
Mitochondrial DNA and Microsatellite Analyses Showed Panmixia between Temporal Samples in Endangered Anguilla japonica in the Pearl River Basin (China)

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Extraction
2.2. Mitochondrial DNA Sequencing
2.3. Microsatellite Genotyping
2.4. Data Analysis for mtDNA
2.5. Data Analysis for nSSR
3. Results
3.1. Genetic Variation and Differentiation Based on Mitochondrial DNA
3.2. Microsatellite Marker Variation and Genetic Diversity
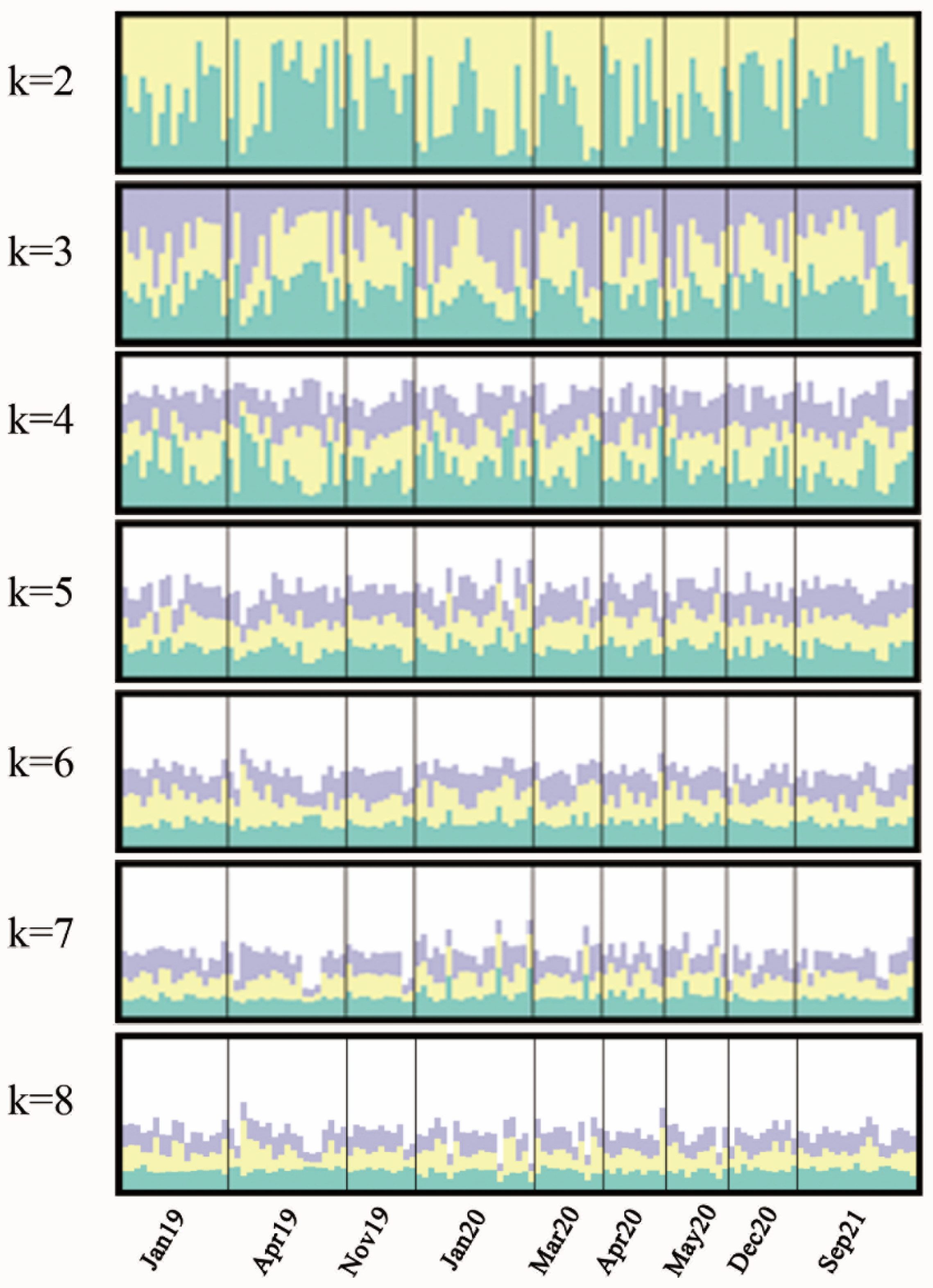
3.3. Genetic Differentiation and Structure Based on Microsatellite Markers
3.4. Demographic History
4. Discussion
4.1. Genetic Diversity
4.2. Evidence of Panmixia
4.3. Demographic Expansion
4.4. Implications for Conservation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Katsumi, T. Oceanic biology: Spawning of eels near a seamount. Nature 2006, 439, 929. [Google Scholar] [CrossRef]
- Han, Y.S.; Zhang, H.; Tseng, Y.H.; Shen, M.L. Larval Japanese eel (Anguilla japonica) as sub-surface current bio-tracers on the East Asia continental shelf. Fish. Oceanogr. 2012, 21, 281–290. [Google Scholar] [CrossRef]
- Hsiung, K.-M.; Kimura, S.; Han, Y.S.; Takeshige, A.; Iizuka, Y. Effect of ENSO events on larval and juvenile duration and transport of Japanese eel (Anguilla japonica). PLoS ONE 2018, 13, e0195544. [Google Scholar] [CrossRef] [PubMed]
- Arai, T. Do we protect freshwater eels or do we drive them to extinction? SpringerPlus 2014, 3, 534. [Google Scholar] [CrossRef] [PubMed]
- Dekker, W. The history of commercial fisheries for European eel commenced only a century ago. Fish. Manag. Ecol. 2019, 26, 6–19. [Google Scholar] [CrossRef]
- Faulks, L.; Kaushik, P.; Taniguchi, S.; Sekino, M.; Nakamichi, R.; Yamamoto, Y.; Fujimori, H.; Okamoto, C.; Kodama, S.; Daryani, A. Inferring the demographic history of Japanese eel (Anguilla japonica) from genomic data: Insights for conservation and fisheries management. Aquat. Conserv. 2022, 32, 1092–1098. [Google Scholar] [CrossRef]
- Yu, L.; Liu, Y.; Liu, J. Gene-associated microsatellite markers confirm panmixia and indicate a different pattern of spatially varying selection in the endangered Japanese eel Anguilla japonica. J. Oceanol. Limnol. 2020, 38, 1572–1583. [Google Scholar] [CrossRef]
- Chen, J.Z.; Huang, S.L.; Han, Y.S. Impact of long-term habitat loss on the Japanese eel Anguilla japonica. Estuar. Coast. Shelf Sci. 2014, 151, 361–369. [Google Scholar] [CrossRef]
- Tseng, M.; Tzeng, W.; Lee, S. Genetic differentiation of the Japanese eel. In American Fisheries Society Symposium; American Fisheries Society: Bethesda, MD, USA, 2009; pp. 59–69. [Google Scholar]
- Gong, X.; Davenport, E.R.; Wang, D.; Clark, A.G. Lack of spatial and temporal genetic structure of Japanese eel (Anguilla japonica) populations. Conserv. Genet. 2019, 20, 467–475. [Google Scholar] [CrossRef]
- Tseng, M.C.; Kao, H.W.; Hung, Y.H.; Lee, T.L. A study of genetic variations, population size, and population dynamics of the catadromous Japanese eel Anguilla japonica (Pisces) in northern Taiwan. Hydrobiologia 2012, 683, 203–216. [Google Scholar] [CrossRef][Green Version]
- Chan, I.; Chan, D.; Lee, S.; Tsukamoto, K. Genetic variability of the Japanese eel Anguilla japonica (Temminck & Schlegel) related to latitude. Ecol. Freshw. Fish. 1997, 6, 45–49. [Google Scholar] [CrossRef]
- Tseng, M.C.; Tzeng, W.N.; Lee, S.C. Population genetic structure of the Japanese eel Anguilla japonica in the northwest Pacific Ocean: Evidence of non-panmictic populations. Mar. Ecol. Prog Ser. 2006, 308, 221–230. [Google Scholar] [CrossRef]
- Shuai, F.M.; Li, X.H.; Yang, W.L.; Chen, W.T.; Lek, S. Habitat use of the Japanese eel (Anguilla japonica) and marbled eel (Anguilla marmorata) in the large subtropical Pearl River. In Annales de Limnologie-International Journal of Limnology; EDP Sciences: Les Ulis, France, 2021; Volume 57, p. 8. [Google Scholar] [CrossRef]
- Shuai, F.M.; Li, X.H.; Zhu, S.L.; Wu, Z.; Zhang, Y.Q.; Chen, W.T.; Liu, Y.Q. The spatial distribution of anguilla japonica and anguilla marmorata in the Pearl River. Acta Hydrobiol. Sin. 2019, 43, 7. [Google Scholar] [CrossRef]
- Han, Y.S.; Hung, C.L.; Liao, Y.F.; Tzeng, W.N. Population genetic structure of the Japanese eel Anguilla japonica: Panmixia at spatial and temporal scales. Mar. Ecol. Prog. Ser. 2010, 401, 221–232. [Google Scholar] [CrossRef]
- Swindell, S.R.; Plasterer, T.N. Seqman. Sequence Data Analysis Guidebook; Springer: Totowa, NJ, USA, 1997; pp. 75–89. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Fu, Y.X. Statistical Tests of Neutrality of Mutations Against Population Growth, Hitchhiking and Background Selection. Genetics. 1997, 147, 915–925. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef]
- Donaldson, K.A.; Wilson, R.R., Jr. Amphi-panamic geminates of snook (Percoidei: Centropomidae) provide a calibration of the divergence rate in the mitochondrial DNA control region of fishes. Mol. Phylogenet. Evol. 1999, 13, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Hulce, D.; Li, X.; Snyder-Leiby, T.; Liu, C.J. GeneMarker® genotyping software: Tools to increase the statistical power of DNA fragment analysis. J. Biomol. Tech. JBT 2011, 22, S35. [Google Scholar]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.; Shipley, P. MICRO-CHECKER: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Benjamini, Y.; Yekutieli, D. The control of the false discovery rate in multiple testing under dependency. Ann. Stat. 2001, 29, 1165–1188. [Google Scholar] [CrossRef]
- Hubisz, M.J.; Falush, D.; Stephens, M.; Pritchard, J.K. Inferring weak population structure with the assistance of sample group information. Mol. Ecol. Resour. 2009, 9, 1322–1332. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg, N.A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef]
- Rosenberg, N.A. DISTRUCT: A program for the graphical display of population structure. Mol. Ecol. Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Sharma, R.; Ahlawat, S.; Sharma, H.; Sharma, P.; Panchal, P.; Arora, R.; Tantia, M. Microsatellite and mitochondrial DNA analyses unveil the genetic structure of native sheep breeds from three major agro-ecological regions of India. Sci. Rep. 2020, 10, 20422. [Google Scholar] [CrossRef]
- Earl, D.A.; VonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Marini, M.; Pedrosa Gerasmio, I.R.; Santos, M.D.; Shibuno, T.; Daryani, A.; Romana Eguia, M.R.R.; Wibowo, A. Genetic diversity, population structure and demographic history of the tropical eel Anguilla bicolor pacifica in Southeast Asia using mitochondrial DNA control region sequences. Glob. Ecol. Conserv. 2021, 26, e01493. [Google Scholar] [CrossRef]
- Butkauskas, D.; Ragauskas, A.; Sruoga, A.; Ložys, L.; Tzeng, W.N. Current knowledge about European eel Anguilla anguilla (L.) mtDNA D-loop region haplotypic variety. Acta Zool. Litu. 2009, 19, 253–267. [Google Scholar] [CrossRef]
- Gong, X.; Ren, S.; Cui, Z.; Yue, L. Genetic evidence for panmixia of Japanese eel (Anguilla japonica) populations in China. Genet. Mol. Res. 2014, 13, 768–781. [Google Scholar] [CrossRef] [PubMed]
- Cheng, P.; Tzeng, W. Timing of metamorphosis and estuarine arrival across the dispersal range of the Japanese eel Anguilla japonica. Mar. Ecol. Prog. Ser. 1996, 131, 87–96. [Google Scholar] [CrossRef]
- Ishikawa, S.; Aoyama, J.; Tsukamoto, K.; Nishida, M. Population structure of the Japanese eel Anguilla japonica as examined by mitochondrial DNA sequencing. Fish. Sci. 2001, 67, 246–253. [Google Scholar] [CrossRef]
- Han, Y.S.; Sun, Y.L.; Liao, Y.F.; Liao, I.; Shen, K.N.; Tzeng, W.N. Temporal analysis of population genetic composition in the overexploited Japanese eel Anguilla japonica. Mar. Biol. 2008, 155, 613–621. [Google Scholar] [CrossRef]
- Han, Y.S.; Iizuka, Y.; Tzeng, W.N. Does variable habitat usage by the Japanese eel lead to population genetic differentiation? Zool. Stud. 2010, 49, 392–397. [Google Scholar]
- Roberts, D.G.; Ayre, D.J. Panmictic population structure in the migratory marine sparid Acanthopagrus australis despite its close association with estuaries. Mar. Ecol. Prog. Ser. 2010, 412, 223–230. [Google Scholar] [CrossRef]
- Khatun, M.; Mondol, M.R.K.; Rahman, S.; Alam, M.A.; Bashar, M.A.; Mahmud, Y. Existence of panmictic Hilsa shad (Tenualosa ilisha) populations from two large river ecosystems in Bangladesh revealed by mitochondrial control region. Bangl. J. Fish. 2022, 34, 77–86. [Google Scholar] [CrossRef]
- Campbell, M.A.; Joslin, S.E.; Goodbla, A.M.; Willmes, M.; Hobbs, J.A.; Lewis, L.S.; Finger, A.J. Polygenic Discrimination of Migratory Phenotypes in an Estuarine Forage Fish. G3-Genes Genom. Genet. 2022, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gascoyne, M.; Benjamin, G.; Schwarcz, H.P.; Ford, D.C. Sea-level lowering during the illinoian glaciation: Evidence from a Bahama “Blue Hole”. Science 1979, 205, 806–808. [Google Scholar] [CrossRef] [PubMed]
- Palm, S.; Dannewitz, J.; Prestegaard, T.; Wickström, H. Panmixia in European eel revisited: No genetic difference between maturing adults from southern and northern Europe. Heredity 2009, 103, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Gubili, C.; Schabetsberger, R.; Poellabauer, C.; Bates, B.; Wagstaff, R.M.; Woodward, L.M.; Sichrowsky, U.; Scheck, A.; Boseto, D.T.; Feunteun, E. High genetic diversity and lack of pronounced population structure in five species of sympatric Pacific eels. Fish. Manag. Ecol. 2019, 26, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Han, Y. Eel conservation in Taiwan. In Proceedings of the 16th East Asia Eel Resource Consortium Meeting, University of Tokyo, Tokyo, Japan, 1 December 2013. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Temporal Sample | N | H | Hd | π |
|---|---|---|---|---|
| Jan19 | 17 | 17 | 1 ± 0.020 | 0.005 ± 0.0004 |
| Apr19 | 19 | 19 | 1 ± 0.017 | 0.005 ± 0.0005 |
| Nov19 | 11 | 11 | 1 ± 0.039 | 0.005 ± 0.0004 |
| Jan20 | 19 | 18 | 0.994 ± 0.019 | 0.006 ± 0.0005 |
| Mar20 | 11 | 11 | 1 ± 0.039 | 0.006 ± 0.0007 |
| Apr20 | 10 | 10 | 1 ± 0.045 | 0.005 ± 0.0004 |
| May20 | 10 | 9 | 1 ± 0.052 | 0.006 ± 0.0007 |
| Dec20 | 11 | 11 | 1 ± 0.039 | 0.005 ± 0.0005 |
| Sep21 | 19 | 18 | 1 ± 0.019 | 0.005 ± 0.0006 |
| Overall | 127 | 124 | 0.997 ± 0.001 | 0.005 ± 0.0002 |
| Temporal Sample | Jan19 | Apr19 | Nov19 | Jan20 | Mar20 | Apr20 | May20 | Dec20 | Sep21 |
|---|---|---|---|---|---|---|---|---|---|
| Jan19 | 0 | 0.010 | 0.005 | 0.018 | 0.017 | 0.011 | 0.008 | 0.011 | 0.009 |
| Apr19 | 0.013 | 0 | 0.013 | 0.025 | 0.021 | 0.022 | 0.017 | 0.014 | 0.009 |
| Nov19 | −0.019 | −0.013 | 0 | 0.014 | 0.028 | −0.001 | −0.001 | 0.0004 | 0.003 |
| Jan20 | −0.016 | 0.005 | −0.014 | 0 | 0.023 | 0.015 | 0.0002 | 0.022 | 0.022 |
| Mar20 | −0.001 | 0.003 | −0.021 | 0.003 | 0 | 0.018 | 0.029 | 0.016 | 0.0002 |
| Apr20 | −0.011 | −0.007 | −0.018 | −0.0101 | −0.009 | 0 | 0.010 | 0.023 | 0.019 |
| May20 | 0.004 | −0.018 | −0.029 | −0.005 | 0.001 | −0.010 | 0 | 0.014 | 0.010 |
| Dec20 | −0.021 | −0.0004 | −0.040 | −0.028 | −0.017 | −0.026 | −0.032 | 0 | 0.007 |
| Sep21 | −0.015 | −0.014 | −0.017 | −0.013 | −0.002 | −0.023 | −0.009 | −0.022 | 0 |
| Source of Variation | d.f. | Sum of Squares | Variance Components | Percentage of Variation | Fixation Index |
|---|---|---|---|---|---|
| Among temporal samples | 8 | 34.49 | −0.06 Va | 1.12 | 0.011 |
| Within temporal samples | 116 | 589.75 | 5.08 Vb | 98.88 | |
| Total | 124 | 624.24 | 5.03 |
| Locus | Total Allele | A | Ae | I | Ho | He | uHe | F | Ar | Fis | Fit | FST |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AM062762 | 17 | 9 | 5.97 | 1.95 | 0.75 | 0.83 | 0.86 | 0.09 | 8.21 | 0.09 | 0.13 | 0.05 |
| AJ297601 | 4 | 3 | 1.75 | 0.77 | 0.49 | 0.42 | 0.43 | −0.17 | 3.04 | −0.17 | −0.11 | 0.05 |
| AJ297602 | 16 | 10 | 7.03 | 2.08 | 0.69 | 0.85 | 0.89 | 0.20 | 8.72 | 0.20 | 0.23 | 0.05 |
| AB051094 | 13 | 9 | 5.40 | 1.88 | 0.80 | 0.81 | 0.84 | 0.01 | 7.78 | 0.01 | 0.05 | 0.04 |
| AM062761 | 29 | 15 | 11.14 | 2.52 | 0.70 | 0.91 | 0.94 | 0.23 | 10.70 | 0.23 | 0.27 | 0.05 |
| AJ297603 | 19 | 9 | 5.97 | 1.95 | 0.72 | 0.82 | 0.86 | 0.13 | 7.94 | 0.13 | 0.18 | 0.06 |
| AJ297600 | 31 | 12 | 8.25 | 2.27 | 0.70 | 0.88 | 0.91 | 0.20 | 10.48 | 0.20 | 0.24 | 0.05 |
| AB051084 | 21 | 13 | 10.02 | 2.41 | 0.85 | 0.90 | 0.93 | 0.06 | 11.17 | 0.06 | 0.10 | 0.04 |
| Mean | 18.75 | 10.01 | 6.94 | 1.98 | 0.70 | 0.80 | 0.83 | 0.09 | 8.50 | 0.09 | 0.13 | 0.05 |
| SE | 2.88 | 1.12 | 0.98 | 0.18 | 0.04 | 0.05 | 0.06 | 0.04 | 0.86 | 0.04 | 0.04 | 0.00 |
| Temporal Sample | Total Alleles | A | Ae | I | Ho | He | uHe | F | Ar |
|---|---|---|---|---|---|---|---|---|---|
| Jan19 | 85 | 10.63 | 7.35 | 2.04 | 0.72 | 0.81 | 0.84 | 0.09 | 8.78 |
| Apr19 | 84 | 10.50 | 6.67 | 1.98 | 0.69 | 0.80 | 0.82 | 0.11 | 8.27 |
| Jun19 | 72 | 9.00 | 6.37 | 1.90 | 0.74 | 0.80 | 0.83 | 0.05 | 8.64 |
| Nov19 | 89 | 11.13 | 7.08 | 2.02 | 0.69 | 0.79 | 0.82 | 0.11 | 8.58 |
| Jan20 | 69 | 8.63 | 6.32 | 1.87 | 0.63 | 0.77 | 0.81 | 0.16 | 8.34 |
| Mar20 | 62 | 7.75 | 5.27 | 1.77 | 0.71 | 0.78 | 0.82 | 0.06 | 7.75 |
| Apr20 | 72 | 9.00 | 7.09 | 1.98 | 0.75 | 0.81 | 0.86 | 0.06 | 8.41 |
| May20 | 69 | 8.75 | 5.98 | 1.87 | 0.72 | 0.79 | 0.83 | 0.10 | 8.67 |
| Apr21 | 86 | 10.75 | 7.49 | 2.07 | 0.69 | 0.82 | 0.85 | 0.15 | 9.09 |
| Mean | 76.44 | 9.57 | 6.62 | 1.94 | 0.70 | 0.80 | 0.83 | 0.10 | 8.50 |
| SE | 3.20 | 0.40 | 0.24 | 0.03 | 0.01 | 0.01 | 0.01 | 0.01 | 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, Z.; Zhu, H.; Fan, J.; Ma, D. Mitochondrial DNA and Microsatellite Analyses Showed Panmixia between Temporal Samples in Endangered Anguilla japonica in the Pearl River Basin (China). Animals 2022, 12, 3380. https://doi.org/10.3390/ani12233380
Zhong Z, Zhu H, Fan J, Ma D. Mitochondrial DNA and Microsatellite Analyses Showed Panmixia between Temporal Samples in Endangered Anguilla japonica in the Pearl River Basin (China). Animals. 2022; 12(23):3380. https://doi.org/10.3390/ani12233380
Chicago/Turabian StyleZhong, Zaixuan, Huaping Zhu, Jiajia Fan, and Dongmei Ma. 2022. "Mitochondrial DNA and Microsatellite Analyses Showed Panmixia between Temporal Samples in Endangered Anguilla japonica in the Pearl River Basin (China)" Animals 12, no. 23: 3380. https://doi.org/10.3390/ani12233380
APA StyleZhong, Z., Zhu, H., Fan, J., & Ma, D. (2022). Mitochondrial DNA and Microsatellite Analyses Showed Panmixia between Temporal Samples in Endangered Anguilla japonica in the Pearl River Basin (China). Animals, 12(23), 3380. https://doi.org/10.3390/ani12233380