
Creating a Design Framework to Diagnose and Enhance Grassland Health under Pastoral Livestock Production Systems

 , , and
, , and 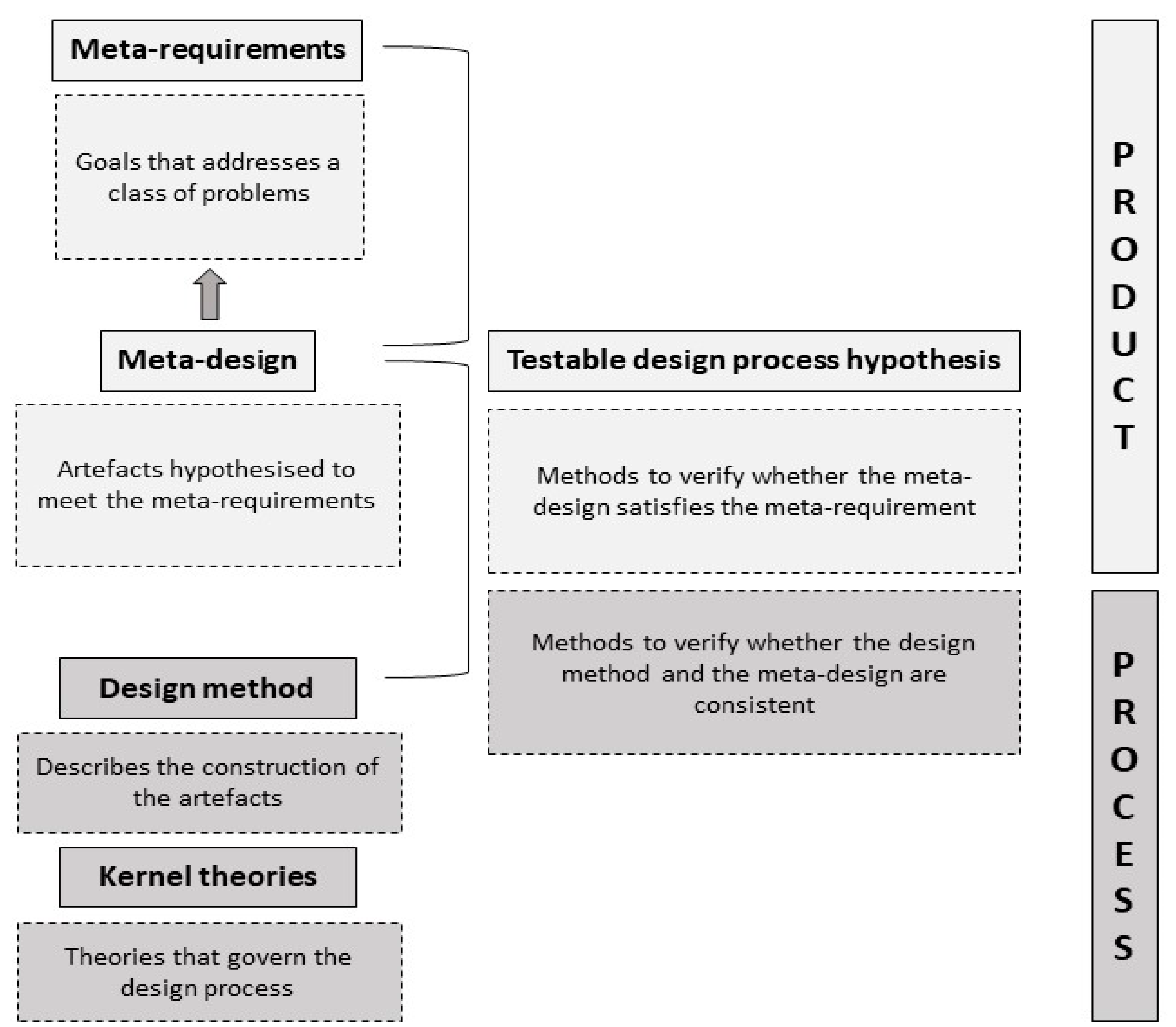{kind=link}
{kind=link}
{kind=link}
Abstract
Simple Summary
Abstract
1. Introduction
Systems Thinking Theory
2. Systems Components
2.1. Soil
2.2. Plants
2.3. Ruminants
3. Design Method
4. Testable Design Process Hypothesis
5. Design Framework and Further Applications
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wyborn, C.; Kalas, N.; Rust, N. Seeds of change: Provocations for a new research agenda. In Proceedings of the Biodiversity Revisited Symposium Conference Proceedings, Vienna, Austria, 11–13 September 2019. [Google Scholar] [CrossRef]
- Lausch, A.; Bastian, O.; Klotz, S.; Leitão, P.J.; Jung, A.; Rocchini, D.; Schaepman, M.E.; Skidmore, A.K.; Tischendorf, L.; Knapp, S. Understanding and assessing vegetation health by in situ species and remote-sensing approaches. Methods Ecol. Evol. 2018, 9, 1799–1809. [Google Scholar] [CrossRef]
- Teague, R.; Kreuter, U. Managing grazing to restore soil health, ecosystem function, and ecosystem services. Front. Sustain. Food Syst. 2020, 4, 534187. [Google Scholar] [CrossRef]
- Xu, D.; Guo, X. Some insights on grassland health assessment based on remote sensing. Sensors 2015, 15, 3070–3089. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, Z.; Wu, J. Grassland ecosystem services: A systematic review of research advances and future directions. Landsc. Ecol. 2020, 35, 793–814. [Google Scholar] [CrossRef]
- Sala, O.E.; Yahdjian, L.; Havstad, K.; Aguiar, M.R. Rangeland ecosystem services: Nature’s supply and humans’ demand. In Rangeland Systems; Briske, D.D., Ed.; Springer Nature: Cham, Switzerland, 2017; pp. 467–489. [Google Scholar] [CrossRef]
- White, R.P.; Murray, S.; Rohweder, M.; Prince, S.D.; Thompson, K.M. Grassland Ecosystems; World Resources Institute: Washington, DC, USA, 2000. [Google Scholar]
- Pereira, F.C.; Smith, C.M.; Maxwell, T.M.; Charters, S.M.; Logan, C.M.; Donovan, M.; Jayathunga, S.; Gregorini, P. Applying Spatial Analysis to Create Modern Rich Pictures for Grassland Health Analysis. Sustainability 2021, 13, 11535. [Google Scholar] [CrossRef]
- Ramankutty, N.; Foley, J.A. Estimating historical changes in global land cover: Croplands from 1700 to 1992. Glob. Biogeochem. Cycles 1999, 13, 997–1027. [Google Scholar] [CrossRef]
- Ramankutty, N.; Mehrabi, Z.; Waha, K.; Kremen, C.; Herrero, M.; Rieseberg, L.H. Trends in global agricultural land use: Implications for environmental health and food security. Annu. Rev. Plant Biol. 2018, 69, 789–815. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.J.; Steinfeld, H.; Henderson, B.; Mottet, A.; Opio, C.; Dijkman, J.; Falcucci, A.; Tempio, G. Tackling Climate Change through Livestock—A Global Assessment of Emissions and Mitigation Opportunities; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2013; ISBN 9789251079201. [Google Scholar]
- UNCCD. United Nations Convention to Combat Desertification. Available online: http://www.unccd.int/message-land-and-soil (accessed on 19 January 2022).
- Newbold, T.; Hudson, L.N.; Hill, S.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Kline, O.; Joshi, N.K. Mitigating the effects of habitat loss on solitary bees in agricultural ecosystems. Agriculture 2020, 10, 115. [Google Scholar] [CrossRef]
- Doggart, N.; Morgan-Brown, T.; Lyimo, E.; Mbilinyi, B.; Meshack, C.K.; Sallu, S.M.; Spracklen, D.V. Agriculture is the main driver of deforestation in Tanzania. Environ. Res. Lett. 2020, 15, 034028. [Google Scholar] [CrossRef]
- Lawrence, D.; Vandecar, K. Effects of tropical deforestation on climate and agriculture. Nat. Clim. Chang. 2015, 5, 27–36. [Google Scholar] [CrossRef]
- De Vries, M.; de Boer, I.J. Comparing environmental impacts for livestock products: A review of life cycle assessments. Livest. Sci. 2010, 128, 1–11. [Google Scholar] [CrossRef]
- Michalk, D.L.; Kemp, D.R.; Badgery, W.B.; Wu, J.; Zhang, Y.; Thomassin, P.J. Sustainability and future food security—A global perspective for livestock production. Land Degrad. Dev. 2019, 30, 561–573. [Google Scholar] [CrossRef]
- Díaz-Pereira, E.; Romero-Díaz, A.; de Vente, J. Sustainable grazing land management to protect ecosystem services. Mitig. Adapt. Strateg. Glob. Change 2020, 25, 1461–1479. [Google Scholar] [CrossRef]
- Duru, M.; Therond, O.; Martin, G.; Martin-Clouaire, R.; Magne, M.A.; Justes, E.; Journet, E.-P.; Aubertot, J.-N.; Savary, S.; Bergez, J.-E.; et al. How to implement biodiversity-based agriculture to enhance ecosystem services: A review. Agron. Sustain. Dev. 2015, 35, 1259–1281. [Google Scholar] [CrossRef]
- Martin, G.; Martin-Clouaire, R.; Duru, M. Farming system design to feed the changing world. A review. Agron. Sustain. Dev. 2013, 33, 131–149. [Google Scholar] [CrossRef]
- Simon, H.A. The science of design: Creating the artificial. Des. Issues 1988, 4, 67–82. [Google Scholar] [CrossRef]
- Lacombe, C.; Couix, N.; Hazard, L. Designing agroecological farming systems with farmers: A review. Agric. Syst. 2018, 165, 208–220. [Google Scholar] [CrossRef]
- Hatchuel, A.; Reich, Y.; Le Masson, P.; Weil, B.; Kazakçi, A. Beyond models and decisions: Situating design through generative functions. In Proceedings of the DS 75-2: 19th International Conference on Engineering Design (ICED13), Design for Harmonies, Vol. 2: Design Theory and Research Methodology, Seoul, Republic of Korea, 19–22 August 2013; pp. 233–242. [Google Scholar]
- Rizzo, D.; Marraccini, E.; Lardon, S.; Rapey, H.; Debolini, M.; Benoit, M.; Thenail, C. Farming systems designing landscapes: Land management units at the interface between agronomy and geography. Geogr. Tidsskr. 2013, 113, 71–86. [Google Scholar] [CrossRef]
- Dominati, E.J.; Maseyk, F.J.; Mackay, A.D.; Rendel, J.M. Farming in a changing environment: Increasing biodiversity on farm for the supply of multiple ecosystem services. Sci. Total Environ. 2019, 662, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Walls, J.G.; Widmeyer, G.R.; El Sawy, O.A. Building an information system design theory for vigilant EIS. Inf. Syst. Res. 1992, 3, 36–59. [Google Scholar] [CrossRef]
- Garrett, K.; Beck, M.R.; Marshall, C.J.; Fleming, A.E.; Logan, C.M.; Maxwell, T.M.; Greer, A.W.; Gregorini, P. Functional diversity vs. monotony: The effect of a multiforage diet as opposed to a single forage diet on animal intake, performance, welfare, and urinary nitrogen excretion. J. Anim. Sci. 2021, 99, skab058. [Google Scholar] [CrossRef]
- Distel, R.A.; Arroquy, J.I.; Lagrange, S.; Villalba, J.J. Designing diverse agricultural pastures for improving ruminant production systems. Front. Sustain. Food Syst. 2020, 4, 596869. [Google Scholar] [CrossRef]
- Beck, M.R.; Gregorini, P. Animal design through functional dietary diversity for future productive landscapes. Front. Sustain. Food Syst. 2021, 5, 546581. [Google Scholar] [CrossRef]
- Thompson, J.; Scoones, I. Addressing the dynamics of agri-food systems: An emerging agenda for social science research. Environ. Sci. Policy 2009, 12, 386–397. [Google Scholar] [CrossRef]
- Schiere, J.B.; Grasman, J. Agro-ecosystem health: Aggregation of systems in time and space. In Proceedings of the Seminar on Agro-Ecosystem Health, Wageningen, The Netherlands, 26 September 1996; Volume 26, pp. 22–36. [Google Scholar]
- Scoones, I.; Leach, M.; Smith, A.; Stagl, S.; Stirling, A.; Thompson, J. Dynamic Systems and the Challenge of Sustainability; STEPS Working Paper 1; STEPS Centre: Brighton, UK, 2007. [Google Scholar]
- Almekinders, C.J.; Fresco, L.O.; Struik, P.C. The need to study and manage variation in agro-ecosystems. Neth. J. Agric. Sci. 1995, 43, 127–142. [Google Scholar] [CrossRef]
- Behnke, R.H.; Scoones, I. Rethinking Range Ecology: Implications for Rangeland Management in Africa; International Institute for Environment and Development: London, UK, 1992. [Google Scholar]
- Banson, K.E.; Nguyen, N.C.; Bosch, O.J. A systems thinking approach to the structure, conduct and performance of the agricultural sector in Ghana. Syst. Res. Behav. Sci. 2018, 35, 39–57. [Google Scholar] [CrossRef]
- Darnhofer, I.; Gibbon, D.; Dedieu, B. Farming Systems Research into the 21st Century: The New Dynamic; Springer Science & Business Media: London, UK, 2012. [Google Scholar]
- Reynolds, M.; Holwell, S. Systems Approaches to Making Change: A Practical Guide; Springer: London, UK, 2020. [Google Scholar] [CrossRef]
- Shongwe, M.I. A systems thinking approach to investigating complex sugarcane supply and processing systems: Integrating rich pictures and Bayesian networks. Syst. Pract. Action Res. 2018, 31, 75–85. [Google Scholar] [CrossRef]
- Teutscherová, N.; Vázquez, E.; Sotelo, M.; Villegas, D.; Velásquez, N.; Baquero, D.; Pulleman, M.M.; Arango, J. Intensive short-duration rotational grazing is associated with improved soil quality within one year after establishment in Colombia. Appl. Soil Ecol. 2021, 159, 103835. [Google Scholar] [CrossRef]
- di Virgilio, A.; Lambertucci, S.A.; Morales, J.M. Sustainable grazing management in rangelands: Over a century searching for a silver bullet. Agric. Ecosyst. Environ. 2019, 283, 106561. [Google Scholar] [CrossRef]
- Bell, S.; Berg, T.; Morse, S. Towards an understanding of rich picture interpretation. Syst. Pract. Action Res. 2019, 32, 601–614. [Google Scholar] [CrossRef]
- Armson, R. Drawing rich pictures. In Growing Wings on The Way; Triarchy Press Station Offices Axminster: Devon, UK, 2011; pp. 57–77. [Google Scholar]
- Conte, K.P.; Davidson, S. Using a ‘rich picture’ to facilitate systems thinking in research coproduction. Health Res. Policy Syst. 2020, 18, 14. [Google Scholar] [CrossRef]
- Gregorini, P.; Maxwell, T. Grazing in future multiscapes—From thoughtscapes to ethical and sustainable foodscapes. Journal 2020, 24, 23–25. [Google Scholar]
- Millennium, M.A. Ecosystem Assessment (MA) Synthesis Report; Millennium Ecosystem Assessment: Washington, DC, USA, 2005. [Google Scholar]
- Sandén, T.; Trajanov, A.; Spiegel, H.; Kuzmanovski, V.; Saby, N.; Picaud, C.; Henriksen, C.B.; Debeljak, M. Development of an agricultural primary productivity decision support model: A case study in France. Front. Environ. Sci. 2019, 7, 58. [Google Scholar] [CrossRef]
- Techen, A.K.; Helming, K. Pressures on soil functions from soil management in Germany. A foresight review. Agron. Sustain. Dev. 2017, 37, 64. [Google Scholar] [CrossRef]
- Lavelle, P.; Decaëns, T.; Aubert, M.; Barot, S.; Blouin, M.; Bureau, F.; Margerie, P.; Mora, P.; Rossi, J.P. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 2006, 42, S3–S15. [Google Scholar] [CrossRef]
- Pellant, M.; Shaver, P.; Pyke, D.A.; Herrick, J.E. Interpreting Indicators of Rangeland Health: Version 4; Technical Reference 1734-6; U.S. Department of the Interior, Bureau of Land Management, National Science and Technology Center: Denver, CO, USA, 2005; 122p.
- Teague, R.; Barnes, M. Grazing management that regenerates ecosystem function and grazing land livelihoods. Afr. J. Range Forage Sci. 2017, 34, 77–86. [Google Scholar] [CrossRef]
- Zhang, B.; Beck, R.; Pan, Q.; Zhao, M.; Hao, X. Soil physical and chemical properties in response to long-term cattle grazing on sloped rough fescue grassland in the foothills of the Rocky Mountains, Alberta. Geoderma 2019, 346, 75–83. [Google Scholar] [CrossRef]
- Hirsch, P.R.; Mauchline, T.H. The importance of the microbial N cycle in soil for crop plant nutrition. Adv. Appl. Microbiol. 2015, 93, 45–71. [Google Scholar] [PubMed]
- de Oliveira Silva, R.; Barioni, L.G.; Hall, J.A.J.; Folegatti Matsuura, M.; Zanett Albertini, T.; Fernandes, F.A.; Moran, D. Increasing beef production could lower greenhouse gas emissions in Brazil if decoupled from deforestation. Nat. Clim. Chang. 2016, 6, 493–497. [Google Scholar] [CrossRef]
- Bünemann, E.K.; Bongiorno, G.; Bai, Z.; Creamer, R.E.; De Deyn, G.; de Goede, R.; fleskens, L.; Geissen, V.; Mäde, P.; Pulleman, M.; et al. Soil quality–A critical review. Soil Biol. Biochem. 2018, 120, 105–125. [Google Scholar] [CrossRef]
- Karlen, D.L.; Mausbach, M.J.; Doran, J.W.; Cline, R.G.; Harris, R.F.; Schuman, G.E. Soil quality: A concept, definition, and framework for evaluation (a guest editorial). Soil Sci. Soc. Am. J. 1997, 61, 4–10. [Google Scholar] [CrossRef]
- Batey, T.; McKenzie, D.C. Soil compaction: Identification directly in the field. Soil Use Manag. 2006, 22, 123–131. [Google Scholar] [CrossRef]
- Mueller, L.; Schindler, U.; Mirschel, W.; Shepherd, T.G.; Ball, B.C.; Helming, K.; Rogasik, J.; Eulenstein, F.; Wiggering, H. Assessing the productivity function of soils. A review. Agron. Sustain. Dev. 2010, 30, 601–614. [Google Scholar] [CrossRef]
- Chen, Z.S.; Hsieh, C.F.; Jiang, F.Y.; Hsieh, T.H.; Sun, I.F. Relations of soil properties to topography and vegetation in a subtropical rain forest in southern Taiwan. Plant Ecol. 1997, 132, 229–241. [Google Scholar] [CrossRef]
- De, M.; Riopel, J.A.; Cihacek, L.J.; Lawrinenko, M.; Baldwin-Kordick, R.; Hall, S.J.; McDaniel, M.D. Soil health recovery after grassland reestablishment on cropland: The effects of time and topographic position. Soil Sci. Soc. Am. J. 2020, 84, 568–586. [Google Scholar] [CrossRef]
- Xue, R.; Yang, Q.; Miao, F.; Wang, X.; Shen, Y. Slope aspect influences plant biomass, soil properties and microbial composition in alpine meadow on the Qinghai-Tibetan plateau. J. Soil Sci. Plant Nutr. 2018, 18, 1–12. [Google Scholar] [CrossRef]
- Imeson, A.C.; Lavee, H. Soil erosion and climate change: The transect approach and the influence of scale. Geomorphology 1998, 23, 219–227. [Google Scholar] [CrossRef]
- Cerdà, A. Aggregate stability against water forces under different climates on agriculture land and scrubland in southern Bolivia. Soil Tillage Res. 2000, 57, 159–166. [Google Scholar] [CrossRef]
- Ziadat, F.M.; Taimeh, A.Y. Effect of rainfall intensity, slope, land use and antecedent soil moisture on soil erosion in an arid environment. Land Degrad. Dev. 2013, 24, 582–590. [Google Scholar] [CrossRef]
- Li, C.; Hao, X.; Zhao, M.; Han, G.; Willms, W.D. Influence of historic sheep grazing on vegetation and soil properties of a Desert Steppe in Inner Mongolia. Agric. Ecosyst. Environ. 2008, 128, 109–116. [Google Scholar] [CrossRef]
- Steffens, M.; Kölbl, A.; Totsche, K.U.; Kögel-Knabner, I. Grazing effects on soil chemical and physical properties in a semiarid steppe of Inner Mongolia (PR China). Geoderma 2008, 143, 63–72. [Google Scholar] [CrossRef]
- Piñeiro, G.; Paruelo, J.M.; Oesterheld, M.; Jobbágy, E.G. Pathways of grazing effects on soil organic carbon and nitrogen. Rangel. Ecol. Manag. 2010, 63, 109–119. [Google Scholar] [CrossRef]
- Le, Q.B.; Nkonya, E.; Mirzabaev, A. Biomass Productivity-Based Mapping of Global Land Degradation Hotspots. In Economics of Land Degradation and Improvement—A Global Assessment for Sustainable Development; Nkonya, E., Mirzabaev, A., Von Braun, J., Eds.; Springer Nature: Berlin/Heidelberg, Germany, 2016; pp. 55–84. [Google Scholar]
- Greenwood, K.L.; McKenzie, B.M. Grazing effects on soil physical properties and the consequences for pastures: A review. Aust. J. Exp. Agric. 2001, 41, 1231–1250. [Google Scholar] [CrossRef]
- Singleton, P.L.; Boyes, M.; Addison, B. Effect of treading by dairy cattle on topsoil physical conditions for six contrasting soil types in Waikato and Northland, New Zealand, with implications for monitoring. N. Z. J. Agric. Res. 2000, 43, 559–567. [Google Scholar] [CrossRef]
- Pietola, L.; Horn, R.; Yli-Halla, M. Effects of trampling by cattle on the hydraulic and mechanical properties of soil. Soil Tillage Res. 2005, 82, 99–108. [Google Scholar] [CrossRef]
- Pulido, M.; Schnabel, S.; Lavado Contador, J.F.; Lozano-Parra, J.; Gonzalez, F. The impact of heavy grazing on soil quality and pasture production in rangelands of SW Spain. Land Degrad. Dev. 2018, 29, 219–230. [Google Scholar] [CrossRef]
- Donovan, M.; Monaghan, R. Impacts of grazing on ground cover, soil physical properties and soil loss via surface erosion: A novel geospatial modelling approach. J. Environ. Manag. 2021, 287, 112206. [Google Scholar] [CrossRef] [PubMed]
- Scholefield, D.; Hall, D.M. A method to measure the susceptibility of pasture soils to poaching by cattle. Soil Use Manag. 1985, 1, 134–138. [Google Scholar] [CrossRef]
- Mayel, S.; Jarrah, M.; Kuka, K. How does grassland management affect physical and biochemical properties of temperate grassland soils? A review study. Grass Forage Sci. 2021, 76, 215–244. [Google Scholar] [CrossRef]
- Elliott, A.H.; Carlson, W.T. Effects of sheep grazing episodes on sediment and nutrient loss in overland flow. Soil Res. 2004, 42, 213–220. [Google Scholar] [CrossRef]
- da Silva, A.P.; Imhoff, S.; Corsi, M. Evaluation of soil compaction in an irrigated short-duration grazing system. Soil Tillage Res. 2003, 70, 83–90. [Google Scholar] [CrossRef]
- Zuazo, V.H.D.; Pleguezuelo, C.R.R. Soil-erosion and runoff prevention by plant covers: A review. Sustain. Agric. Res. 2009, 28, 785–811. [Google Scholar] [CrossRef]
- Taboada, M.A.; Rubio, G.; Chaneton, E.J. Grazing impacts on soil physical, chemical, and ecological properties in forage production systems. In Soil Management: Building a Stable Base for Agriculture; Hatfield, J.L., Sauer, T.L., Eds.; American Society of Agronomy and Soil Science Society of America: Madison, MI, USA, 2011; pp. 301–320. [Google Scholar] [CrossRef]
- Sayer, E.J. Using experimental manipulation to assess the roles of leaf litter in the functioning of forest ecosystems. Biol. Rev. 2006, 81, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Yates, C.J.; Norton, D.A.; Hobbs, R.J. Grazing effects on plant cover, soil and microclimate in fragmented woodlands in south-western Australia: Implications for restoration. Austral Ecol. 2000, 25, 36–47. [Google Scholar] [CrossRef]
- Schjønning, P.; van den Akker, J.J.; Keller, T.; Greve, M.H.; Lamandé, M.; Simojoki, A.; Stettler, M.; Arvidsson, J.; Breuning-Madsen, H. Driver-pressure-state-impact-response (DPSIR) analysis and risk assessment for soil compaction—A European perspective. Adv. Agron. 2015, 133, 183–237. [Google Scholar] [CrossRef]
- Hamilton, E.W., III; Frank, D.A. Can plants stimulate soil microbes and their own nutrient supply? Evidence from a grazing tolerant grass. Ecology 2001, 82, 2397–2402. [Google Scholar] [CrossRef]
- Holland, J.N.; Cheng, W.; Crossley, D. Herbivore-induced changes in plant carbon allocation: Assessment of below-ground C fluxes using carbon-14. Oecologia 1996, 107, 87–94. [Google Scholar] [CrossRef]
- Brevik, E.C.; Cerdà, A.; Mataix-Solera, J.; Pereg, L.; Quinton, J.N.; Six, J.; Van Oost, K. The interdisciplinary nature of SOIL. Soil 2015, 1, 117–129. [Google Scholar] [CrossRef]
- Nguyen, M.L.; Sheath, G.W.; Smith, C.M.; Cooper, A.B. Impact of cattle treading on hill land: 2. Soil physical properties and contaminant runoff. N. Z. J. Agric. Res. 1998, 41, 279–290. [Google Scholar] [CrossRef]
- Sahu, N.; Vasu, D.; Sahu, A.; Lal, N.; Singh, S.K. Strength of Microbes in Nutrient Cycling: A Key to Soil Health. In Agriculturally Important Microbes for Sustainable Agriculture; Meena, V., Mishra, P., Bisht, J., Pattanayak, A., Eds.; Springer: Singapore, 2017; pp. 69–86. [Google Scholar]
- Machado Filho, L.C.P.; Seó, H.L.; Daros, R.R.; Enriquez-Hidalgo, D.; Wendling, A.V.; Pinheiro Machado, L.C. Voisin rational grazing as a sustainable alternative for livestock production. Animals 2021, 11, 3494. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 2nd ed.; Academic Press: London, UK, 1997. [Google Scholar]
- Morriën, E.; Hannula, S.E.; Snoek, L.B.; Helmsing, N.R.; Zweers, H.; De Hollander, M.; Soto, R.J.; Bouffaud, M.-L.; Buée, M.; Dimmers, W.; et al. Soil networks become more connected and take up more carbon as nature restoration progresses. Nat. Commun. 2017, 8, 14349. [Google Scholar] [CrossRef]
- Xu, S.; Rowntree, J.; Borrelli, P.; Hodbod, J.; Raven, M.R. Ecological Health Index: A Short Term Monitoring Method for Land Managers to Assess Grazing Lands Ecological Health. Environments 2019, 6, 67. [Google Scholar] [CrossRef]
- Li, Z.; Xu, D.; Guo, X. Remote sensing of ecosystem health: Opportunities, challenges, and future perspectives. Sensors 2014, 14, 21117–21139. [Google Scholar] [CrossRef] [PubMed]
- Costanza, R.; Mageau, M. What is a healthy ecosystem? Aquat. Ecol. 1999, 33, 105–115. [Google Scholar] [CrossRef]
- Rapport, D.J.; Costanza, R.; McMichael, A.J. Assessing ecosystem health. Trends Ecol. Evol. 1998, 13, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Costanza, R. Toward an operational definition of ecosystem health. Ecosyst. Health New Goals Environ. Manag. 1992, 239, 269. [Google Scholar]
- Flood, P.J.; Harbinson, J.; Aarts, M.G. Natural genetic variation in plant photosynthesis. Trends Plant Sci. 2011, 16, 327–335. [Google Scholar] [CrossRef]
- Feng, X.; Dietze, M. Scale dependence in the effects of leaf ecophysiological traits on photosynthesis: Bayesian parameterization of photosynthesis models. New Phytol. 2013, 200, 1132–1144. [Google Scholar] [CrossRef]
- Parker, G.G. Tamm review: Leaf Area Index (LAI) is both a determinant and a consequence of important processes in vegetation canopies. For. Ecol. Manag. 2020, 477, 118496. [Google Scholar] [CrossRef]
- Hikosaka, K. Interspecific difference in the photosynthesis–nitrogen relationship: Patterns, physiological causes, and ecological importance. J. Plant Res. 2004, 117, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Wolf, D.D.; Carson, E.W.; Brown, R.H. Leaf area index and specific leaf area determinations. J. Agron. Educ. 1972, 1, 24–27. [Google Scholar] [CrossRef]
- Klumpp, K.; Soussana, J.F. Using functional traits to predict grassland ecosystem change: A mathematical test of the response-and-effect trait approach. Glob. Change Biol. 2009, 15, 2921–2934. [Google Scholar] [CrossRef]
- McSherry, M.E.; Ritchie, M.E. Effects of grazing on grassland soil carbon: A global review. Glob. Change Biol. 2013, 19, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Sundqvist, M.K.; Sanders, N.J.; Wardle, D.A. Community and ecosystem responses to elevational gradients: Processes, mechanisms, and insights for global change. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 261–280. [Google Scholar] [CrossRef]
- Kumar, M.; Kumar, S.; Sheikh, M.A. Effect of altitudes on soil and vegetation characteristics of Pinus roxburghii forest in Garhwal Himalaya. J. Adv. Lab. Res. Biol. 2010, 1, 130–133. [Google Scholar]
- Yang, J.; El-Kassaby, Y.A.; Guan, W. The effect of slope aspect on vegetation attributes in a mountainous dry valley, Southwest China. Sci. Rep. 2020, 10, 16465. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, K.F. Stability and instability of ecological systems in New Zealand mountains. Mt. Res. Dev. 1984, 4, 15–29. [Google Scholar] [CrossRef]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Barboni, D.; Harrison, S.P.; Bartlein, P.J.; Jalut, G.; New, M.; Prentice, I.C.; Stevenson, A.C. Relationships between plant traits and climate in the Mediterranean region: A pollen data analysis. J. Veg. Sci. 2004, 15, 635–646. [Google Scholar] [CrossRef]
- Mark, A.F. Effects of burning and grazing on sustainable utilization of upland snow tussock (Chionochloa spp.) rangelands for pastoralism in South Island, New Zealand. Aust. J. Bot. 1994, 42, 149–161. [Google Scholar] [CrossRef]
- Mueller, K.E.; Tilman, D.; Fornara, D.A.; Hobbie, S.E. Root depth distribution and the diversity–productivity relationship in a long-term grassland experiment. Ecology 2013, 94, 787–793. [Google Scholar] [CrossRef]
- Faucon, M.P.; Houben, D.; Lambers, H. Plant functional traits: Soil and ecosystem services. Trends Plant Sci. 2017, 22, 385–394. [Google Scholar] [CrossRef]
- Suter, M.; Huguenin-Elie, O.; Lüscher, A. Multispecies for multifunctions: Combining four complementary species enhances multifunctionality of sown grassland. Sci. Rep. 2021, 11, 3835. [Google Scholar] [CrossRef] [PubMed]
- Husse, S.; Lüscher, A.; Buchmann, N.; Hoekstra, N.J.; Huguenin-Elie, O. Effects of mixing forage species contrasting in vertical and temporal nutrient capture on nutrient yields and fertilizer recovery in productive grasslands. Plant Soil 2017, 420, 505–521. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Hallett, L.M.; Stein, C.; Suding, K.N. Functional diversity increases ecological stability in a grazed grassland. Oecologia 2017, 183, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Pott, D.M.; Osorio, S.; Vallarino, J.G. From central to specialized metabolism: An overview of some secondary compounds derived from the primary metabolism for their role in conferring nutritional and organoleptic characteristics to fruit. Front. Plant Sci. 2019, 10, 835. [Google Scholar] [CrossRef] [PubMed]
- Tajner-Czopek, A.; Gertchen, M.; Rytel, E.; Kita, A.; Kucharska, A.Z.; Sokół-Łętowska, A. Study of antioxidant activity of some medicinal plants having high content of caffeic acid derivatives. Antioxidants 2020, 9, 412. [Google Scholar] [CrossRef] [PubMed]
- Zduńska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant properties of ferulic acid and its possible application. Skin Pharmacol. Physiol. 2018, 31, 6. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.R.K.; Elghandour, M.M.M.Y.; Salem, A.Z.M.; Yasaswini, D.; Reddy, P.P.R.; Reddy, A.N.; Hyder, I. Plant secondary metabolites as feed additives in calves for antimicrobial 610 stewardship. Anim. Feed Sci. Technol. 2020, 264, 114469. [Google Scholar] [CrossRef]
- Al-Snafi, A.E. Medical benefit of Malva neglecta-A review. IOSR J. Pharm. 2019, 9, 60–67. [Google Scholar]
- Sabudak, T.; Orak, H.H.; Gulen, D.; Ozer, M.; Caliskan, H.; Bahrisefit, I.; Cabi, E. Investigation of some antibacterial and antioxidant properties of wild Cirsium vulgare from Turkey. Indian J. Pharm. Educ. Res. 2017, 51, 363. [Google Scholar] [CrossRef]
- Díaz, G.J.; Hernández, G.T.; Zamilpa, A.; Pérez, C.M.B.; Bribiesca, J.E.R.; Mendo, O.H.; Arroyo, H.S.; Cortazar, M.G.; Gives, P.M. In vitro assessment of Argemone mexicana, Taraxacum officinale, Ruta chalepensis and Tagetes filifolia against Haemonchus contortus nematode eggs and infective (L3) larvae. Microb. Pathog. 2017, 109, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Ganeshpurkar, A.; Saluja, A.K. The pharmacological potential of rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Vasta, V.; Luciano, G. The effects of dietary consumption of plants secondary compounds on small ruminants’ products quality. Small Rumin. Res. 2011, 101, 150–159. [Google Scholar] [CrossRef]
- Bhatta, R.; Uyeno, Y.; Tajima, K.; Takenaka, A.; Yabumoto, Y.; Nonaka, I.; Enishi, O.; Kurihara, M. Difference in the nature of tannins on in vitro ruminal methane and volatile fatty acid production and on methanogenic archaea and protozoal populations. J. Dairy Sci. 2009, 92, 5512–5522. [Google Scholar] [CrossRef] [PubMed]
- Peyraud, J.L.; Delagarde, R. Managing variations in dairy cow nutrient supply under grazing. Animal 2013, 7, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Orford, K.A.; Murray, P.J.; Vaughan, I.P.; Memmott, J. Modest enhancements to conventional grassland diversity improve the provision of pollination services. J. Appl. Ecol. 2016, 53, 906–915. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, P.C.; Nicolson, S.W.; Wright, G.A. Plant secondary metabolites in nectar: Impacts on pollinators and ecological functions. Funct. Ecol. 2017, 31, 65–75. [Google Scholar] [CrossRef]
- Lucas-Barbosa, D. Integrating studies on plant–pollinator and plant–herbivore interactions. Trends Plant Sci. 2016, 21, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Loreau, M.; Mouquet, N.; Gonzalez, A. Biodiversity as spatial insurance in heterogeneous landscapes. Proc. Natl Acad. Sci. USA 2003, 100, 12765–12770. [Google Scholar] [CrossRef] [PubMed]
- Milestad, R.; Dedieu, B.; Darnhofer, I.; Bellon, S. Farms and farmers facing change: The adaptive approach. In Farming Systems Research into the 21st Century: The New Dynamic; Darnhofer, I., Gibbon, D., Dedieu, B., Eds.; Springer Science & Business Media: London, UK, 2012; pp. 365–385. [Google Scholar]
- Allan, E.; Manning, P.; Alt, F.; Binkenstein, J.; Blaser, S.; Blüthgen, N.; Böhm, S.; Grassein, F.; Hölzel, N.; Klaus, V.H.; et al. Land use intensification alters ecosystem multifunctionality via loss of biodiversity and changes to functional composition. Ecol. Lett. 2015, 18, 834–843. [Google Scholar] [CrossRef] [PubMed]
- Lehman, C.L.; Tilman, D. Biodiversity, stability, and productivity in competitive communities. Am. Nat. 2000, 156, 534–552. [Google Scholar] [CrossRef]
- Van Coller, H.; Siebert, F.; Scogings, P.F.; Ellis, S. Herbaceous responses to herbivory, fire and rainfall variability differ between grasses and forbs. S. Afr. J. Bot. 2018, 119, 94–103. [Google Scholar] [CrossRef]
- Smith, M.D. An ecological perspective on extreme climatic events: A synthetic definition and framework to guide future research. J. Ecol. 2011, 99, 656–663. [Google Scholar] [CrossRef]
- Hoover, D.L.; Knapp, A.K.; Smith, M.D. Resistance and resilience of a grassland ecosystem to climate extremes. Ecology 2014, 95, 2646–2656. [Google Scholar] [CrossRef]
- Huntly, N. Herbivores and the dynamics of communities and ecosystems. Annu. Rev. Ecol. Evol. Syst. 1991, 22, 477–503. [Google Scholar] [CrossRef]
- McNaughton, S.J. Compensatory plant growth as a response to herbivory. Oikos 1983, 40, 329–336. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; Van Der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Janzen, H.H. What place for livestock on a re-greening earth? Anim. Feed Sci. Technol. 2011, 166, 783–796. [Google Scholar] [CrossRef]
- Steinfeld, H.; Gerber, P. Livestock production and the global environment: Consume less or produce better? Proc. Natl. Acad. Sci. USA 2010, 107, 18237–18238. [Google Scholar] [CrossRef]
- Hobbs, N.T. Modification of ecosystems by ungulates. J. Wildl. Manag. 1996, 60, 695–713. [Google Scholar] [CrossRef]
- Berndt, A.; Tomkins, N.W. Measurement and mitigation of methane emissions from beef cattle in tropical grazing systems: A perspective from Australia and Brazil. Animal 2013, 7, 363–372. [Google Scholar] [CrossRef]
- Sanderman, J.; Hengl, T.; Fiske, G.J. Soil carbon debt of 12,000 years of human land use. Proc. Natl. Acad. Sci. USA 2017, 114, 9575–9580. [Google Scholar] [CrossRef]
- Pierrehumbert, R.T.; Eshel, G. Climate impact of beef: An analysis considering multiple time scales and production methods without use of global warming potentials. Environ. Res. Lett. 2015, 10, 085002. [Google Scholar] [CrossRef]
- Grange, G.; Brophy, C.; Finn, J. Effects of plant diversity on yield in intensively managed grasslands. In Proceedings of the Joint 20th Symposium of the European Grassland Federation and the 33rd Meeting of the EUCARPIA Section ‘Fodder Crops and Amenity Grasses’, Zurich, Switzerland, 24–27 June 2019; Huguenin-Elie, O., Studer, B., Kolliker, R., Reheul, D., Probo, M., Barre, P., Feuerstein, U., Roldan-Ruiz, I., Mariotte, P., Hopkins, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; pp. 54–56. [Google Scholar]
- Nyfeler, D.; Huguenin-Elie, O.; Suter, M.; Frossard, E.; Connolly, J.; Lüscher, A. Strong mixture effects among four species in fertilized agricultural grassland led to persistent and consistent transgressive overyielding. J. Appl. Ecol. 2009, 46, 683–691. [Google Scholar] [CrossRef]
- Suter, M.; Connolly, J.; Finn, J.A.; Loges, R.; Kirwan, L.; Sebastià, M.T.; Lüscher, A. Nitrogen yield advantage from grass–legume mixtures is robust over a wide range of legume proportions and environmental conditions. Glob. Change Biol. 2015, 21, 2424–2438. [Google Scholar] [CrossRef]
- Teague, R.; Provenza, F.; Kreuter, U.; Steffens, T.; Barnes, M. Multi-paddock grazing on rangelands: Why the perceptual dichotomy between research results and rancher experience? J. Environ. Manag. 2013, 128, 699–717. [Google Scholar] [CrossRef] [PubMed]
- Meier, J.S.; Kreuzer, M.; Marquardt, S. Design and methodology of choice feeding experiments with ruminant livestock. Appl. Anim. Behav. Sci. 2012, 140, 105–120. [Google Scholar] [CrossRef]
- Beck, M.R.; Gregorini, P. How dietary diversity enhances hedonic and eudaimonic well-being in grazing ruminants. Front. Vet. Sci. 2020, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Villalba, J.J.; Provenza, F.D.; Catanese, F.; Distel, R.A. Understanding and manipulating diet choice in grazing animals. Anim. Prod. Sci. 2015, 55, 261–271. [Google Scholar] [CrossRef]
- Searle, K.R.; Hobbs, N.T.; Gordon, I.J. It’s the “foodscape”, not the landscape: Using foraging behavior to make functional assessments of landscape condition. Isr. J. Ecol. Evol. 2007, 53, 297–316. [Google Scholar] [CrossRef]
- Westoby, M. What are the biological bases of varied diets? Am. Nat. 1978, 112, 627–631. [Google Scholar] [CrossRef]
- Freeland, W.J.; Janzen, D.H. Strategies in herbivory by mammals: The role of plant secondary compounds. Am. Nat. 1974, 108, 269–289. [Google Scholar] [CrossRef]
- Provenza, F.D.; Kronberg, S.L.; Gregorini, P. Is grassfed meat and dairy better for human and environmental health? Front. Nutr. 2019, 6, 26. [Google Scholar] [CrossRef]
- Archimède, H.; Rira, M.; Eugène, M.; Fleury, J.; Lastel, M.L.; Periacarpin, F.; Silou-Etienne, T.; Morgavi, D.P.; Doreau, M. Intake, total-tract digestibility and methane emissions of Texel and Blackbelly sheep fed C4 and C3 grasses tested simultaneously in a temperate and a tropical area. J. Clean. Prod. 2018, 185, 455–463. [Google Scholar] [CrossRef]
- Gregorini, P.; Villalba, J.J.; Chilibroste, P.; Provenza, F.D. Grazing management: Setting the table, designing the menu and influencing the diner. Anim. Prod. Sci. 2017, 57, 1248–1268. [Google Scholar] [CrossRef]
- Pringle, H.J.; Landsberg, J. Predicting the distribution of livestock grazing pressure in rangelands. Austral Ecol. 2004, 29, 31–39. [Google Scholar] [CrossRef]
- di Virgilio, A.; Morales, J.M. Towards evenly distributed grazing patterns: Including social context in sheep management strategies. PeerJ 2016, 4, e2152. [Google Scholar] [CrossRef]
- Retallack, G.J. Global cooling by grassland soils of the geological past and near future. Annu. Rev. Earth Planet. Sci. 2013, 41, 69–86. [Google Scholar] [CrossRef]
- Voisin, A. Produtividade do Pasto; Mestre Jou: São Paulo, Brazil, 1974. [Google Scholar]
- Pereira, F.C.; Machado Filho, L.C.P.; Kazama, D.C.S.; Júnior, R.G.; Pereira, L.G.R.; Enríquez-Hidalgo, D. Effect of recovery period of mixture pasture on cattle behaviour, pasture biomass production and pasture nutritional value. Animal 2020, 14, 1961–1968. [Google Scholar] [CrossRef] [PubMed]
- Stanley, P.L.; Rowntree, J.E.; Beede, D.K.; DeLonge, M.S.; Hamm, M.W. Impacts of soil carbon sequestration on life cycle greenhouse gas emissions in Midwestern USA beef finishing systems. Agric. Syst. 2018, 162, 249–258. [Google Scholar] [CrossRef]
- Seó, H.L.S.; Machado Filho, L.C.P.; Brugnara, D. Rationally Managed Pastures Stock More Carbon than No-Tillage Fields. Front. Environ. Sci. 2017, 5, 87. [Google Scholar] [CrossRef]
- Frank, D.A.; Wallen, R.L.; Hamilton, E.W., III; White, P.J.; Fridley, J.D. Manipulating the system: How large herbivores control bottom-up regulation of grasslands. J. Ecol. 2018, 106, 434–443. [Google Scholar] [CrossRef]
- Hamilton, E.W., III; Frank, D.A.; Hinchey, P.M.; Murray, T.R. Defoliation induces root exudation and triggers positive rhizospheric feedbacks in a temperate grassland. Soil. Biol. Biochem. 2008, 40, 2865–2873. [Google Scholar] [CrossRef]
- Laliberté, E.; Tylianakis, J.M. Cascading effects of long-term land-use changes on plant traits and ecosystem functioning. Ecology 2012, 93, 145–155. [Google Scholar] [CrossRef]
- Wilson, C.H.; Strickland, M.S.; Hutchings, J.A.; Bianchi, T.S.; Flory, S.L. Grazing enhances belowground carbon allocation, microbial biomass, and soil carbon in a subtropical grassland. Glob. Change Biol. 2018, 24, 2997–3009. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, J.A.; Svenning, J.C.; Georgiou, K.; Malhi, Y. Can large herbivores enhance ecosystem carbon persistence? Trends Ecol. Evol. 2021, 37, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Bardgett, R.D.; Wardle, D.A. Herbivore-mediated linkages between aboveground and belowground communities. Ecology 2003, 84, 2258–2268. [Google Scholar] [CrossRef]
- Gregorini, P.; Tamminga, S.; Gunter, S.A. Review: Behavior and daily grazing patterns of cattle. Prof. Anim. Sci. 2006, 22, 201–209. [Google Scholar] [CrossRef]
- Gregorini, P.; Clark, C.; McLeod, K.; Glassey, C.; Romera, A.; Jago, J. Feeding station behavior of grazing dairy cows in response to restriction of time at pasture. Livest. Sci. 2011, 137, 287–291. [Google Scholar] [CrossRef]
- Villalba, J.J.; Provenza, F.D. Self-medication and homeostatic behaviour in herbivores: Learning about the benefits of nature’s pharmacy. Animal 2007, 1, 1360–1370. [Google Scholar] [CrossRef] [PubMed]
- Provenza, F.D. Postingestive feedback as an elementary determinant of food preference and intake in ruminants. Rangel. Ecol. Manag. 1995, 48, 2–17. [Google Scholar] [CrossRef]
- Chapman, D.F.; Parsons, A.J.; Cosgrove, G.P.; Barker, D.J.; Marotti, D.M.; Venning, K.J.; Rutter, S.M.; Hill, J.; Thompson, A.N. Impacts of spatial patterns in pasture on animal grazing behavior, intake, and performance. Crop Sci. 2007, 47, 399–415. [Google Scholar] [CrossRef]
- Pereira, F.C.; Machado Filho, L.C.P.; Kazama, D.C.D.S.; Guimarães, R., Jr. Black oat grown with common vetch improves the chemical composition and degradability rate of forage. Acta Sci. 2020, 42, e49951. [Google Scholar] [CrossRef]
- Provenza, F.D.; Pfister, J.A.; Cheney, C.D. Mechanisms of learning in diet selection with reference to phytotoxicosis in herbivores. Rangel. Ecol. Manag. 1992, 45, 36–45. [Google Scholar] [CrossRef]
- Hidiroglou, M. Trace element deficiencies and fertility in ruminants: A review. J. Dairy Sci. 1979, 62, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Villalba, J.J.; Provenza, F.D.; Manteca, X. Links between ruminants’ food preference and their welfare. Animal 2010, 4, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Manteca, X.; Villalba, J.J.; Atwood, S.B.; Dziba, L.; Provenza, F.D. Is dietary choice important to animal welfare? J. Vet. Behav. 2008, 3, 229–239. [Google Scholar] [CrossRef]
- Provenza, F.D. Acquired aversions as the basis for varied diets of ruminants foraging on rangelands. J. Anim. Sci. 1996, 74, 2010–2020. [Google Scholar] [CrossRef]
- Villalba, J.J.; Manteca, X. A case for eustress in grazing animals. Front. Vet. Sci. 2019, 6, 303. [Google Scholar] [CrossRef] [PubMed]
- Raubenheimer, D. Tannic acid, protein, and digestible carbohydrate: Dietary imbalance and nutritional compensation in locusts. Ecology 1992, 73, 1012–1027. [Google Scholar] [CrossRef]
- Yoshihara, Y.; Mizuno, H.; Ogura, S.; Sasaki, T.; Sato, S. Increasing the number of plant species in a pasture improves the mineral balance of grazing beef cattle. Anim. Feed. Sci. Technol. 2013, 179, 138–143. [Google Scholar] [CrossRef]
- Bertoni, G.; Minuti, A.; Trevisi, E. Immune system, inflammation and nutrition in dairy cattle. Anim. Prod. Sci. 2015, 55, 943–948. [Google Scholar] [CrossRef]
- Kumar, B.; Manuja, A.; Aich, P. Stress and its impact on farm animals. Front. Biosci. 2012, 4, 1759–1767. [Google Scholar] [CrossRef] [PubMed]
- Soder, K.J.; Rook, A.J.; Sanderson, M.A.; Goslee, S.C. Interaction of plant species diversity on grazing behavior and performance of livestock grazing temperate region pastures. Crop Sci. 2007, 47, 416–425. [Google Scholar] [CrossRef]
- Johnson, K.A.; Johnson, D.E. Methane emissions from cattle. J. Anim. Sci. 1995, 73, 2483–2492. [Google Scholar] [CrossRef]
- Benchaar, C.; Pomar, C.; Chiquette, J. Evaluation of dietary strategies to reduce methane production in ruminants: A modelling approach. Can. J. Anim. Sci. 2001, 81, 563–574. [Google Scholar] [CrossRef]
- Beauchemin, K.A.; Kreuzer, M.; O’mara, F.; McAllister, T.A. Nutritional management for enteric methane abatement: A review. Aust. J. Exp. Agric. 2008, 48, 21–27. [Google Scholar] [CrossRef]
- Haque, M.N. Dietary manipulation: A sustainable way to mitigate methane emissions from ruminants. J. Anim. Sci. Technol. 2018, 60, 15. [Google Scholar] [CrossRef]
- Castillo, A.R.; Kebreab, E.; Beever, D.E.; France, J. A review of efficiency of nitrogen utilisation in lactating dairy cows and its relationship with environmental pollution. J. Anim. Feed. Sci. 2000, 9, 1–32. [Google Scholar] [CrossRef]
- Wrage, N.; Strodthoff, J.; Cuchillo, H.M.; Isselstein, J.; Kayser, M. Phytodiversity of temperate permanent grasslands: Ecosystem services for agriculture and livestock management for diversity conservation. Biodivers. Conserv. 2011, 20, 3317–3339. [Google Scholar] [CrossRef]
- Fuke, G.; Nornberg, J.L. Systematic evaluation on the effectiveness of conjugated linoleic acid in human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Chen, H.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, Y.Q.; Chen, W. Review of the roles of conjugated linoleic acid in health and disease. J. Funct. Foods 2015, 15, 314–325. [Google Scholar] [CrossRef]
- Blondeau, N.; Lipsky, R.H.; Bourourou, M.; Duncan, M.W.; Gorelick, P.B.; Marini, A.M. Alpha-linolenic acid: An omega-3 fatty acid with neuroprotective properties—Ready for use in the stroke clinic? BioMed Res. Int. 2015, 2015, 519830. [Google Scholar] [CrossRef] [PubMed]
- Rajaram, S. Health benefits of plant-derived a-linolenic acid1–3. Am. J. Clin. Nutr. 2015, 100, 1. [Google Scholar] [CrossRef]
- Kolodziejczyk-Czepas, J. Trifolium species-derived substances and extracts—Biological activity and prospects for medicinal applications. J. Ethnopharmacol. 2012, 143, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Van Vliet, S.; Provenza, F.D.; Kronberg, S.L. Health-promoting phytonutrients are higher in grass-fed meat and milk. Front. Sustain. Food Syst. 2021, 4, 299. [Google Scholar] [CrossRef]
- Fraser, M.D.; Davies, D.A.; Vale, J.E.; Nute, G.R.; Hallett, K.G.; Richardson, R.I.; Wright, I.A. Performance and meat quality of native and continental cross steers grazing improved upland pasture or semi-natural rough grazing. Livest. Sci. 2009, 123, 70–82. [Google Scholar] [CrossRef]
- Renna, M.; Ferlay, A.; Lussiana, C.; Bany, D.; Graulet, B.; Wyss, U.; Enri, S.R.; Battaglini, L.M.; Coppa, M. Relative hierarchy of farming practices affecting the fatty acid composition of permanent grasslands and of the derived bulk milk. Anim. Feed. Sci. Technol. 2020, 267, 114561. [Google Scholar] [CrossRef]
- Kuhnen, S.; Holz, D.T.; Moacyr, J.R.; Piccinin, I.N.; Pinheiro Machado Filho, L.C. Effect of pasture management on bioactive compounds of Lolium multiflorum and Avena strigosa for dairy cows and its effect on milk quality. Agroecol. Sustain. Food Syst. 2022, 46, 3–22. [Google Scholar] [CrossRef]
- Villalba, J.J.; Bach, A.; Ipharraguerre, I.R. Feeding behavior and performance of lambs are influenced by flavor diversity. J. Anim. Sci. 2011, 89, 2571–2581. [Google Scholar] [CrossRef] [PubMed]
- Torres-Fajardo, R.A.; González-Pech, P.G.; Sandoval-Castro, C.A.; Torres-Acosta, J.F.D.J. Small ruminant production based on rangelands to optimize animal nutrition and health: Building an interdisciplinary approach to evaluate nutraceutical plants. Animals 2020, 10, 1799. [Google Scholar] [CrossRef] [PubMed]
- Villalba, J.J.; Miller, J.; Ungar, E.D.; Landau, S.Y.; Glendinning, J. Ruminant self-medication against gastrointestinal nematodes: Evidence, mechanism, and origins. Parasite 2014, 21, 31. [Google Scholar] [CrossRef]
- Holechek, J.L. Considerations concerning grazing systems. Rangel. Arch. 1983, 5, 208–211. [Google Scholar]
- Hawkins, H.J. A global assessment of Holistic Planned Grazing™ compared with season-long, continuous grazing: Meta-analysis findings. Afr. J. Range Forage Sci. 2017, 34, 65–75. [Google Scholar] [CrossRef]
- Dahal, S.; Franklin, D.; Subedi, A.; Cabrera, M.; Hancock, D.; Mahmud, K.; Ney, L.; Park, C.; Mishra, D. Strategic grazing in beef-pastures for improved soil health and reduced runoff-nitrate-a step towards sustainability. Sustainability 2020, 12, 558. [Google Scholar] [CrossRef]
- Drewry, J.J. Natural recovery of soil physical properties from treading damage of pastoral soils in New Zealand and Australia: A review. Agric. Ecosyst. Environ. 2006, 114, 159–169. [Google Scholar] [CrossRef]
- Badgery, W.; Millar, G.; Broadfoot, K.; Martin, J.; Pottie, D.; Simmons, A.; Cranney, P. Better management of intensive rotational grazing systems maintains pastures and improves animal performance. Crop Pasture Sci. 2017, 68, 1131–1140. [Google Scholar] [CrossRef]
- Török, P.; Penksza, K.; Tóth, E.; Kelemen, A.; Sonkoly, J.; Tóthmérész, B. Vegetation type and grazing intensity jointly shape grazing effects on grassland biodiversity. Ecol. Evol. 2018, 8, 10326–10335. [Google Scholar] [CrossRef]
- Herrero-Jáuregui, C.; Oesterheld, M. Effects of grazing intensity on plant richness and diversity: A meta-analysis. Oikos 2018, 127, 757–766. [Google Scholar] [CrossRef]
- Owen-Smith, N. Credible models for herbivore-vegetation systems: Towards an ecology of equations: Starfield Festschrift. S. Afr. J. Sci. 2002, 98, 445–449. [Google Scholar]
- Owen-Smith, N. Functional heterogeneity in resources within landscapes and herbivore population dynamics. Landsc. Ecol. 2004, 19, 761–771. [Google Scholar] [CrossRef]
- Conway, G.R. Agroecosystem analysis. Agric. Adm. 1985, 20, 31–55. [Google Scholar] [CrossRef]
- Johnson, P.D.; Besselsen, D.G. Practical aspects of experimental design in animal research. ILAR J. 2002, 43, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Sauvant, D.; Letourneau-Montminy, M.P.; Schmidely, P.; Boval, M.; Loncke, C.; Daniel, J.B. Use and misuse of meta-analysis in Animal Science. Animal 2020, 14, s207–s222. [Google Scholar] [CrossRef] [PubMed]
- Bryant, J.R.; Snow, V.O. Modelling pastoral farm agro-ecosystems: A review. N. Z. J. Agric. Res. 2008, 51, 349–363. [Google Scholar] [CrossRef]
- Barnes, M.K.; Steffens, T.J.; Rittenhouse, L.R. Grazing period stocking rate drives livestock performance in rotational stocking. In Proceedings of the 9th International Rangeland Congress: Diverse Rangelands for a Sustainable Society, Rosario, Argentina, 3–10 April 2011; Feldman, S.R., Oliva, G.E., Sacido, M.B., Eds.; United States Department of Agriculture: Washington, DC, USA, 2011; p. 633. [Google Scholar]
- Rawat, J.S.; Kumar, M. Monitoring land use/cover change using remote sensing and GIS techniques: A case study of Hawalbagh block, district Almora, Uttarakhand, India. Egypt. J. Remote Sens. Space Sci. 2015, 18, 77–84. [Google Scholar] [CrossRef]
- Gashaw, T.; Bantider, A.; Mahari, A. Evaluations of land use/land cover changes and land degradation in Dera District, Ethiopia: GIS and remote sensing based analysis. Int. J. Sci. Res. Environ. Sci. 2014, 2, 199. [Google Scholar] [CrossRef]
- Nath, S.S.; Bolte, J.P.; Ross, L.G.; Aguilar-Manjarrez, J. Applications of geographical information systems (GIS) for spatial decision support in aquaculture. Aquac. Eng. 2000, 23, 233–278. [Google Scholar] [CrossRef]
- Moore, A.; Johnson, M.; Lord, J.; Coutts, S.; Pagan, M.; Gbolagun, J.; Hall, G.B. Applying spatial analysis to the agroecology-led management of an indigenous farm in New Zealand. Ecol. Inform. 2016, 31, 49–58. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, F.C.; Smith, C.M.S.; Charters, S.M.; Gregorini, P. Creating a Design Framework to Diagnose and Enhance Grassland Health under Pastoral Livestock Production Systems. Animals 2022, 12, 3306. https://doi.org/10.3390/ani12233306
Pereira FC, Smith CMS, Charters SM, Gregorini P. Creating a Design Framework to Diagnose and Enhance Grassland Health under Pastoral Livestock Production Systems. Animals. 2022; 12(23):3306. https://doi.org/10.3390/ani12233306
Chicago/Turabian StylePereira, Fabiellen C., Carol M. S. Smith, Stuart M. Charters, and Pablo Gregorini. 2022. "Creating a Design Framework to Diagnose and Enhance Grassland Health under Pastoral Livestock Production Systems" Animals 12, no. 23: 3306. https://doi.org/10.3390/ani12233306
APA StylePereira, F. C., Smith, C. M. S., Charters, S. M., & Gregorini, P. (2022). Creating a Design Framework to Diagnose and Enhance Grassland Health under Pastoral Livestock Production Systems. Animals, 12(23), 3306. https://doi.org/10.3390/ani12233306