
Bioavailability of Different Zinc Sources in Pigs 0–3 Weeks Post-Weaning

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Housing and Experimental Diets
2.2. Registrations and Samples
2.3. Analysis of Feed, Faeces and Blood
2.4. Calculation of the Total Tract Apparent Digestibility of Zn
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EU. Commission, Commission Regulation (EC) No 1334/2003 of 25 July 2003 amending the conditions for authorisation of a number of additives in feedingstuffs belonging to the group of trace elements. Off. J. Eur. Union 2003, 187, 11. [Google Scholar]
- EU. Commission implementing regulation (EU) 2016/1095 of 6 July 2016 concerning the authorization of zinc acetate dihydrate, zinc chloride anhydrous, zinx oxide, zinc sulphate heptahydrate, zinc sulphate monohydrate, zinc chelate of anmino acids hydrate, zinc chelate of protein hydrolysates, zinc chelate of glycine hydrate (solid) and zincchelate of glycine hydrate (liquid) as feed additives for all animal species and amending regulations (EC). No 1334/2003, (EC) No 479/2006, (EU) No 335/2010 and Implementing Regulations (EU) No 991/2012 and (EU) No 636/2013. Off. J. Eur. Union 2016, 182, 7–27. [Google Scholar]
- Poulsen, H.D.; Larsen, T. Zinc excretion and retention in growing pigs fed increasing levels of zinc oxide. Livest. Prod. Sci. 1995, 43, 235–242. [Google Scholar] [CrossRef]
- Brugger, D.; Windisch, W.M. Zn metabolism of monogastric species and consequences for the definition of feeding requirements and the estimation of feed Zn bioavailability. J. Zhejiang Univ. Sci. B 2019, 20, 617–627. [Google Scholar] [CrossRef]
- Brugger, D.; Buffler, M.; Windisch, W. Development of an experimental model to assess the bioavailability of zinc in practical piglet diets. Arch. Anim. Nutr. 2014, 68, 73–92. [Google Scholar] [CrossRef]
- Lönnerdal, B. Dietary factors influencing zinc absorption. J. Nutr. 2000, 130, 1378s–1383s. [Google Scholar] [CrossRef]
- Hambidge, K.M.; Miller, L.V.; Westcott, J.E.; Sheng, X.; Krebs, N.F. Zinc bioavailability and homeostasis. Am. J. Clin. Nutr. 2010, 91, 1478s–1483s. [Google Scholar] [CrossRef]
- Ollig, J.; Kloubert, V.; Weßels, I.; Haase, H.; Rink, L. Parameters Influencing Zinc in Experimental Systems in Vivo and in Vitro. Metals 2016, 6, 71. [Google Scholar] [CrossRef]
- Schlegel, P.; Nys, Y.; Jondreville, C. Zinc availability and digestive zinc solubility in piglets and broilers fed diets varying in their phytate contents, phytase activity and supplemented zinc source. Animal 2010, 4, 200–209. [Google Scholar] [CrossRef]
- Miller, L.V.; Krebs, N.F.; Hambidge, K.M. A mathematical model of zinc absorption in humans as a function of dietary zinc and phytate. J. Nutr. 2007, 137, 135–141. [Google Scholar] [CrossRef]
- Bikker, P.; Jongbloed, A.W.; Thissen, J.T. Meta-analysis of effects of microbial phytase on digestibility and bioavailability of copper and zinc in growing pigs. J. Anim. Sci. 2012, 90 (Suppl. 4), 134–136. [Google Scholar] [CrossRef] [PubMed]
- Revy, P.S.; Jondreville, C.; Dourmad, J.Y.; Nys, Y. Effect of zinc supplemented as either an organic or an inorganic source and of microbial phytase on zinc and other minerals utilisation by weanling pigs. Anim. Feed Sci. Technol. 2004, 116, 93–112. [Google Scholar] [CrossRef]
- Lopez, H.W.; Levrat-Verny, M.-A.; Coudray, C.; Besson, C.; Krespine, V.; Messager, A.; Demigné, C.; RéMéSy, C. Class 2 resistant starches lower plasma and liver lipids and improve mineral retention in rats. J. Nutr. 2001, 131, 1283–1289. [Google Scholar] [CrossRef] [PubMed]
- Nitrayova, S.; Windisch, W.; von Heimendahl, E.; Müller, A.; Bartelt, J. Bioavailability of zinc from different sources in pigs. J. Anim. Sci. 2012, 90 (Suppl. 4), 185–187. [Google Scholar] [CrossRef]
- Oh, H.-J.; Park, Y.-J.; Cho, J.H.; Song, M.-H.; Gu, B.-H.; Yun, W.; Lee, J.-H.; An, J.-S.; Kim, Y.-J.; Lee, J.-S.; et al. Changes in diarrhea score, nutrient digestibility, zinc utilization, intestinal immune profiles, and fecal microbiome in weaned piglets by different forms of zinc. Animals 2021, 11, 1356. [Google Scholar] [CrossRef]
- Jou, M.Y.; Philipps, A.F.; Kelleher, S.L.; Lönnerdal, B. Effects of zinc exposure on zinc transporter expression in human intestinal cells of varying maturity. J. Pediatr. Gastroenterol. Nutr. 2010, 50, 587–595. [Google Scholar] [CrossRef]
- Byrne, L.; Hynes, M.J.; Connolly, C.D.; Murphy, R.A. Influence of the chelation process on the stability of organic trace mineral supplements used in animal nutrition. Animals 2021, 11, 1730. [Google Scholar] [CrossRef]
- Diao, H.; Yan, J.; Li, S.; Kuang, S.; Wei, X.; Zhou, M.; Zhang, J.; Huang, C.; He, P.; Tang, W. Effects of dietary zinc sources on growth performance and gut health of weaned piglets. Front. Microbiol. 2021, 12, 771617. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, Q.; Wang, L.; Wang, Y.; Cheng, Z.; Yang, Z.; Yang, W. The effects of partially or completely substituted dietary zinc sulfate by lower levels of zinc methionine on growth performance, apparent total tract igestibility, immune function, and visceral indices in weaned piglets. Animals 2019, 9, 236. [Google Scholar] [CrossRef]
- Peng, P.; Chen, J.; Yao, K.; Yin, Y.; Long, L.; Fang, R. The effects of dietary supplementation with porous zinc oxide on growth performance, intestinal microbiota, morphology, and permeability in weaned piglets. Anim. Sci. J. 2019, 90, 1220–1228. [Google Scholar] [CrossRef]
- Zhang, Y.; Ward, T.L.; Ji, F.; Peng, C.; Zhu, L.; Gong, L.; Dong, B. Effects of zinc sources and levels of zinc amino acid complex on growth performance, hematological and biochemical parameters in weanling pigs. Asian-Australas J. Anim. Sci. 2018, 31, 1267–1274. [Google Scholar] [CrossRef] [PubMed]
- Tybirk, P.; Sloth, N.M.; Kjeldsen, N.; Weber, N.R. Danish Nutrient Standards, 30th ed.; SEGES Svineproduktion: Copenhagen, Denmark, 2020. [Google Scholar]
- Petersen, L.; Minkkinen, P.; Esbensen, K.H. Representative sampling for reliable data analysis: Theory of Sampling. Chemom. Intell. Lab. Syst. 2005, 77, 261–277. [Google Scholar] [CrossRef]
- Pedersen, K.S.; Toft, N. Intra- and inter-observer agreement when using a descriptive classification scale for clinical assessment of faecal consistency in growing pigs. Prev. Vet. Med. 2011, 98, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.V.; Nørskov, N.P.; Nørgaard, J.V.; Woyengo, T.A.; Poulsen, H.D.; Nielsen, T.S. Determination of the optimal level of dietary zinc for newly weaned pigs: A dose-response study. Animals 2022, 12, 1552. [Google Scholar] [CrossRef]
- Myers, W.D.; Ludden, P.A.; Nayigihugu, V.; Hess, B.W. Technical note: A procedure for the preparation and quantitative analysis of samples for titanium dioxide. J. Anim. Sci. 2004, 82, 179–183. [Google Scholar] [CrossRef] [PubMed]
- National Research Council; Committee on Nutrient Requirements of Swine; Board on Agriculture and Natural Resources; Division on Earth and Life Studies. Nutrient Requirements of Swine, 11th ed.; The National Academics Press: Washington, DC, USA, 2012; ISBN 978-0-309-22423-9. [Google Scholar]
- Bruininx, E.M.A.M.; van der Peet-Schwering, C.M.C.; Schrama, J.W.; Vereijken, P.F.G.; Vesseur, P.C.; Everts, H.; den Hartog, L.A.; Beynen, A.C. Individually measured feed intake characteristics and growth performance of group-housed weanling pigs: Effects of sex, initial body weight, and body weight distribution within groups. J. Anim. Sci. 2001, 79, 301–308. [Google Scholar] [CrossRef]
- Lynegaard, J.C.; Kjeldsen, N.J.; Bache, J.K.; Weber, N.R.; Hansen, C.F.; Nielsen, J.P.; Amdi, C. A very low CP level reduced diarrhoea and productivity in weaner pigs, but no differences between post-weaning diets including soybean meal or soy protein concentrate were found. Animals 2021, 11, 678. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Scientific opinion on the characterisation of zinc compound ‘Zinc chelate of amino acids, hydrate (Availa®Zinc)’ as a feed additive for all animal species. EFSA J. 2013, 11, 3369. [Google Scholar] [CrossRef][Green Version]
- Sauer, A.K.; Pfaender, S.; Hagmeyer, S.; Tarana, L.; Mattes, A.-K.; Briel, F.; Küry, S.; Boeckers, T.M.; Grabrucker, A.M. Characterization of zinc amino acid complexes for zinc delivery in vitro using Caco-2 cells and enterocytes from hiPSC. BioMetals 2017, 30, 643–661. [Google Scholar] [CrossRef]
- Hill, D.A.; Peo, E.R., Jr.; Lewis, A.J.; Crenshaw, J.D. Zinc-amino acid complexes for swine. J. Anim. Sci. 1986, 63, 121–130. [Google Scholar] [CrossRef][Green Version]
- Wedekind, K.J.; Lewis, A.J.; Giesemann, M.A.; Miller, P.S. Bioavailability of zinc from inorganic and organic sources for pigs fed corn-soybean meal diets. J. Anim. Sci. 1994, 72, 2681–2689. [Google Scholar] [CrossRef] [PubMed]
- Dourmad, J.-Y.; Guinotte, F.O.; Nys, Y. Bioavailability of two sources of zinc in weanling pigs. Anim. Res. 2002, 51, 315–326. [Google Scholar] [CrossRef]
- Alkhtib, A.; Scholey, D.; Carter, N.; Cave, G.W.V.; Hanafy, B.I.I.; Kempster, S.R.J.; Mekapothula, S.; Roxborough, E.T.; Burton, E.J. Bioavailability of methionine-coated zinc nanoparticles as a dietary supplement leads to improved performance and bone strength in broiler chicken production. Animals 2020, 10, 1482. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, H.A.; Jalal, M.; Al-Titi, H.H.; Souad, A. Effect of sources and levels of dietary zinc on the performance, carcass traits and blood parameters of broilers. Rev. Bras. Ciência Avícola 2017, 19, 519–526. [Google Scholar] [CrossRef]
- Abd El-Hack, M.E.; Alagawany, M.; Salah, A.S.; Abdel-Latif, M.A.; Farghly, M.F.A. Effects of dietary supplementation of zinc oxide and zinc methionine on layer performance, egg quality, and blood serum indices. Biol. Trace Elem. Res. 2018, 184, 456–462. [Google Scholar] [CrossRef]
- King, J.C.; Brown, K.H.; Gibson, R.S.; Krebs, N.F.; Lowe, N.M.; Siekmann, J.H.; Raiten, D.J. Biomarkers of nutrition for development (BOND)-Zinc review. J. Nutr. 2015, 146, 858s–885s. [Google Scholar] [CrossRef]
- Lowe, N.M.; Fekete, K.; Decsi, T. Methods of assessment of zinc status in humans: A systematic review. Am. J. Clin. Nutr. 2009, 89, 2040s–2051s. [Google Scholar] [CrossRef]
- Carlson, D.; Beattie, J.H.; Poulsen, H.D. Assessment of zinc and copper status in weaned piglets in relation to dietary zinc and copper supply. J. Anim. Physiol. Anim. Nutr. 2007, 91, 19–28. [Google Scholar] [CrossRef]
- Burrough, E.R.; De Mille, C.; Gabler, N.K. Zinc overload in weaned pigs: Tissue accumulation, pathology, and growth impacts. J. Vet. Diagn. Investig. 2019, 31, 537–545. [Google Scholar] [CrossRef]


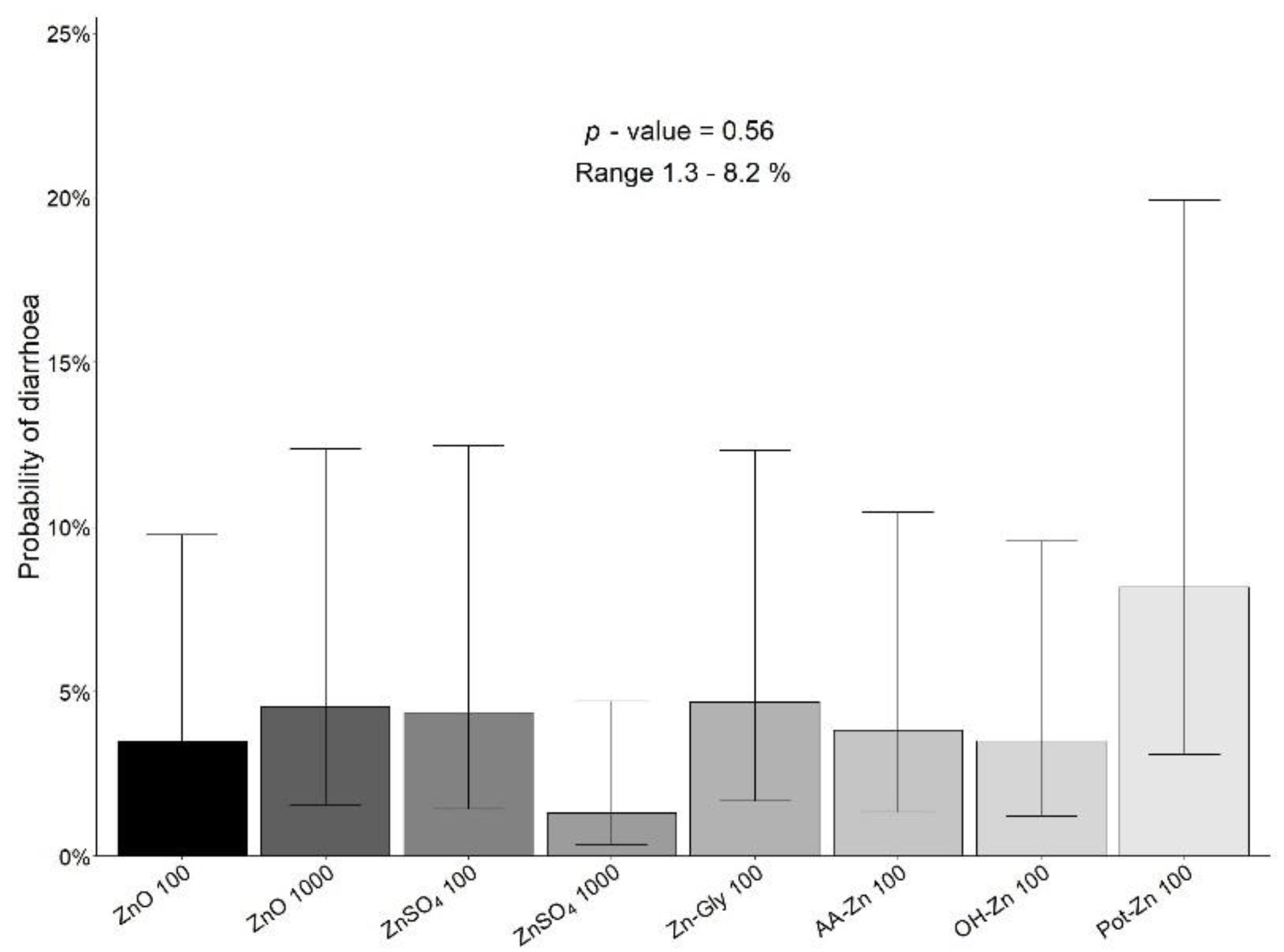{kind=link}
| Ingredients | Content (g/kg, as-Fed Basis) |
|---|---|
| Wheat | 563 |
| Barley | 200 |
| Premix 1 | 52 |
| Soy protein concentrate | 51 |
| Soybean meal | 50 |
| Fish meal | 30 |
| Potato protein | 27 |
| Soybean oil | 21 |
| Titanium dioxide | 5 |
| Aroma | 1 |
| Calculated Composition | |
| Starch | 432 |
| Crude protein | 176 |
| Fat | 45 |
| Ash | 53 |
| Soluble fibre | 28 |
| Insoluble fibre | 99 |
| Zn (mg/kg) | 27 |
| Fe (mg/kg) | 226 |
| Cu (mg/kg) | 179 |
| Diet | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| Zn Source | ZnO | ZnO | ZnSO4 | ZnSO4 | Zn-Gly | AA-Zn | OH-Zn | Pot-Zn |
| Declared Zn content in the source, % | 72 | 72 | 35 | 35 | 25 | 10 | 56 | 75 |
| Inclusion level, mg Zn/kg | 100 | 1000 | 100 | 1000 | 100 | 100 | 100 | 100 |
| Analysed total Zn 1, mg/kg as-is | 140 | 1044 | 149 | 1108 | 139 | 162 | 140 | 145 |
| Zinc Source | ZnO | ZnO | ZnSO4 | ZnSO4 | Zn-Gly | AA-Zn | OH-Zn | Pot-Zn | SEM | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|
| Inclusion Level, mg/kg | 100 | 1000 | 100 | 1000 | 100 | 100 | 100 | 100 | ||
| BW d 0, kg | 6.82 | 6.88 | 6.73 | 6.91 | 6.70 | 6.85 | 6.82 | 6.90 | 0.25 | 1.0 |
| Feed intake, g/d | ||||||||||
| d 0–7 | 112 | 79 | 151 | 125 | 78 | 121 | 131 | 148 | 23 | 0.16 |
| d 7–14 | 323 | 254 | 346 | 370 | 330 | 364 | 376 | 367 | 35 | 0.19 |
| d 14–21 | 573 | 444 | 583 | 563 | 506 | 592 | 543 | 566 | 42 | 0.19 |
| d 0–21 | 336 | 259 | 360 | 352 | 317 | 359 | 350 | 361 | 29 | 0.17 |
| Gain, g/d | ||||||||||
| d 0–7 | 46 | −2 | 99 | 50 | 23 | 56 | 78 | 91 | 37 | 0.50 |
| d 7–14 | 295 | 228 | 297 | 333 | 330 | 332 | 323 | 317 | 33 | 0.27 |
| d 14–21 | 504 | 379 | 490 | 462 | 442 | 529 | 387 | 443 | 45 | 0.22 |
| d 0–21 | 282 | 201 | 295 | 282 | 277 | 306 | 263 | 284 | 28 | 0.21 |
| Zinc Source | ZnO | ZnO | ZnSO4 | ZnSO4 | Zn-Gly | AA-Zn | OH-Zn | Pot-Zn | SEM | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|
| Inclusion Level, mg/kg | 100 | 1000 | 100 | 1000 | 100 | 100 | 100 | 100 | ||
| Zn intake, mg/d | ||||||||||
| d 0–7 1 | 10 c [6;16] | 50 b [30;82] | 20 c [12;34] | 105 a [64;172] | 10 c [6;17] | 18 c [11;29] | 14 c [9;23] | 18 c [11;31] | <0.001 | |
| d 7–14 1 | 43 c [35;53] | 230 b [187;284] | 50 c [40;62] | 393 a [319;484] | 43 c [35;53] | 58 c [47;71] | 48 c [39;60] | 51 c [41;64] | <0.001 | |
| d 14–21 1 | 80 b [67;94] | 422 a [357;499] | 86 b [72;103] | 594 a [502;702] | 69 b [58;81] | 95 b [80;112] | 71 b [60;84] | 81 b [68;97] | <0.001 | |
| d 0–21 1 | 46 c [39;55] | 246 b [207;292] | 53 c [44;63] | 375 a [316;445] | 41 c [35;49] | 58 c [49;68] | 45 c [38;54] | 51 c [42;61] | <0.001 | |
| Faecal Zn (g/kg DM) d 14 | 1.5 b | 6.4 a | 1.2 b | 6.8 a | 1.2 b | 1.1 b | 1.1 b | 1.0 b | 0.3 | <0.001 |
| ATTD of Zn d 14, % | −6.82 bcd | 3.42 ab | −9.56 bcd | 8.70 a | −16.48 d | −0.55 abc | −9.87 cd | −4.07 bc | 2.8 | <0.001 |
| Apparently digested Zn d 14, mg/kg feed intake | −9.50 bc | 34.0 b | −12.5 c | 96.4 a | −22.9 c | −0.90 bc | −13.7 c | −5.89 bc | 10.5 | <0.001 |
| Zinc Source | ZnO | ZnO | ZnSO4 | ZnSO4 | Zn-Gly | AA-Zn | OH-Zn | Pot-Zn | SEM | p-Value |
|---|---|---|---|---|---|---|---|---|---|---|
| Inclusion Level, mg/kg | 100 | 1000 | 100 | 1000 | 100 | 100 | 100 | 100 | ||
| Serum Zn, μg/L | ||||||||||
| d 0 | 795 | 869 | 819 | 867 | 835 | 882 | 860 | 890 | 54 | 0.91 |
| d 7 | 656 b | 883 b | 651 b | 1502 a | 610 b | 622 b | 630 b | 646 b | 83 | <0.001 |
| d 14 | 679 c | 1137 b | 660 c | 1643 a | 665 c | 684 c | 655 c | 648 c | 87 | <0.001 |
| d 21 | 757 c | 1754 b | 775 c | 2264 a | 788 c | 844 c | 811 c | 790 c | 117 | <0.001 |
| Δ Serum Zn, μg/L | ||||||||||
| d 0–7 | −139 b | 14 b | −168 b | 635 a | −224 b | −261 b | −230 b | −244 b | 98 | <0.001 |
| d 0–14 | −117 bc | 268 b | −159 bc | 776 a | −170 c | −198 c | −206 c | −242 c | 101 | <0.001 |
| d 0–21 | −39 b | 885 a | −44 b | 1397 a | −47 b | −38 b | −50 b | −100 b | 127 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nielsen, T.S.; Engelsmann, M.N.; Hansen, S.V.; Maribo, H. Bioavailability of Different Zinc Sources in Pigs 0–3 Weeks Post-Weaning. Animals 2022, 12, 2921. https://doi.org/10.3390/ani12212921
Nielsen TS, Engelsmann MN, Hansen SV, Maribo H. Bioavailability of Different Zinc Sources in Pigs 0–3 Weeks Post-Weaning. Animals. 2022; 12(21):2921. https://doi.org/10.3390/ani12212921
Chicago/Turabian StyleNielsen, Tina S., Maiken N. Engelsmann, Sally V. Hansen, and Hanne Maribo. 2022. "Bioavailability of Different Zinc Sources in Pigs 0–3 Weeks Post-Weaning" Animals 12, no. 21: 2921. https://doi.org/10.3390/ani12212921
APA StyleNielsen, T. S., Engelsmann, M. N., Hansen, S. V., & Maribo, H. (2022). Bioavailability of Different Zinc Sources in Pigs 0–3 Weeks Post-Weaning. Animals, 12(21), 2921. https://doi.org/10.3390/ani12212921