


Chromosomal Diversification in Pseudacanthicus Species (Loricariidae, Hypostominae) Revealed by Comparative Mapping of Repetitive Sequences

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Chromosomal Analysis
2.3. Probes Labeling and Fluorescence In Situ Hybridization
2.4. Image Capture and Analysis
3. Results
3.1. Classical Cytogenetics
3.2. Molecular Cytogenetics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. (Eds.) Eschmeyer’s Catalog of Fishes: Genera, Species, References. 2021. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 21 April 2021).
- Armbruster, J.W. Phylogenetic relationships of the suckermouth armoured catfishes Loricariidae with emphasis on the Hypostominae and the Ancistrinae. Zool. J. Linn. Soc. 2004, 141, 1–80. [Google Scholar] [CrossRef]
- Armbruster, J.W. The genus Peckoltia with the description of two new species and a reanalysis of the phylogeny of the genera of the Hypostominae (Siluriformes: Loricariidae). Zootaxa 2008, 1822, 1–76. [Google Scholar] [CrossRef]
- Lujan, N.K.; Armbruster, J.W.; Lovejoy, N.R.; López-Fernández, H. Multilocus molecular phylogeny of the suckermouth armored catfishes (Siluriformes: Loricariidae) with a focus on subfamily Hypostominae. Mol. Phylogenetics Evol. 2015, 82, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Chamon, C.C. Pseudacanthicus pitanga: A new species of Ancistrini (Siluriformes: Loricariidae: Hypostominae) from rio Tocantins Basin, North Brazil. Zootaxa 2015, 3973, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Chamon, C.C.; Silva, T.C. Pseudacanthicus major: Description of one of the largest known Loricariidae (Hypostominae: Ancistrini), a species from rio Tocantins basin, Brazil. Zootaxa 2018, 4387, 499–510. [Google Scholar] [CrossRef]
- Chamon, C.C.; Sousa, L.M. A new species of the leopard pleco genus Pseudacanthicus (Siluriformes: Loricariidae) from the Rio Xingu, Brazil. J. Fish Biol. 2017, 90, 356–369. [Google Scholar] [CrossRef]
- Milhomem, S.S.; Pieczarka, J.C.; Crampton, W.G.; Silva, D.S.; De Souza, A.C.P.; Carvalho, J.R.; Nagamachi, C.Y. Chromosomal evidence for a putative cryptic species in the Gymnotus carapo species-complex (Gymnotiformes, Gymnotidae). BMC Genet. 2008, 75, 1–10. [Google Scholar] [CrossRef]
- Pety, A.M.; Cardoso, A.L.; Nagamachi, C.Y.; Pieczarka, J.C.; de Sousa, L.M.; Noronha, R.C.R. In situ localization of ribosomal sites in Peckoltia and Ancistomus (Loricariidae: Hypostominae) from the Amazon Basin. Zebrafish 2018, 15, 263–269. [Google Scholar] [CrossRef]
- da Silva, K.S.; de Souza, A.C.P.; Pety, A.M.; Noronha, R.C.R.; Vicari, M.R.; Pieczarka, J.C.; Nagamachi, C.Y. Comparative cytogenetics analysis among Peckoltia species (Siluriformes, Loricariidae): Insights on karyotype evolution and biogeography in the amazon region. Front. Genet. 2021, 12, 779464. [Google Scholar] [CrossRef]
- Kavalco, K.F.; Pazza, R.; Bertollo, L.A.; Moreira-Filho, O. Karyotypic diversity and evolution of Loricariidae (Pisces, Siluriformes). Heredity 2005, 94, 180–186. [Google Scholar] [CrossRef]
- Glugoski, L.; Deon, G.; Schott, S.; Vicari, M.R.; Nogaroto, V.; Moreira-Filho, O. Comparative cytogenetic analyses in Ancistrus species (Siluriformes: Loricariidae). Neotrop. Ichthyol. 2020, 18. [Google Scholar] [CrossRef]
- Bueno, V.; Konerat, J.T.; Zawadzki, C.H.; Venere, P.C.; Blanco, D.R.; Margarido, V.P. Divergent Chromosome Evolution in Hypostominae Tribes (Siluriformes: Loricariidae): Correlation of Chromosomal Data with Morphological and Molecular Phylogenies. Zebrafish 2018, 15, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Ziemniczak, K.; Barros, A.V.; Rosa, K.O.; Nogaroto, V.; Almeida, M.C.; Cestari, M.M.; Moreira-Filho, O.; Artoni, R.F.; Vicari, M.R. Comparative cytogenetics of Loricariidae (Actinopterygii: Siluriformes): Emphasis in Neoplecostominae and Hypoptopomatinae. Ital. J. Zool. 2012, 79, 492–501. [Google Scholar] [CrossRef]
- Eickbush, T.H.; Eickbush, D.G. Finely orchestrated movements: Evolution of the ribosomal RNA genes. Genetics 2007, 175, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Long, E.O.; Dawid, I.B. Repeated genes in eukaryotes. Annu. Rev. Biochem. 1980, 49, 727–764. [Google Scholar] [CrossRef] [PubMed]
- Busch, H.; Reddy, R.; Rothblum, L.; Choi, Y.C. SnRNAs, SnRNPs, and RNA processing. Annu. Rev. Biochem. 1982, 51, 617–654. [Google Scholar] [CrossRef]
- Cabral-de-Mello, D.C.; Valente, G.T.; Nakajima, R.T.; Martins, C. Genomic organization and comparative chromosome mapping of the U1 snRNA gene in cichlid fish, with an emphasis in Oreochromis niloticus. Chromosome Res. 2012, 20, 279–292. [Google Scholar] [CrossRef]
- Silva, D.M.; Utsunomia, R.; Pansonato-Alves, J.C.; Oliveira, C.; Foresti, F. Chromosomal mapping of repetitive DNA sequences in five species of Astyanax (Characiformes, Characidae) reveals independent location of U1 and U2 snRNA sites and association of U1 snRNA and 5S rDNA. Cytogenet. Genome Res. 2015, 146, 144–152. [Google Scholar] [CrossRef]
- Barros, A.V.; Wolski, M.A.V.; Nogaroto, V.; Almeida, M.C.; Moreira-Filho, O.; Vicari, M.R. Fragile sites, dysfunctional telomere and chromosome fusions: What is 5S rDNA role? Gene 2017, 608, 20–27. [Google Scholar] [CrossRef]
- Yano, C.F.; Bertollo, L.A.C.; Rebordinos, L.; Merlo, M.A.; Liehr, T.; Portela-Bens, S.; Cioffi, M.B. Evolutionary dynamics of rDNAs and U2 small nuclear DNAs in Triportheus (Characiformes, Triportheidae): High variability and particular syntenic organization. Zebrafish 2017, 14, 146–154. [Google Scholar] [CrossRef]
- Ditcharoen, S.; Bertollo, A.C.L.; Ráb, P.; Hnátková, E.; Franco Molina, W.; Liehr, T.; Tanomtong, A.; Triantaphyllidis, C.; Ozouf-Costa, C.; Yongnunui, S.; et al. Genomic organization of repetitive DNA elements and extensive karyotype diversity of silurid catfishes (Teleostei: Siluriformes): A comparative cytogenetic approach. Int. J. Mol. Sci. 2019, 20, 3545. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, D.S.; da Silva Filho, H.F.; Cioffi, M.B.; de Oliveira, E.H.; Gomes, A.J.B. Comparative cytogenetics in four Leptodactylus species (Amphibia, Anura, Leptodactylidae): Evidence of inner chromosomal diversification in highly conserved karyotypes. Cytogenet. Genome Res. 2021, 161, 52–62. [Google Scholar] [CrossRef]
- Bertollo, L.A.C.; Takahashi, C.S.; Moreira-Filho, O. Cytotaxonomic considerations on Hoplias lacerdae (Pisces Erythrinidae). Braz. J. Genet. 1978, 1, 103–120. [Google Scholar]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Martins, C.; Galetti Jr, P.M. Chromosomal localization of 5S rDNA genes in Leporinus Fish (Anostomidae, Characiformes). Chromosome Res. 1999, 7, 363–367. [Google Scholar] [CrossRef]
- Suarez, P.; Barroso, I.C.G.P.; Silva, D.S.; Milhomem, S.S.R.; Cabral-de-Mello, D.C.; Martins, C.; Pieczarka, J.C.; Nagamachi, C.Y. Highest diploid number among Gymnotiformes: First cytogenetic insights into Rhandolichops (Sternopygidae). Zebrafish 2017, 14, 272–279. [Google Scholar] [CrossRef]
- Colgan, D.J.; McLauchlan, A.; Wilson, G.D.F.; Livingston, S.P.; Edgecombe, G.D.; Macaranas, J.; Cassi, G.; Gray, M.R. Histone H3 and U2 snRNA DNA sequences and arthropod molecular evolution. Aust. J. Zool. 1998, 46, 419–437. [Google Scholar] [CrossRef]
- Hatanaka, T.; Galetti, P.M., Jr. Mapping of 18S and 5S ribosomal RNA genes in the fish Prochilodus argenteus Agassiz, 1829 (Characiformes, Prochilodontidae). Genetica 2004, 122, 239–244. [Google Scholar] [CrossRef]
- Ijdo, J.W.; Wells, R.A.; Baldini, A.; Reeders, S.T. Improved telomere detection using a telomere repeat probe (TTAGGG)n generated by PCR. Nucleic Acids Res. 1991, 19, 4780. [Google Scholar]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Artoni, R.F.; Bertollo, L.A.C. Trends in the karyotype evolution of Loricariidae fish (Siluriformes). Hereditas 2001, 134, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Santos da Silva, K.; Glugoski, L.; Vicari, M.R.; de Souza, A.C.P.; Noronha, R.C.R.; Pieczarka, J.C.; Nagamachi, C.Y. Chromosomal Diversification in Ancistrus Species (Siluriformes: Loricariidae) Inferred From Repetitive Sequence Analysis. Front. Genet. 2022, 13, 838462. [Google Scholar] [CrossRef]
- Cereali, S.S.; Pomini, E.; Rosa, R.; Zawadzki, C.H.; Froehlich, O.; Giuliano-Caetano, L. Karyotype description of two species of Hypostomus (Siluriformes, Loricariidae) of the Planalto da Bodoquena, Brazil. Genet. Mol. Res. 2008, 7, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Bueno, V.; Venere, P.C.; Zawadzki, C.H.; Margarido, V.P. Karyotypic diversification in Hypostomus Lacepède, 1803 (Siluriformes, Loricariidae): Biogeographical and phylogenetic perspectives. Rev. Fish Biol. Fish. 2013, 23, 103–112. [Google Scholar] [CrossRef]
- Da Silva, M.; Ribeiro, E.D.; Matoso, D.A.; Sousa, L.M.; Hrbek, T.; Py-Daniel, L.R.; Feldberg, E. Chromosomal polymorphism in two species of Hypancistrus (Siluriformes: Loricariidae): An integrative approach for understanding their biodiversity. Genetica 2014, 142, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Tchurikov, N.A.; Fedoseeva, D.M.; Sosin, D.V.; Snezhkina, A.V.; Melnikova, N.V.; Kudryavtseva, A.V.; Kravatsky, Y.V.; Kretova, O.V. Hot spots of DNA double-strand breaks and genomic contacts of human rDNA units are involved in epigenetic regulation. J. Mol. Cell Biol. 2015, 7, 366–382. [Google Scholar] [CrossRef]
- Cazaux, B.; Catalan, J.; Veyrunes, F.; Douzery, E.J.; Britton-Davidian, J. Are ribosomal DNA clusters rearrangement hotspots? A case study in the genus Mus (Rodentia, Muridae). BMC Evol. Biol. 2011, 11, 1–14. [Google Scholar] [CrossRef]
- Gornung, E. Twenty years of physical mapping of major ribosomal RNA genes across the teleosts: A review of research. Cytogenet. Genome Res. 2013, 141, 90–102. [Google Scholar] [CrossRef]
- Rebordinos, L.; Cross, I.; Merlo, A. High evolutionary dynamism in 5S rDNA of fish: State of the art. Cytogenet. Genome Res. 2013, 141, 103–113. [Google Scholar] [CrossRef]
- Kidwell, M.G.; Lisch, D.R. Transposable elements and host genome evolution. Tree 2000, 15, 3.0169–5347. [Google Scholar] [CrossRef]
- Dimitri, P.; Caizzi, R.; Giordano, E.; Accardo, M.C.; Lattanzi, G.; Biamonti, G. Constitutive heterochromatin: A surprising variety of expressed sequences. Chromosoma 2009, 118, 419–435. [Google Scholar] [CrossRef] [PubMed]
- Cioffi, M.B.; Martins, C.; Bertollo, L.A.C. Chromosome spreading of associated transposable elements and ribosomal DNA in the fish Erythrinus erythrinus. Implications for genome change and karyoevolution in fish. BMC Evol. Biol. 2010, 10, 271. [Google Scholar] [CrossRef]
- Vicari, M.R.; Artoni, R.F.; Bertollo, L.A.C. Heterochromatin polymorphism associated with 18S rDNA: A differential pathway among Hoplias malabaricus fish populations. Cytogenet. Genome Res. 2003, 101, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Ma, L.; Yang, F.; Fei, S.Z.; Li, L. 45S rDNA regions are chromosome fragile sites expressed as gaps in vitro on metaphase chromosomes of root-tip meristematic cells in Lolium spp. PLoS ONE 2008, 3, e2167. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, B.R.R.; Milhomem-Paixão, S.S.R.; Noronha, R.C.R.; Nagamachi, C.Y.; Da Costa, M.J.R.; de Oliveira Pardal, P.P.; Coelho, S.J.; Pieczarka, J.C. Karyotype diversity and chromosomal organization of repetitive DNA in Tityus obscurus (Scorpiones, Buthidae). BMC Genet. 2017, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Cavalcante, M.G.; Souza, L.F.; Vicari, M.R.; de Bastos, C.E.M.; de Sousa, J.V.; Nagamachi, C.Y.; Pieczarka, J.C.; Martins, C.; Noronha, R.C.R.N. Molecular cytogenetics characterization of Rhinoclemmys punctularia (Testudines, Geoemydidae) and description of a Gypsy-H3 association in its genome. Gene 2020, 738, 144477. [Google Scholar] [CrossRef]
- Ubeda-Manzanaro, M.; Merlo, M.A.; Palazon, J.L.; Cross, I.; Sarasquete, C.; Rebordinos, L. Chromosomal mapping of the major and minor ribosomal genes, (GATA)n and U2 snRNA gene by double-colour FISH in species of the Batrachoididae family. Genetica 2010, 138, 787–794. [Google Scholar] [CrossRef]
- Scacchetti, P.C.; Utsunomia, R.; Pansonato-Alves, J.C.; Silva, G.J.C.; Vicari, M.R.; Artoni, R.F.; Oliveira, C.; Foresti, F. Repetitive DNA Sequences and Evolution of ZZ/ZW Sex Chromosomes in Characidium (Teleostei: Characiformes). PLoS ONE 2015, 10, e0137231. [Google Scholar] [CrossRef]
- Piscor, D.; Centofante, L.; Parise-Maltempi, P.P. Highly similar morphologies between chromosomes bearing U2 snRNA gene clusters in the group Astyanax Baird and Girard, 1854 (Characiformes, Characidae): An evolutionary approach in species with 2n = 36, 46, 48, and 50. Zebrafish 2016, 13, 565–570. [Google Scholar] [CrossRef]
- Malimpensa, G.D.C.; Traldi, J.B.; Martinez, J.D.F.; Deon, G.; Azambuja, M.; Nogaroto, V.; Vicari, M.R.; Moreira-Filho, O. Chromosomal diversification in two species of Pimelodus (Siluriformes: Pimelodidae): Comparative cytogenetic mapping of multigene families. Zebrafish 2020, 17, 278–286. [Google Scholar] [CrossRef]
- Dulz, T.A.; Azambuja, M.; Nascimento, V.D.; Lorscheider, C.A.; Noleto, R.B.; Moreira-Filho, O.; Nogaroto, V.; Diniz, D.; Afonso, P.R.A.M.; Vicari, M.R. Karyotypic diversification in two Megaleporinus species (Characiformes, Anostomidae) inferred from in situ localization of repetitive DNA sequences. Zebrafish 2020, 17, 333–341. [Google Scholar] [PubMed]
- Azambuja, M.; Schemberger, M.O.; Nogaroto, V.; Moreira-Filho, O.; Martins, C.; Vicari, M.R. Major and minor U small nuclear RNAs genes characterization in a neotropical fish genome: Chromosomal remodeling and repeat units dispersion in Parodontidae. Gene 2022, 826, 146459. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | L-Number Code | Sex | Voucher * | Rivers | Hydrographic Basin |
|---|---|---|---|---|---|
| Pseudacanthicus spinosus | L-160 | 6♂/2♀ | P4212 | Tocantins river | Tocantins-Araguaia basin |
| Pseudacanthicus leopardus | L-114 | 1♂/1♀ | P4252 | Xingu river | Amazon basin |
| Pseudacanthicus sp. | L-185 | 1♂/1♀ | P4258 | Xingu river | Amazon basin |
| Species | Karyotypes | Molecular Cytogenetics | |||||
|---|---|---|---|---|---|---|---|
| 2n | NF | KF | rDNA 18S | rDNA 5S | snDNA U2 | Tel. | |
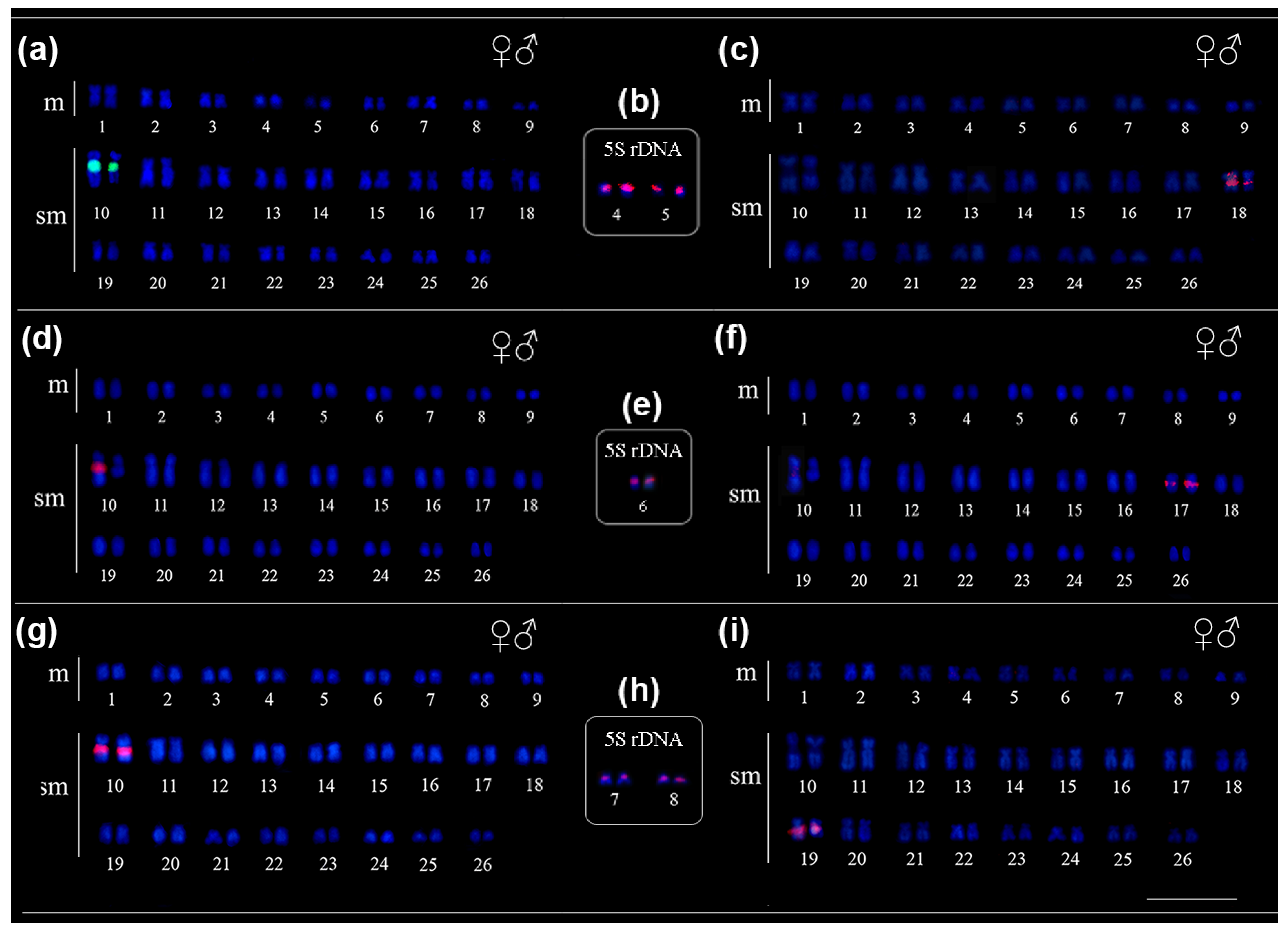
| Pseudacanthicus spinosus | 52 | 104 | 18 m + 34 sm | 10q | 4, 5 | 18q | distal |
| Pseudacanthicus leopardus | 52 | 104 | 18 m + 34 sm | homologue 10q | 6 | 17q | distal |
| Pseudacanthicussp. | 52 | 104 | 18 m + 34 sm | 10q | 7, 8 | 19q | distal |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos da Silva, K.; de Souza, A.C.P.; Rodrigues, L.R.R.; Pieczarka, J.C.; Nagamachi, C.Y. Chromosomal Diversification in Pseudacanthicus Species (Loricariidae, Hypostominae) Revealed by Comparative Mapping of Repetitive Sequences. Animals 2022, 12, 2612. https://doi.org/10.3390/ani12192612
Santos da Silva K, de Souza ACP, Rodrigues LRR, Pieczarka JC, Nagamachi CY. Chromosomal Diversification in Pseudacanthicus Species (Loricariidae, Hypostominae) Revealed by Comparative Mapping of Repetitive Sequences. Animals. 2022; 12(19):2612. https://doi.org/10.3390/ani12192612
Chicago/Turabian StyleSantos da Silva, Kevin, Augusto César Paes de Souza, Luís Reginaldo Ribeiro Rodrigues, Julio Cesar Pieczarka, and Cleusa Yoshiko Nagamachi. 2022. "Chromosomal Diversification in Pseudacanthicus Species (Loricariidae, Hypostominae) Revealed by Comparative Mapping of Repetitive Sequences" Animals 12, no. 19: 2612. https://doi.org/10.3390/ani12192612
APA StyleSantos da Silva, K., de Souza, A. C. P., Rodrigues, L. R. R., Pieczarka, J. C., & Nagamachi, C. Y. (2022). Chromosomal Diversification in Pseudacanthicus Species (Loricariidae, Hypostominae) Revealed by Comparative Mapping of Repetitive Sequences. Animals, 12(19), 2612. https://doi.org/10.3390/ani12192612