
Effects of Low-Ambient-Temperature Stimulation on Modifying the Intestinal Structure and Function of Different Pig Breeds

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Body Temperature and Complete Blood Counts
3.2. Duodenal Pathological Observations
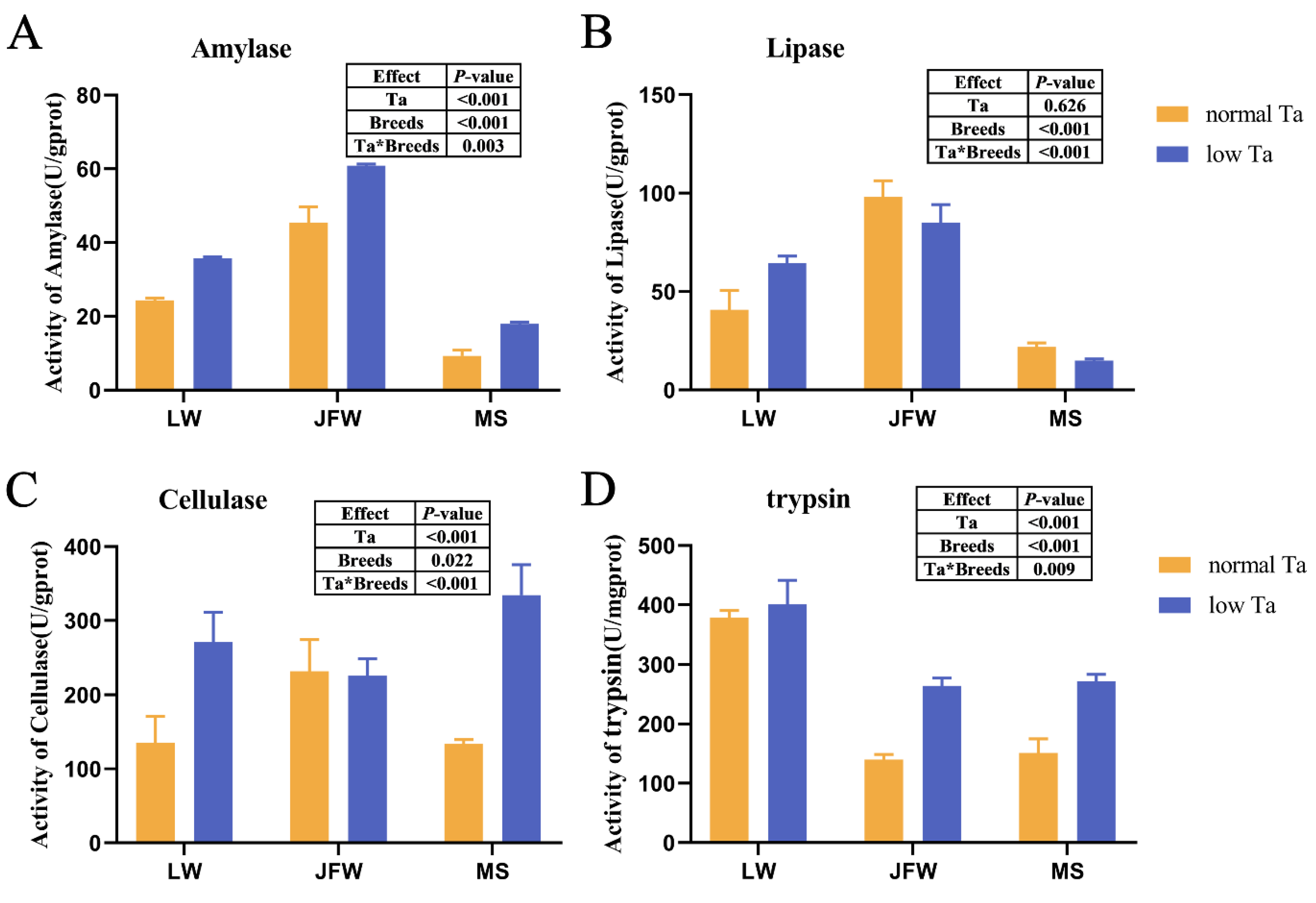
3.3. Changes in Digestive Enzyme Activity
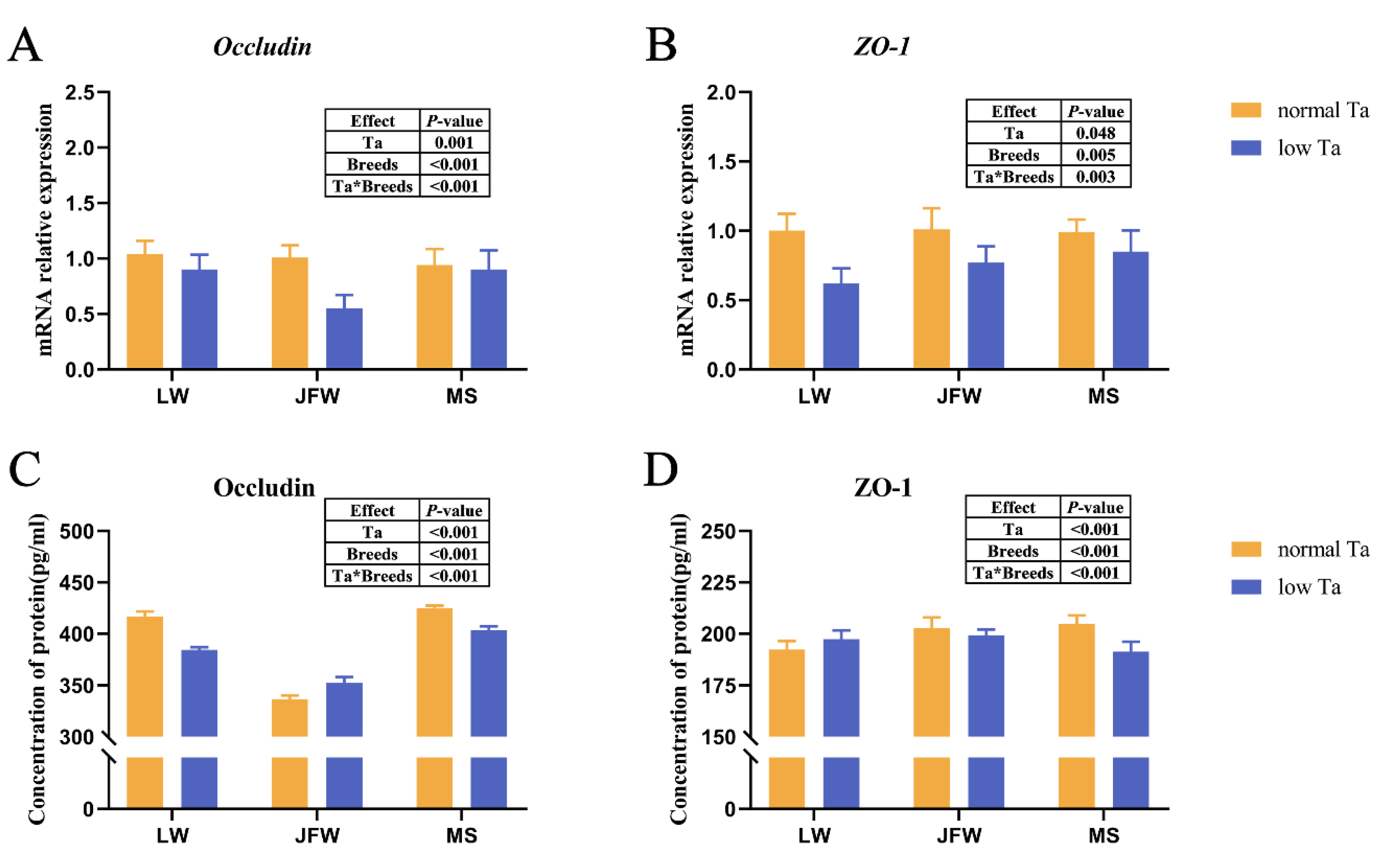
3.4. Tight Junction Expression
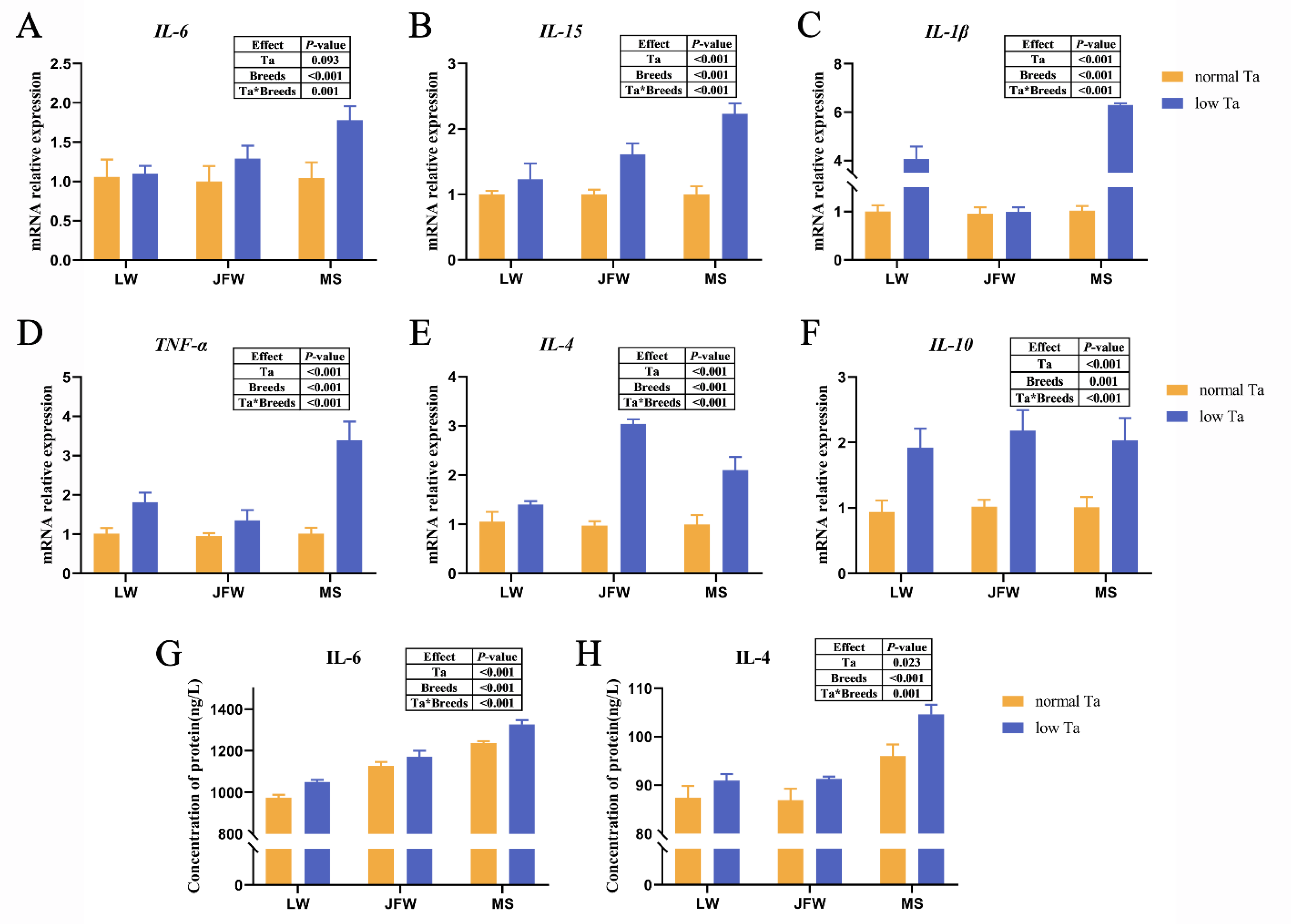
3.5. Inflammatory Cytokine Expression
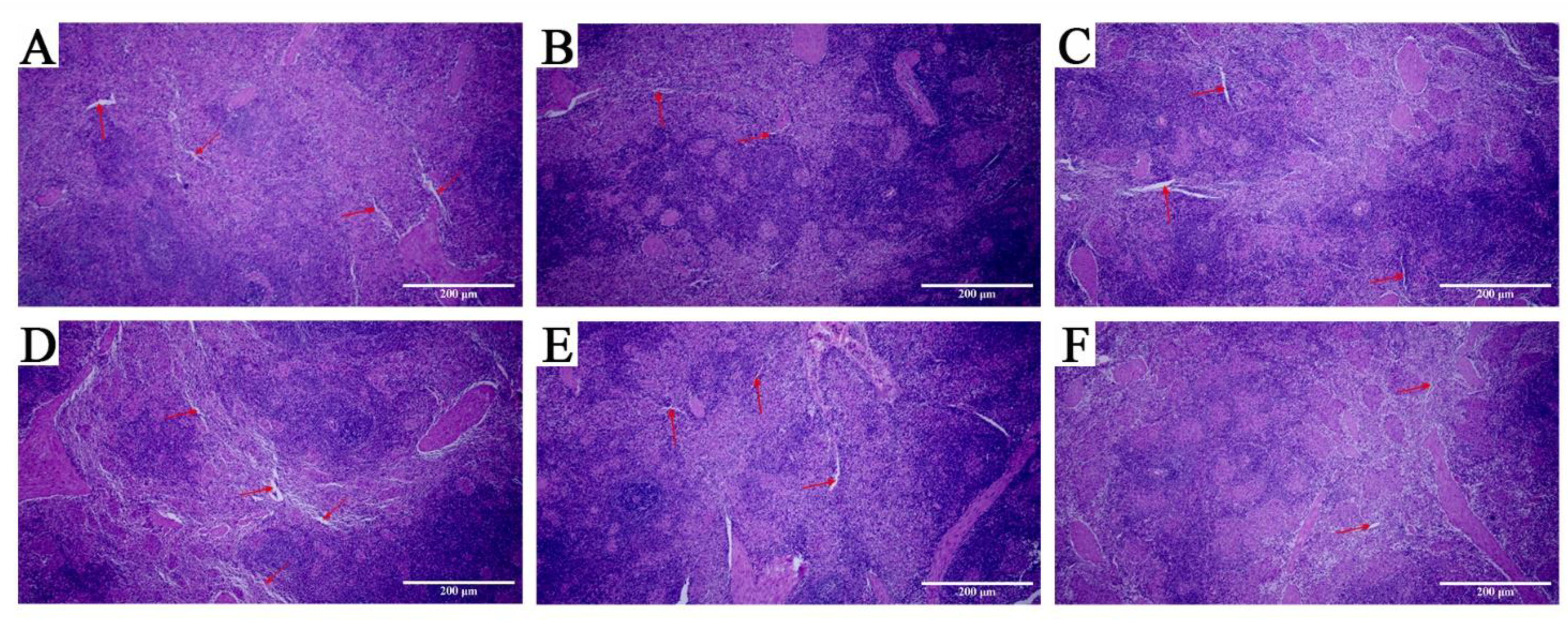
3.6. Pathological Observation of Spleen
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ganta, C.K.; Helwig, B.G.; Blecha, F.; Ganta, R.R.; Cober, R.; Parimi, S.; Musch, T.I.; Fels, R.J.; Kenney, M.J. Hypothermia-enhanced splenic cytokine gene expression is independent of the sympathetic nervous system. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 291, R558–R565. [Google Scholar] [CrossRef] [PubMed]
- Leppaluoto, J.; Westerlund, T.; Huttunen, P.; Oksa, J.; Smolander, J.; Dugue, B.; Mikkelsson, M. Effects of long-term whole-body cold exposures on plasma concentrations of ACTH, beta-endorphin, cortisol, catecholamines and cytokines in healthy females. Scand. J. Clin. Lab. Invest. 2008, 68, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Carroll, J.A.; Burdick, N.C.; Chase, C.C., Jr.; Coleman, S.W.; Spiers, D.E. Influence of environmental temperature on the physiological, endocrine, and immune responses in livestock exposed to a provocative immune challenge. Domest. Anim. Endocrinol. 2012, 43, 146–153. [Google Scholar] [CrossRef]
- Wei, H.; Zhang, R.; Su, Y.; Bi, Y.; Li, X.; Zhang, X.; Li, J.; Bao, J. Effects of Acute Cold Stress After Long-Term Cold Stimulation on Antioxidant Status, Heat Shock Proteins, Inflammation and Immune Cytokines in Broiler Heart. Front. Physiol. 2018, 9, 1589. [Google Scholar] [CrossRef]
- Nadler, S.F.; Weingand, K.; Kruse, R.J. The physiologic basis and clinical applications of cryotherapy and thermotherapy for the pain practitioner. Pain Physician 2004, 7, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Marino, F.; Sockler, J.M.; Fry, J.M. Thermoregulatory, metabolic and sympathoadrenal responses to repeated brief exposure to cold. Scand. J. Clin. Lab. Invest. 1998, 58, 537–545. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Liu, Y.; Li, C.; Zhang, R.; Bao, J. Effects of Intermittent Mild Cold Stimulation on mRNA Expression of Immunoglobulins, Cytokines, and Toll-Like Receptors in the Small Intestine of Broilers. Animals 2020, 10, 1492. [Google Scholar] [CrossRef]
- Fan, F.; Li, L.; Liu, W.; Yang, M.; Ma, X.; Sun, H. Astrocytes and neurons in locus coeruleus mediate restraint water immersion stress-induced gastric mucosal damage through the ERK1/2 signaling pathway. Neurosci. Lett. 2018, 675, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, L.; Zhu, R.-Z.; Zhang, S.; Liu, S.; Wang, Y.; Wu, Y.; Xing, S.; Liao, X.; Mi, J. Cold Stress Activates the UCP3 Pathway via Gut Microbiota in Piglet. 2021. Available online: https://www.researchgate.net/publication/350882667_Cold_Stress_Activates_the_UCP3_Pathway_via_Gut_Microbiota_in_Piglet (accessed on 3 April 2021).
- Zhou, H.J.; Kong, L.L.; Zhu, L.X.; Hu, X.Y.; Busye, J.; Song, Z.G. Effects of cold stress on growth performance, serum biochemistry, intestinal barrier molecules, and adenosine monophosphate-activated protein kinase in broilers. Animal 2021, 15, 100138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, S.; Liu, Y.; Ma, J.; Li, W.; Xu, X.; Wang, Y.; Luo, Y.; Cheng, K.; Zhuang, R. Acute Cold Water-Immersion Restraint Stress Induces Intestinal Injury and Reduces the Diversity of Gut Microbiota in Mice. Front. Cell Infect. Microbiol. 2021, 11, 706849. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, C.; Stojanovic, O.; Colin, D.J.; Suarez-Zamorano, N.; Tarallo, V.; Veyrat-Durebex, C.; Rigo, D.; Fabbiano, S.; Stevanovic, A.; Hagemann, S.; et al. Gut Microbiota Orchestrates Energy Homeostasis during Cold. Cell 2015, 163, 1360–1374. [Google Scholar] [CrossRef]
- Bo, T.B.; Zhang, X.Y.; Wen, J.; Deng, K.; Qin, X.W.; Wang, D.H. The microbiota-gut-brain interaction in regulating host metabolic adaptation to cold in male Brandt’s voles (Lasiopodomys brandtii). ISME J. 2019, 13, 3037–3053. [Google Scholar] [CrossRef]
- Xu, Y.; Jia, J.; Xie, C.; Wu, Y.; Tu, W. Transient Receptor Potential Ankyrin 1 and Substance P Mediate the Development of Gastric Mucosal Lesions in a Water Immersion Restraint Stress Rat Model. Digestion 2018, 97, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Chaudhry, M.T.; Zhao, D.; Lin, T.; Tian, Y.; Fu, J. Heat shock protein 70 protects the quail cecum against oxidant stress, inflammatory injury, and microbiota imbalance induced by cold stress. Poult. Sci. 2019, 98, 5432–5445. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Lin, J.; Huang, J.; Zhang, H.; Zhang, R.; Zhang, X.; Cao, C.; Hambly, C.; Qin, G.; Yao, J.; et al. Reconstitution of UCP1 using CRISPR/Cas9 in the white adipose tissue of pigs decreases fat deposition and improves thermogenic capacity. Proc. Natl. Acad. Sci. USA 2017, 114, E9474–E9482. [Google Scholar] [CrossRef]
- Teleglow, A.; Romanovski, V.; Skowron, B.; Mucha, D.; Tota, L.; Rosinczuk, J.; Mucha, D. The Effect of Extreme Cold on Complete Blood Count and Biochemical Indicators: A Case Study. Int. J. Environ. Res. Public Health 2021, 19, 424. [Google Scholar] [CrossRef]
- Ma, H.; Fu, B.; Zhang, X.; Wang, L.; Li, Z.; Liu, D. Expression and subcellular localization of HSPC117 in min pig tissues and the PK15 cell line. Technol. Health Care 2019, 27, 301–306. [Google Scholar] [CrossRef]
- Li, M.; Zhang, N.; Zhang, W.; Hei, W.; Cai, C.; Yang, Y.; Lu, C.; Gao, P.; Guo, X.; Cao, G.; et al. Comprehensive analysis of differentially expressed circRNAs and ceRNA regulatory network in porcine skeletal muscle. BMC Genom. 2021, 22, 320. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Cheng, Z.; Li, M.; Zhang, N.; Le, B.; Zhang, W.; Song, P.; Guo, X.; Li, B.; Cao, G. Selection of candidate genes affecting meat quality and preliminary exploration of related molecular mechanisms in the Mashen pig. Asian-Australas J. Anim. Sci. 2019, 32, 1084–1094. [Google Scholar] [CrossRef]
- Cai, C.; Li, M.; Zhang, Y.; Meng, S.; Yang, Y.; Gao, P.; Guo, X.; Cao, G.; Li, B. Comparative Transcriptome Analyses of Longissimus thoracis Between Pig Breeds Differing in Muscle Characteristics. Front. Genet. 2020, 11, 526309. [Google Scholar] [CrossRef]
- Lu, C.; Liu, Y.; Ma, Y.; Wang, S.; Cai, C.; Yang, Y.; Zhao, Y.; Liang, G.; Cao, G.; Li, B.; et al. Comparative Evaluation of the Ileum Microbiota Composition in Piglets at Different Growth Stages. Front. Microbiol. 2021, 12, 765691. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, Y.; Liu, J.; Wang, H.; Guo, Y.; Du, M.; Cai, C.; Zhao, Y.; Lu, C.; Guo, X.; et al. Composition of the Fecal Microbiota of Piglets at Various Growth Stages. Front. Vet. Sci. 2021, 8, 661671. [Google Scholar] [CrossRef] [PubMed]
- Kisielinski, K.; Willis, S.; Prescher, A.; Klosterhalfen, B.; Schumpelick, V. A simple new method to calculate small intestine absorptive surface in the rat. Clin. Exp. Med. 2002, 2, 131–135. [Google Scholar] [CrossRef] [PubMed]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.; Moorman, A.F. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Knechtle, B.; Waskiewicz, Z.; Sousa, C.V.; Hill, L.; Nikolaidis, P.T. Cold Water Swimming-Benefits and Risks: A Narrative Review. Int. J. Environ. Res. Public Health 2020, 17, 8984. [Google Scholar] [CrossRef] [PubMed]
- Glaser, R.; Kiecolt-Glaser, J.K. Stress-induced immune dysfunction: Implications for health. Nat. Rev. Immunol. 2005, 5, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Poller, W.C.; Downey, J.; Mooslechner, A.A.; Khan, N.; Li, L.; Chan, C.T.; McAlpine, C.S.; Xu, C.; Kahles, F.; He, S.; et al. Brain motor and fear circuits regulate leukocytes during acute stress. Nature 2022, 607, 578–584. [Google Scholar] [CrossRef]
- Bischoff, S.C. ‘Gut health’: A new objective in medicine? BMC Med. 2011, 9, 24. [Google Scholar] [CrossRef]
- Chung, H.; Kasper, D.L. Microbiota-stimulated immune mechanisms to maintain gut homeostasis. Curr. Opin. Immunol. 2010, 22, 455–460. [Google Scholar] [CrossRef]
- Fasano, A. Zonulin and its regulation of intestinal barrier function: The biological door to inflammation, autoimmunity, and cancer. Physiol. Rev. 2011, 91, 151–175. [Google Scholar] [CrossRef]
- Sharma, R.; Young, C.; Neu, J. Molecular Modulation of Intestinal Epithelial Barrier: Contribution of Microbiota. J. Biomed. Biotechnol. 2010, 2010, 305879. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.B.; Ravindran, V.; Thomas, D.G.; Birtles, M.J.; Hendriks, W.H. Influence of method of whole wheat inclusion and xylanase supplementation on the performance, apparent metabolisable energy, digestive tract measurements and gut morphology of broilers. Br. Poult. Sci. 2004, 45, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.Q.; Zhang, Z.W.; Yao, H.D.; Wang, L.L.; Liu, T.; Yu, X.Y.; Li, S.; Xu, S.W. Effects of cold stress on mRNA expression of immunoglobulin and cytokine in the small intestine of broilers. Res. Vet. Sci. 2013, 95, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xie, Y.; Zhong, R.; Liu, L.; Lin, C.; Xiao, L.; Chen, L.; Zhang, H.; Beckers, Y.; Everaert, N. Effects of Xylo-Oligosaccharides on Growth and Gut Microbiota as Potential Replacements for Antibiotic in Weaning Piglets. Front. Microbiol. 2021, 12, 641172. [Google Scholar] [CrossRef] [PubMed]
- Krug, S.M.; Schulzke, J.D.; Fromm, M. Tight junction, selective permeability, and related diseases. Semin Cell Dev. Biol. 2014, 36, 166–176. [Google Scholar] [CrossRef]
- He, W.; Gao, Y.; Guo, Z.; Yang, Z.; Wang, X.; Liu, H.; Sun, H.; Shi, B. Effects of fermented wheat bran and yeast culture on growth performance, immunity, and intestinal microflora in growing-finishing pigs. J. Anim. Sci. 2021, 99, skab308. [Google Scholar] [CrossRef]
- Dodd, D.; Spitzer, M.H.; Van Treuren, W.; Merrill, B.D.; Hryckowian, A.J.; Higginbottom, S.K.; Le, A.; Cowan, T.M.; Nolan, G.P.; Fischbach, M.A.; et al. A gut bacterial pathway metabolizes aromatic amino acids into nine circulating metabolites. Nature 2017, 551, 648–652. [Google Scholar] [CrossRef]
- Thaiss, C.A.; Levy, M.; Grosheva, I.; Zheng, D.; Soffer, E.; Blacher, E.; Braverman, S.; Tengeler, A.C.; Barak, O.; Elazar, M.; et al. Hyperglycemia drives intestinal barrier dysfunction and risk for enteric infection. Science 2018, 359, 1376–1383. [Google Scholar] [CrossRef]
- Neurath, M.F. Cytokines in inflammatory bowel disease. Nat. Rev. Immunol. 2014, 14, 329–342. [Google Scholar] [CrossRef]
- Liu, T.; Guo, Y.; Lu, C.; Cai, C.; Gao, P.; Cao, G.; Li, B.; Guo, X.; Yang, Y. Effect of Different Pig Fecal Microbiota Transplantation on Mice Intestinal Function and Microbiota Changes During Cold Exposure. Front. Vet. Sci. 2022, 9, 805815. [Google Scholar] [CrossRef]
- Kim, M.H.; Kang, S.G.; Park, J.H.; Yanagisawa, M.; Kim, C.H. Short-chain fatty acids activate GPR41 and GPR43 on intestinal epithelial cells to promote inflammatory responses in mice. Gastroenterology 2013, 145, 396–406.e10. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences (5′ -> 3′) | Product Length (bp) | GenBank Accession No. |
|---|---|---|---|
| Occludin | F: CGAGACAGACTACACGACGG R: TTCATCAGCAGCAGCCATGT | 247 | NM_001163647.2 |
| ZO-1 1 | F: AGCCCGAGGCGTGTTT R: GGTGGGAGGATGCTGTTG | 147 | XM_021098891.1 |
| IL-4 2 | F: TCACCTCCCAACTGATCCCA R: GCTCCATGCACGAGTTCTTT | 144 | NM_214123.1 |
| IL-10 3 | F: CCACAAGTCCGACTCAACGA R: GGCAACCCAGGTAACCCTTA | 267 | NM_214041.1 |
| IL-6 4 | F: AGACCCTGAGGCAAAAGGGAAA R: CGGCATCAATCTCAGGTGCC | 209 | NM_214399.1 |
| IL-15 5 | F: TGCATCCAGTGCTACTTGTGT R: CCTGCACTGATACAGCCCAA | 92 | NM_214390.1 |
| IL-1β 6 | F: CCAATTCAGGGACCCTACCC R: GTTTTGGGTGCAGCACTTCAT | 174 | NM_214055.1 |
| TNF-α 7 | F: TGCACTTCGAGGTTATCGGC R: CGGCTTTGACATTGGCTACAA | 141 | NM_214022.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Liu, T.; Li, W.; Zhang, W.; Cai, C.; Lu, C.; Gao, P.; Cao, G.; Li, B.; Guo, X.; et al. Effects of Low-Ambient-Temperature Stimulation on Modifying the Intestinal Structure and Function of Different Pig Breeds. Animals 2022, 12, 2740. https://doi.org/10.3390/ani12202740
Guo Y, Liu T, Li W, Zhang W, Cai C, Lu C, Gao P, Cao G, Li B, Guo X, et al. Effects of Low-Ambient-Temperature Stimulation on Modifying the Intestinal Structure and Function of Different Pig Breeds. Animals. 2022; 12(20):2740. https://doi.org/10.3390/ani12202740
Chicago/Turabian StyleGuo, Yanbo, Ting Liu, Wenxia Li, Wanfeng Zhang, Chunbo Cai, Chang Lu, Pengfei Gao, Guoqing Cao, Bugao Li, Xiaohong Guo, and et al. 2022. "Effects of Low-Ambient-Temperature Stimulation on Modifying the Intestinal Structure and Function of Different Pig Breeds" Animals 12, no. 20: 2740. https://doi.org/10.3390/ani12202740
APA StyleGuo, Y., Liu, T., Li, W., Zhang, W., Cai, C., Lu, C., Gao, P., Cao, G., Li, B., Guo, X., & Yang, Y. (2022). Effects of Low-Ambient-Temperature Stimulation on Modifying the Intestinal Structure and Function of Different Pig Breeds. Animals, 12(20), 2740. https://doi.org/10.3390/ani12202740