.; Galvez, A.; Pulido, R.P. Antimicrobial resistance in enterococci. J. Infect. Dis. Ther. 2014, 2, 4. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.O.; Baptiste, K.E. Vancomycin-resistant enterococci: A review of antimicrobial resistance mechanisms and perspectives of human and animal health. Microb. Drug Resist. 2018, 24, 590–606. [Google Scholar] [CrossRef] [Green Version]
- Riviere, J.E.; Spoo, J.W. Chapter 42. Tetracycline Antibiotics. In Veterinary Pharmacology and Therapeutics, 8th ed.; Adams, R.H., Ed.; Iowa State University Press: Ames, IA, USA, 2001; pp. 828–840. [Google Scholar]
- Papich, M.G.; Riviere, J.E. Chapter 45. Fluoroquinolone Antimicrobial Drugs. In Veterinary Pharmacology and Therapeutics, 8th ed.; Adams, R.H., Ed.; Iowa State University Press: Ames, IA, USA, 2001; pp. 898–917. [Google Scholar]
- Græsbøll, K.; Damborg, P.; Mellerup, A.; Herrero-Fresno, A.; Larsen, I.; Holm, A.; Nielsen, J.P.; Christiansen, L.E.; Angen, Ø.; Ahmed, S.; et al. Effect of tetracycline dose and treatment mode on selection of resistant coliform bacteria in nursery pigs. Appl. Environ. Microbiol. 2017, 83, 00538-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geser, N.; Stephan, R.; Hächler, H. Occurrence and characteristics of extended spectrum β-lactamase (ESBL) producing Enterobacteriaceae in food producing animals, minced meat and raw milk. BMC Vet. Res. 2012, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Loayza, F.; Graham, J.P.; Trueba, G. Factors obscuring the role of E. coli from Domestic animals in the global antimicrobial resistance crisis: An evidence-based review. Int. J. Environ. Res. Public Health 2020, 17, 3061. [Google Scholar] [CrossRef] [PubMed]
- Escudero, E.; Vinue, L.; Teshager, T.; Torres, C.; Moreno, M.A. Resistance mechanisms and farm-level distribution of fecal Escherichia coli isolates resistant to extended-spectrum cephalosporins in pigs in Spain. Res. Vet. Sci. 2010, 88, 83–87. [Google Scholar] [CrossRef]
- Ewers, C.; Bethe, A.; Semmler, T.; Guenther, S.; Wieler, L.H. Extended spectrum beta-lactamase-producing and AmpC-producing Escherichia coli from livestock and companion animals, and their putative impact on public health: A global perspective. Clin. Microbiol. Infect. 2012, 18, 646–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, J.; Hille, K.; Ruddat, I.; Mellmann, A.; Köck, R.; Kreienbrock, L. Simultaneous occurrence of MRSA and ESBL-producing Enterobacterales on pig farms and in nasal and stool samples from farmers. Vet. Microbiol. 2017, 200, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, I.; Garcia-Graells, C.; Biasino, W.; Gowda, T.; Botteldoorn, N.; De Zutter, L. High Abundance and diversity of extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli in faeces and tonsils of pigs at slaughter. Vet. Microbiol. 2017, 208, 190–194. [Google Scholar] [CrossRef] [PubMed]
- De Koster, S.; Ringenier, M.; Lammens, C.; Stegeman, A.; Tobias, T.; Velkers, F.; Vernooij, H.; Kluytmans-Van Den Bergh, M.; Kluytmans, J.; Dewulf, J.; et al. ESBL-producing, carbapenem and ciprofloxacin-resistant Escherichia coli in belgian and dutch broiler and pig farms: A cross-sectional and cross-border study. Antibiotics 2021, 10, 945. [Google Scholar] [CrossRef] [PubMed]
- Fournier, C.; Aires de Sousa, M.; Nordmann, P.; Poirel, L. Occurrence of CTX-M-15- and MCR-1-producing Enterobacterales in pigs in Portugal: Evidence of direct links with antibiotic selective pressure. Int. J. Antimicrob. Agents 2020, 55, 105802. [Google Scholar] [CrossRef]
- Picozzi, S.C.M.; Casellato, S.; Rossini, M.; Paola, G.; Tejada, M.; Costa, E.; Carmignani, L. Extended-spectrum beta-lactamase-positive Escherichia coli causing complicated upper urinary tract infection: Urologist should act in time. Urol. Ann. 2014, 6, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Liu, H.; Wang, L.; Peng, Q.; Li, Y.; Zhou, H.; Li, Q. Molecular characterization of extended-spectrum β-lactamase-producing multidrug resistant Escherichia coli from swine in Northwest China. Front. Microbiol. 2018, 9, 1756. [Google Scholar] [CrossRef] [Green Version]
- Benavides, J.A.; Salgado-Caxito, M.; Opazo-Capurro, A.; Muñoz, P.G.; Piñeiro, A.; Medina, M.O.; Rivas, L.; Munita, J.; Millán, J. ESBL-producing Escherichia coli carrying CTX-M genes circulating among livestock, dogs, and wild mammals in small-scale farms of central Chile. Antibiotics 2021, 10, 510. [Google Scholar] [CrossRef]
- Benavides, J.A.; Shiva, C.; Virhuez, M.; Tello, C.; Appelgren, A.; Vendrell, J.; Solassol, J.; Godreuil, S.; Streicker, D.G. Extended spectrum beta-lactamase-producing Escherichia coli in common vampire bats Desmodus rotundus and livestock in Peru. Zoonoses Public Health 2018, 65, 454–458. [Google Scholar] [CrossRef] [Green Version]
- Benavides, J.A.; Streicker, D.G.; Gonzales, M.S.; Rojas-Paniagua, E.; Shiva, C. Knowledge and use of antibiotics among low-income small-scale farmers of Peru. Prev. Vet. Med. 2021, 189, 105287. [Google Scholar] [CrossRef] [PubMed]
- Snow, L.; Warner, R.; Cheney, T.; Wearing, H.; Stokes, M.; Harris, K.; Teale, C.; Coldham, N. Risk factors associated with extended spectrum beta-lactamase Escherichia coli (CTX-M) on dairy farms in NorthWest England and NorthWales. Prev. Vet. Med. 2012, 106, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Hille, K.; Felski, M.; Ruddat, I.; Woydt, J.; Schmid, A.; Friese, A.; Fischer, J.; Sharp, H.; Valentin, L.; Michael, G.B.; et al. Association of farm-related factors with characteristics profiles of extended-spectrum β-lactamase-/plasmid-mediated AmpC -lactamase-producing Escherichia coli isolates from German livestock farms. Vet. Microbiol. 2018, 223, 93–99. [Google Scholar] [CrossRef]
- Gay, N.; LeClaire, A.; Laval, M.; Miltgen, G.; Jégo, M.; Stéphane, R.; Jaubert, J.; Belmonte, O.; Cardinale, E. Risk factors of extended-spectrum β-lactamase producing Enterobacteriaceae occurrence in farms in Reunion, Madagascar and Mayotte Islands, 2016–2017. Vet. Sci. 2018, 5, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roschanski, N.; Friese, A.; von Salviati-Claudius, C.; Hering, J.; Kaesbohrer, A.; Kreienbrock, L.; Roesler, U. Prevalence of carbapenemase producing Enterobacterales isolated from German pig-fattening farms during the years 2011–2013. Vet. Microbiol. 2017, 200, 124–129. [Google Scholar] [CrossRef]
- Mollenkopf, D.F.; Stull, J.W.; Mathys, D.A.; Bowman, A.S.; Feicht, S.M.; Grooters, S.V.; Daniels, J.B.; Wittuma, T.E. Carbapenemase-producing Enterobacterales recovered from the environment of a swine farrow-to-finish operation in the United States. Antimicrob. Agents Chemother. 2017, 61, e01298-16. [Google Scholar] [CrossRef] [Green Version]
- Canton, R.; Coque, T.M. The CTX-M b-lactamase pandemic. Curr. Opin. Microbiol. 2006, 9, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Naas, T.; Oxacelay, C.; Nordmann, P. Identification of CTXM-type extended-spectrum-b-lactamase genes using real-time PCR and pyrosequencing. Antimicrob. Agents Chemother. 2007, 51, 223–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, D.L. Resistance in Gram-negative bacteria: Enterobacteriaceae. Am. J. Med. 2006, 119, S20–S28. [Google Scholar] [CrossRef] [PubMed]
- Réglier-Poupet, H.; Naas, T.; Carrer, A.; Cady, A.; Adam, J.M.; Fortineau, N.; Poyart, C. Performance of ChromID ESBL, a chromogenic medium for detection of Enterobacteriaceae producing extended-spectrum b-lactamases. J. Med. Microbiol. 2008, 573, 310–315. [Google Scholar] [CrossRef] [Green Version]
- Burow, E.; Rostalski, A.; Harlizius, J.; Gangl, A.; Simoneit, C.; Grobbel, M.; Kollas, C.; Tenhagen, B.A.; Käsbohrer, A. Antibiotic resistance in Escherichia coli from pigs from birth to slaughter and its association with antibiotic treatment. Prev. Vet. Med. 2019, 165, 52–62. [Google Scholar] [CrossRef]
- Modi, S.R.; Collins, J.J.; Relman, D.A. Antibiotics and the gut microbiota. J. Clin. Investig. 2014, 124, 4212–4218. [Google Scholar] [CrossRef] [Green Version]
- Wei, R.; Ge, F.; Huang, S.; Chen, M.; Wang, R. Occurrence of veterinary antibiotics in animal wastewater and surface water around farms in Jiangsu Province, China. Chemosphere 2011, 82, 1408–1414. [Google Scholar] [CrossRef]
- Lay, K.K.; Koowattananukul, C.; Chansong, N.; Chuanchuen, R. Antimicrobial resistance, virulence, and phylogenetic characteristics of Escherichia coli isolates from clinically healthy swine. Foodborne Pathog. Dis. 2012, 9, 992–1001. [Google Scholar] [CrossRef]
- Kanwar, N.; Scott, H.M.; Norby, B.; Loneragan, G.H.; Vinasco, J.; Cottell, J.L.; Chalmers, G.; Chengappa, M.M.; Bai, J.; Boerlin, P. Impact of treatment strategies on cephalosporin and tetracycline resistance gene quantities in the bovine fecal metagenome. Sci. Rep. 2014, 4, 5100. [Google Scholar] [CrossRef] [Green Version]
- Jensen, L.B.; Birk, T.; Borck Høg, B.; Stehr, L.; Aabo, S.; Korsgaard, H. Cross and coresistance among Danish porcine E. coli isolates. Res. Vet. Sci. 2018, 119, 247–249. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.J.; Chen, C.X.; Yue, L.; Sun, Y.X.; Ding, H.Z.; Liu, Y.H. Excretion of enrofloxacin in pigs and its effect on ecological environment. Environ. Toxicol. Pharmacol. 2008, 26, 272–277. [Google Scholar] [CrossRef]
- Edlund, C.; Nord, C.E. Effect of quinolones on intestinal ecology. Drugs 1999, 58, 65–70. [Google Scholar] [CrossRef]
- Giguère, S.; Prescott, J.; Dowling, P. Antimicrobial Therapy in Veterinary Medicine, 5th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Alvarez, A.; Pérez, M.; Prieto, J.; Molina, A.; Real, R.; Merino, G. Fluoroquinolone efflux mediated by ABC transporters. J. Pharm. Sci. 2008, 97, 3483–3493. [Google Scholar] [CrossRef]
- Donskey, C.J.; Helfand, M.S.; Pultz, N.J.; Rice, L.B. Effect of parenteral fluoroquinolone administration on persistence of vancomycin-resistant Enterococcus faecium in the mouse gastrointestinal tract. Antimicrob. Agents Chemother. 2004, 48, 326–328. [Google Scholar] [CrossRef] [Green Version]
- Erwin, S.; Foster, D.M.; Jacob, M.E.; Papich, M.G.; Lanzas, C. The effect of enrofloxacin on enteric Escherichia coli: Fitting a mathematical model to in vivo data. PLoS ONE 2020, 15, e0228138. [Google Scholar] [CrossRef]
- Béraud, R.; Huneault, L.; Bernier, D.; Beaudry, F.; Letellier, A.; del Castillo, J.R.E. Comparison of the selection of antimicrobial resistance in fecal Escherichia coli during enrofloxacin administration with a local drug delivery system or with intramuscular injections in a swine model. Can. J. Vet. Res. 2008, 72, 311–319. [Google Scholar]
- Römer, A.; Scherz, G.; Reupke, S.; Meißner, J.; Wallmann, J.; Kietzmann, M.; Kaspar, H. Effects of intramuscularly administered enrofloxacin on the susceptibility of commensal intestinal Escherichia coli in pigs (Sus scrofa domestica). BMC Vet. Res. 2017, 13, 378. [Google Scholar] [CrossRef] [Green Version]
- Wiuff, C.; Lykkesfeldt, J.; Aarestrup, F.M.; Svendsen, O. Distribution of enrofloxacin in intestinal tissue and contents of healthy pigs after oral and intramuscular administrations. J. Vet. Pharmacol. Ther. 2002, 25, 335–342. [Google Scholar] [CrossRef]
- Anadon, A.; Martinez-Larranaga, M.R.; Diaz, M.J.; Fernandez-Cruz, M.L.; Martinez, M.A.; Frejo, M.T.; Martínez, M.; Iturbe, J.; Tafur, M. Pharmacokinetic variables and tissue residues of enrofloxacin and ciprofloxacin in healthy pigs. Am. J. Vet. Res. 1999, 60, 1377–1382. [Google Scholar]
- Ojer-Usoz, E.; Gonzalez, D.; Vitas, A.I.; Leiva, J.; Garcia-Jalon, I.; Febles-Casquero, A.; Escolano, M.S. Prevalence of extended-spectrum beta-lactamase-producing Enterobacteriaceae in meat products sold in Navarra, Spain. Meat Sci. 2013, 93, 316–321. [Google Scholar] [CrossRef]
- Leangapichart, T.; Lunha, K.; Jiwakanon, J.; Angkititrakul, S.; Järhult, J.D.; Magnusson, U.; Sunde, M. Characterization of Klebsiella pneumoniae complex isolates from pigs and humans in farms in Thailand: Population genomic structure, antibiotic resistance and virulence genes. J. Antimicrob. Chemother. 2021, 76, 2012–2016. [Google Scholar] [CrossRef]
- Pulss, S.; Semmler, T.; Prenger-Berninghoff, E.; Bauerfeind, R.; Ewers, C. First report of an Escherichia coli strain from swine carrying an OXA- 181 carbapenemase and the colistin resistance determinant MCR-1. Int. J. Antimicrob. Agents 2017, 50, 232–236. [Google Scholar] [CrossRef]
- Pruthvishree, B.S.; Vinodh Kumar, O.R.; Sinha, D.K.; Malik, Y.P.S.; Dubal, Z.B.; Desingu, P.A.; Shivakumar, M.; Krishnaswamy, N.; Singh, B.R. Spatial molecular epidemiology of carbapenem-resistant and New Delhi metallo beta-lactamase (blaNDM) producing Escherichia coli in the piglets of organized farms in India. J. Appl. Microbiol. 2017, 122, 1537–1546. [Google Scholar] [CrossRef]
- Fischer, J.; Rodríguez, I.; Schmoger, S.; Friese, A.; Roesler, U.; Helmuth, R.; Guerra, B. Escherichia coli producing VIM-1 carbapenemase isolated on a pig farm. J. Antimicrob. Chemother. 2012, 67, 1793–1795. [Google Scholar] [CrossRef]
- Collignon, P.C.; Conly, J.M.; Andremont, A.; McEwen, S.A.; Aidara-Kane, A. world health organization ranking of antimicrobials according to their importance in human medicine: A critical step for developing risk management strategies to control antimicrobial resistance from food animal production. Clin. Infect. Dis. 2016, 63, 1087–1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samra, Z.; Bahar, J.; Madar-Shapiro, L.; Aziz, N.; Israel, S.; Bishara, J. Evaluation of CHROMagar KPC for rapid detection of carbapenem-resistant Enterobacteriaceae. J. Clin. Microbiol. 2008, 46, 3110–3111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdallah, H.M.; Reuland, E.A.; Wintermans, B.B.; Al Naiemi, N.; Koek, A.; Abdelwahab, A.M.; Ammar, A.M.; Mohamed, A.A.; Vandenbroucke-Grauls, C.M.J.E. Extended-spectrum beta lactamases and/or carbapenemases-producing Enterobacteriaceae isolated from retail chicken meat in Zagazig, Egypt. PLoS ONE 2015, 10, e0136052. [Google Scholar]
- Hamza, E.; Dorgham, S.M.; Hamza, D.A. Carbapenemase-producing Klebsiella pneumoniae in broiler poultry farming in Egypt. J. Glob. Antimicrob. Resist. 2016, 7, 8–10. [Google Scholar] [CrossRef]
- Masters, N.; Christie, M.; Stratton, H.; Katouli, M. Viability and stability of Escherichia coli and enterococci populations in fecal samples upon freezing. Can. J. Microbiol. 2015, 61, 495–501. [Google Scholar] [CrossRef]
- Saliba, R.; Zahar, J.R.; El Allaoui, F.; Carbonnelle, E.; Lescat, M. Impact of freeze/thaw cycles and single freezing at −80 °C on the viability of aerobic bacteria from rectal swabs performed with the Eswab™ system. Diagn. Microbiol. Infect. Dis. 2020, 96, 114895. [Google Scholar] [CrossRef] [PubMed]
- Tedjo, D.I.; Jonkers, D.M.A.E.; Savelkoul, P.H.; Masclee, A.A.; van Best, N.; Pierik, M.J.; Penders, J. The effect of sampling and storage on the fecal microbiota composition in healthy and diseased subjects. PLoS ONE 2015, 10, e0126685. [Google Scholar] [CrossRef]
- Deschamps, C.; Fournier, E.; Uriot, O.; Lajoie, F.; Verdierm, C.; Comtet-Marre, S.; Thomas, M.; Kapel, N.; Cherbuy, C.; Alric, M.; et al. Comparative methods for fecal sample storage to preserve gut microbial structure and function in an in vitro model of the human colon. Appl. Microbiol. Biotechnol. 2020, 104, 10233–10247. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Day | Sample Code | Number of Animals |
|---|---|---|---|
| Control | 0 | C0 | 4 |
| 14 | C14 | 4 | |
| Oxytetracycline | 0 | T0 | 10 |
| 6 | T6 | 6 | |
| 13 | T13 | 5 | |
| 15 | T15 | 6 | |
| 17 | T17 | 6 | |
| 19 | T19 | 6 | |
| Enrofloxacin | 0 | E0 | 12 |
| 3 | E3 | 6 | |
| 4 | E4 | 6 | |
| 5 | E5 | 6 | |
| 6 | E6 | 6 | |
| 7 | E7 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Fandos, E.; Martínez-Laorden, A.; Abad-Fau, A.; Sevilla, E.; Bolea, R.; Serrano, M.J.; Mitjana, O.; Bonastre, C.; Laborda, A.; Falceto, M.V.; et al. Effect of Intramuscularly Administered Oxytetracycline or Enrofloxacin on Vancomycin-Resistant Enterococci, Extended Spectrum Beta-Lactamase- and Carbapenemase-Producing Enterobacteriaceae in Pigs. Animals 2022, 12, 622. https://doi.org/10.3390/ani12050622
González-Fandos E, Martínez-Laorden A, Abad-Fau A, Sevilla E, Bolea R, Serrano MJ, Mitjana O, Bonastre C, Laborda A, Falceto MV, et al. Effect of Intramuscularly Administered Oxytetracycline or Enrofloxacin on Vancomycin-Resistant Enterococci, Extended Spectrum Beta-Lactamase- and Carbapenemase-Producing Enterobacteriaceae in Pigs. Animals. 2022; 12(5):622. https://doi.org/10.3390/ani12050622
Chicago/Turabian StyleGonzález-Fandos, Elena, Alba Martínez-Laorden, Ana Abad-Fau, Eloisa Sevilla, Rosa Bolea, María Jesús Serrano, Olga Mitjana, Cristina Bonastre, Alicia Laborda, María Victoria Falceto, and et al. 2022. "Effect of Intramuscularly Administered Oxytetracycline or Enrofloxacin on Vancomycin-Resistant Enterococci, Extended Spectrum Beta-Lactamase- and Carbapenemase-Producing Enterobacteriaceae in Pigs" Animals 12, no. 5: 622. https://doi.org/10.3390/ani12050622
APA StyleGonzález-Fandos, E., Martínez-Laorden, A., Abad-Fau, A., Sevilla, E., Bolea, R., Serrano, M. J., Mitjana, O., Bonastre, C., Laborda, A., Falceto, M. V., & Pagán, R. (2022). Effect of Intramuscularly Administered Oxytetracycline or Enrofloxacin on Vancomycin-Resistant Enterococci, Extended Spectrum Beta-Lactamase- and Carbapenemase-Producing Enterobacteriaceae in Pigs. Animals, 12(5), 622. https://doi.org/10.3390/ani12050622