

Yet Another Non-Unique Human Behaviour: Leave-Taking in Wild Chacma Baboons (Papio ursinus)


Abstract
Simple Summary
Abstract
1. Introduction
1.1. Lack of Leave-Taking Study
1.2. Addressing Previous Limitations
1.3. Baboon Models for Leave Taking
2. Materials and Methods
2.1. Addressing Methodological Issues
2.1.1. Using a Considered Definition
2.1.2. Defining the Level of Study
2.1.3. Data Collection Methods
2.1.4. Determining Candidate Behaviours
2.1.5. Control Conditions
2.2. Analyses
2.3. Study Site, Subjects, and Protocol
2.3.1. Gorongosa National Park
2.3.2. Data Collection Protocol
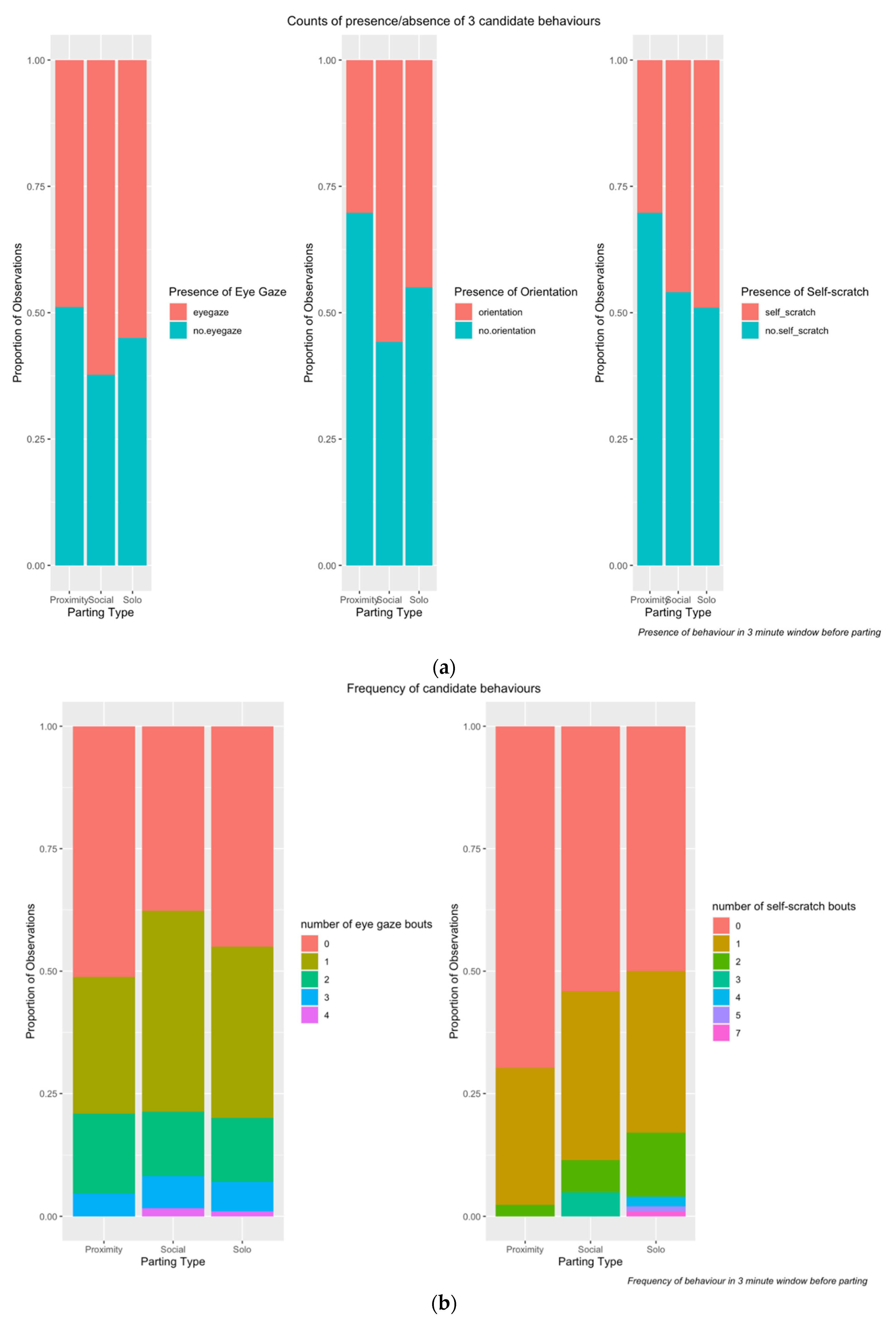
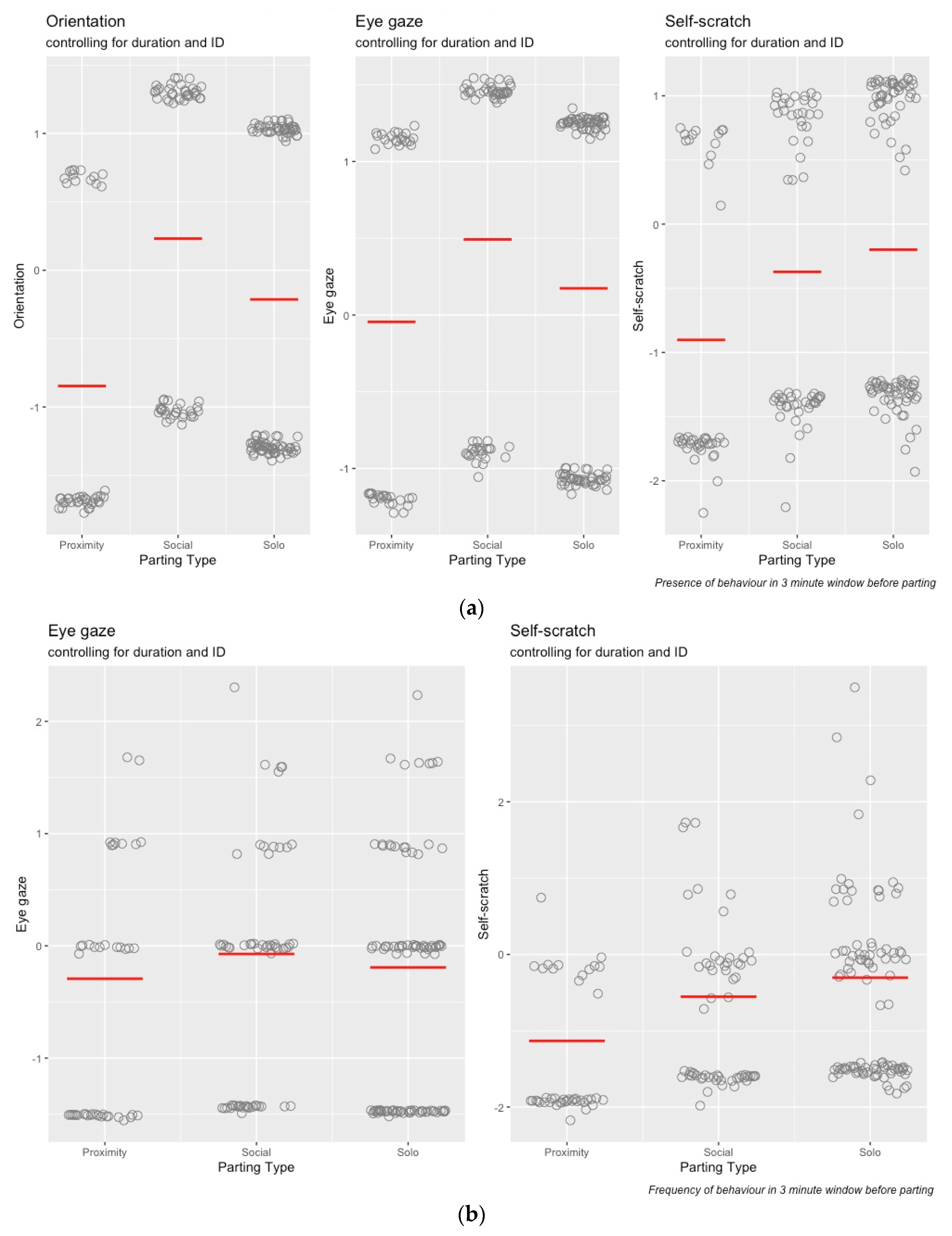
3. Results
3.1. IO Reliability
3.2. Statistical Analysis
4. Discussion
4.1. Evolutionary Context
4.2. Towards a New Framework for Leave Taking
4.2.1. Definition
4.2.2. Control Conditions
4.2.3. Levels of Separation
4.2.4. Data Collection Methods
4.2.5. Candidate Behaviours
4.2.6. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baehren, L. Saying “goodbye” to the conundrum of leave-taking: A cross-disciplinary review. Humanit. Soc. Sci. Commun. 2022, 9, 46. [Google Scholar] [CrossRef]
- Jolly, C.J. A proper study for mankind: Analogies from the papionin monkeys and their implications for human evolution. Am. J. Phys. Anthropol. 2001, 116, 177–204. [Google Scholar] [CrossRef] [PubMed]
- Nishida, T.; Kano, T.; Goodall, J.; Mcgrew, W.C.; Nakamura, M. Ethogram and ethnography of Mahale chimpanzees. Anthropol. Sci. 1999, 107, 141–188. [Google Scholar] [CrossRef]
- Scheumann, M.; Linn, S.; Zimmerman, E. Vocal greeting during mother-infant reunions in a nocturnal primate, the gray mouse lemur (Microcebus murinus). Sci. Rep. 2017, 7, 10321. [Google Scholar] [CrossRef]
- Goodall, J. The Chimpanzees of Gombe: Patterns of Behavior; Belknap Press: Cambridge, UK, 1986. [Google Scholar]
- Albert, S.; Kessler, S. Ending social encounters. J. Exp. Soc. Psychol. 1978, 14, 541–553. [Google Scholar] [CrossRef]
- Mcgrew, W.C.; Baehren, L. “Parting Is Such Sweet Sorrow”, But Only for Humans? Hum. Ethol. Bull. 2016, 31, 5–14. [Google Scholar] [CrossRef][Green Version]
- Van Lawick-Goodall, J. The behaviour of free-living chimpanzees in the Gombe Stream Reserve. Anim. Behav. Monogr. 1968, 1, IN1–IN311. [Google Scholar] [CrossRef]
- De Waal, F.; Waal, F.B. Chimpanzee Politics: Power and Sex among Apes; JHU Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Duranti, A. Universal and Culture-Specific Properties of Greetings. J. Linguist. Anthropol. 1997, 7, 63–97. [Google Scholar] [CrossRef]
- Lockard, J.S.; Allen, D.J.; Schiele, B.J.; Wiemer, M.J. Human postural signals: Stance, weight-shifts and social distance as intention movements to depart. Anim. Behav. 1978, 26, 219–224. [Google Scholar] [CrossRef]
- Kummer, H. Social Organization of Hamadryas Baboons; Karger Publishers: London, UK, 1968. [Google Scholar]
- Whittaker, S.; Frohlich, D.; Daly-Jones, O. Informal workplace communication: What is it like and how might we support it? In Proceedings of the SIGCHI Conference on Human Factors in Computing Systems, Boston, MA, USA, 24 April 1994; pp. 131–137. [Google Scholar]
- Wang, Y. A Pedagogy of Leave-Taking in Chinese. Doctoral Dissertation, The Ohio State University, Columbus, OH, USA, 2005. [Google Scholar]
- Gräfenhain, M.; Behne, T.; Carpenter, M.; Tomasello, M. Young children’s understanding of joint commitments. Dev. Psychol. 2009, 45, 1430. [Google Scholar] [CrossRef]
- Steiper, M.E.; Young, N.M. Primate molecular divergence dates. Mol. Phylogenet. Evol. 2006, 41, 384–394. [Google Scholar] [CrossRef]
- Langergraber, K.E.; Watts, D.P.; Vigilant, L.; Mitani, J.C. Group augmentation, collective action, and territorial boundary patrols by male chimpanzees. Proc. Natl. Acad. Sci. USA 2017, 114, 7337–7342. [Google Scholar] [CrossRef] [PubMed]
- Smuts, B.B.; Cheney, D.L.; Seyfarth, R.M.; Wrangham, R.W. Primate Societies; University of Chicago Press: Chicago, IL, USA, 2008. [Google Scholar]
- Grueter, C.C. Primate model offers insights into male bonding in complex societies. Proc. Natl. Acad. Sci. USA 2014, 111, 14645–14646. [Google Scholar] [CrossRef] [PubMed]
- Patzelt, A.; Kopp, G.H.; Ndao, I.; Kalbitzer, U.; Zinner, D.; Fischer, J. Male tolerance and male–male bonds in a multilevel primate society. Proc. Natl. Acad. Sci. USA 2014, 111, 14740–14745. [Google Scholar] [CrossRef] [PubMed]
- Silk, J.B.; Beehner, J.C.; Bergman, T.J.; Crockford, C.; Engh, A.L.; Moscovice, L.R.; Wittig, R.M.; Seyfarth, R.M.; Cheney, D.L. Strong and consistent social bonds enhance the longevity of female baboons. Curr. Biol. 2010, 20, 1359–1361. [Google Scholar] [CrossRef] [PubMed]
- Silk, J.B.; Alberts, S.C.; Altmann, J. Social bonds of female baboons enhance infant survival. Science 2003, 302, 1231–1234. [Google Scholar] [CrossRef]
- Cheney, D.L.; Seyfarth, R.M. Flexible usage and social function in primate vocalizations. Proc. Natl. Acad. Sci. USA 2018, 115, 1974–1979. [Google Scholar] [CrossRef]
- Goffe, A.S.; Zinner, D.; Fischer, J. Sex and friendship in a multilevel society: Behavioural patterns and associations between female and male Guinea baboons. Behav. Ecol. Sociobiol. 2016, 70, 323–336. [Google Scholar] [CrossRef]
- Buchan, J.C.; Alberts, S.C.; Silk, J.B.; Altmann, J. True paternal care in a multi-male primate society. Nature 2003, 425, 179–181. [Google Scholar] [CrossRef] [PubMed]
- Huchard, E.; Alvergne, A.; Féjan, D.; Knapp, L.A.; Cowlishaw, G.; Raymond, M. More than friends? Behavioural and genetic aspects of heterosexual associations in wild chacma baboons. Behav. Ecol. Sociobiol. 2010, 64, 769–781. [Google Scholar] [CrossRef]
- Lemasson, A.; Palombit, R.A.; Jubin, R. Friendships between males and lactating females in a free-ranging group of olive baboons (Papio hamadryas anubis): Evidence from playback experiments. Behav. Ecol. Sociobiol. 2008, 62, 1027–1035. [Google Scholar] [CrossRef]
- Nguyen, N.; Van Horn, R.C.; Alberts, S.C.; Altmann, J. “Friendships” between new mothers and adult males: Adaptive benefits and determinants in wild baboons (Papio cynocephalus). Behav. Ecol. Sociobiol. 2009, 63, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Fagot, J.; Boë, L.; Berthomier, F.; Claidière, N.; Malassis, R.; Meguerditchian, A.; Rey, A.; Montant, M. The baboon: A model for the study of language evolution. J. Hum. Evol. 2019, 126, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Chomsky, N.; Keyser, S.J. Language and Problems of Knowledge: The Managua Lectures; MIT Press: Cambridge, MA, USA, 1988; Volume 16. [Google Scholar]
- Stueckle, S.; Zinner, D. To follow or not to follow: Decision making and leadership during the morning departure in chacma baboons. Anim. Behav. 2008, 75, 1995–2004. [Google Scholar] [CrossRef]
- Strandburg-Peshkin, A.; Farine, D.R.; Couzin, I.D.; Crofoot, M.C. Shared decision-making drives collective movement in wild baboons. Science 2015, 348, 1358–1361. [Google Scholar] [CrossRef] [PubMed]
- Megeurditchian, A. On the gestural origins of language: What baboons’ gestures and brain have told us after 15 years of research. Ethol. Ecol. Evol. 2022, 34, 288–302. [Google Scholar] [CrossRef]
- Friard, O.; Gamba, M. BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol. Evol. 2016, 7, 1325–1330. [Google Scholar] [CrossRef]
- Knapp, M.L.; Hart, R.P.; Friedrich, G.W.; Shulman, G.M. The rhetoric of goodbye: Verbal and nonverbal correlates of human leave-taking. Commun. Monogr. 1973, 40, 182–198. [Google Scholar] [CrossRef]
- Kendon, A. Spatial organization in social encounters: The F-formation system. In Conducting Interaction: Patterns of Behavior in Focused Encounters; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Murray, L. Sensitivities and Expressive Capacities of Young Infants in Communication with Their Mothers. Ph.D. Thesis, The University of Edinburgh, Edinburgh, UK, 1980. [Google Scholar]
- Eibl-Eibesfeldt, I. Ethology: The Biology of Behavior; Holt, Rinehart and Winston: New York, NY, USA, 1970. [Google Scholar]
- Subramanian, S.V.; Jones, K.; Kaddour, A.; Krieger, N. Revisiting Robinson: The perils of individualistic and ecologic fallacy. Int. J. Epidemiol. 2009, 38, 342–360. [Google Scholar] [CrossRef]
- Deeming, C.; Jones, K. Investigating the macro determinants of self-rated health and well-being using the European social survey: Methodological innovations across countries and time. Int. J. Sociol. 2015, 45, 256–285. [Google Scholar] [CrossRef]
- Martin, P.; Bateson PP, G.; Bateson, P. Measuring Behaviour: An Introductory Guide; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Martinez, F.I.; Capelli, C.; Da Silva Maria J Ferreira Aldeias, V.; Alemseged, Z.; Archer, W.; Bamford, M.; Biro, D.; Bobe, R.; Braun, D.R. A missing piece of the Papio puzzle: Gorongosa baboon phenostructure and intrageneric relationships. J. Hum. Evol. 2019, 130, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Bobe, R.; Martinez, F.; Carvalho, S. Primate adaptations and evolution in the southern african rift valley. Evol. Anthropol. 2020, 29, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Santander, C.; Molinaro, L.; Mutti, G.; Martínez, F.I.; Mathe, J.; Ferreira da Silva, M.J.; Caldon, M.; Oteo-Garcia, G.; Aldeias, V.; Archer, W.; et al. Genomic variation in baboons from central mozambique unveils complex evolutionary relationships with other papio species. BMC Ecol. Evol. 2022, 22, 44. [Google Scholar] [CrossRef] [PubMed]
- Hammond, P.; Lewis-Bevan, L.; Biro, D.; Carvalho, S. Risk perception and terrestriality in primates: A quasi-experiment through habituation of chacma baboons (papio ursinus) in gorongosa national park, mozambique. Am. J. Biol. Anthropol. 2022, 179, 48–59. [Google Scholar] [CrossRef]
- Wilson, E.O. A Window on Eternity: A Biologist’s Walk through Gorongosa National Park; Simon and Schuster: New York, NY, USA, 2014. [Google Scholar]
- Watkins, M.W.; Pachego, M. Interobserver agreement in behavioral research: Importance and calculation. J. Behav. Educ. 2000, 10, 205–212. [Google Scholar] [CrossRef]
- Nieuwenhuis, R.; Te Grotenhuis, H.F.; Pelzer, B.J. Influence. ME: Tools for detecting influential data in mixed effects models. R J. 2012, 4, 38–47. [Google Scholar] [CrossRef]
- Bonat, W.H. Multiple response variables regression models in R: The mcglm package. J. Stat. Softw. 2018, 84, 1–30. [Google Scholar] [CrossRef]
- Breheny, P.; Burchett, W.; Breheny, M.P. Package ‘Visreg’. R Package Version 2.7. 2020. Available online: http://pbreheny.github.io/visreg/ (accessed on 16 August 2022).
- Dufon, M.A. The socialization of leave-taking in indonesian. Pragmat. Lang. Learn. 2010, 12, 91–111. [Google Scholar]
- Aureli, F.; Schaffner, C.M. Aggression and conflict management at fusion in spider monkeys. Biol. Lett. 2007, 3, 147–149. [Google Scholar] [CrossRef]
- Lorenz, K.Z. The companion in the bird’s world. Auk 1937, 54, 245–273. [Google Scholar] [CrossRef]
- Okamoto, K.; Agetsuma, N.; Kojima, S. Greeting behavior during party encounters in captive chimpanzees. Primates 2001, 42, 161–165. [Google Scholar] [CrossRef]
- Corsaro, W.A. ‘We’re friends, right?’: Children’s use of access rituals in a nursery school. Lang. Soc. 1979, 8, 315–336. [Google Scholar] [CrossRef]
- Betholia, C. Entries and exits: An analysis of greetings and leave taking in Meitei speech community. Mon-Khmer Stud. 2009, 38, 105. [Google Scholar]
- Goffman, E. Relations in Public: Microstudies of the Social Order; Allen Lane: London, UK, 1971. [Google Scholar]
- Anderson, J.R.; Mcgrew, W.C. Guinea baboons (Papio papio) at a sleeping site. Am. J. Primatol. 1984, 6, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Schegloff, E.A.; Sacks, H. Opening up closings. Semiotica 1973, 8, 289–327. [Google Scholar] [CrossRef]
- Van Der Hart, O.; Goossens, F.A. Leave-taking rituals in mourning therapy. Isr. J. Psychiatry Relat. Sci. 1987, 24, 87–98. [Google Scholar]
- Mondada, L.; Meguerditchian, A. Sequence organization and embodied mutual orientations: Openings of social interactions between baboons. Philos. Trans. R. Soc. B 2022, 377, 20210101. [Google Scholar] [CrossRef]
- Schofield, D.; Nagrani, A.; Zisserman, A.; Hayashi, M.; Matsusawa, T.; Biro, D.; Carvalho, S. Chimpanzee face recognition from videos in the wild using deep learning. Sci. Adv. 2019, 5, eaaw0736. [Google Scholar] [CrossRef]
- Macedonia, J.M.; Evans, C.S. Essay on Contemporary Issues in Ethology: Variation among Mammalian Alarm Call Systems and the Problem of Meaning in Animal Signals; Wiley Online Library: Hoboken, NJ, USA, 1993. [Google Scholar]
- Laidre, M.E.; Johnstone, R.A. Animal signals. Curr. Biol. 2013, 23, R829–R833. [Google Scholar] [CrossRef]
- Troisi, A. Displacement activities as a behavioral measure of stress in nonhuman primates and human subjects. Stress 2002, 5, 47–54. [Google Scholar] [CrossRef]
- Maestripieri, D. Maternal anxiety in rhesus macaques (Macaca mulatta) I. Measurement of anxiety and identification of anxiety-eliciting situations. Ethology 1993, 95, 19–31. [Google Scholar] [CrossRef]
- Dal Pesco, F.; Fischer, J. On the evolution of baboon greeting rituals. Philos. Trans. R. Soc. B 2020, 375, 20190420. [Google Scholar] [CrossRef]
- Washburn, S.L.; Devore, I. The social life of baboons. Sci. Am. 1961, 204, 62–71. [Google Scholar] [CrossRef]
- Goffman, E. Frame Analysis: An Essay on the Organization of Experience; Harvard University Press: Cambridge, MA, USA, 1974. [Google Scholar]
- Fröhlich, M.; Hobaiter, C. The development of gestural communication in great apes. Behav. Ecol. Sociobiol. 2018, 72, 1–14. [Google Scholar] [CrossRef]
- Heesen, R.; Bangerter, A.; Zuberbühler, K.; Iglesias, K.; Neumann, C.; Pajot, A.; Perrenoud, L.; Guéry, J.P.; Rossano, F.; Genty, E. Assessing joint commitment as a process in great apes. IScience 2021, 24, 102872. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solo Departure | Interaction End | Proximity Departure | |
|---|---|---|---|
| Individuals present | Solo individual (no individuals within 1 m) | Dyad (within 1 m) | Dyad (within 1 m) |
| Activity end | End solo activity, e.g., rest, forage, self-groom | End joint activity, e.g., groom, play, or copulate | End solo activity (at least one of the dyad), e.g., rest, forage, or self-groom |
| Movement | Moves away from current spot (>1 m) | Move away from one another (>1 m) | Move away from one another (>1 m) |
| Adult Males | Adult Females | Juveniles | Troop Total |
|---|---|---|---|
| 10 | 12 | 15 | 37 |
| Parting Type | Number of Observations |
|---|---|
| Solo | 100 |
| Proximity | 43 |
| Social | 61 |
| Total | 204 |
| Effects | Estimate | SE | z-Statistics | p-Value | |
|---|---|---|---|---|---|
| Eye gaze | Intercept | −0.0649759376 | 0.3287922059 | −0.1976201 | 0.8433423 |
| Duration | 0.0002494702 | 0.0008174598 | 0.3051774 | 0.7602310 | |
| Parting type (social) | 0.5343620581 | 0.4121778865 | 1.2964355 | 0.1948255 | |
| Parting type (solo) | 0.2090787868 | 0.3727807890 | 0.5608626 | 0.5748912 | |
| Self-scratch | Intercept | −1.106534038 | 0.3591822027 | −3.080704 | 0.002065119 |
| Duration | 0.002242691 | 0.0008856267 | 2.532321 | 0.011331019 | |
| Parting type (social) | 0.526836372 | 0.4339131629 | 1.214152 | 0.224689829 | |
| Parting type (solo) | 0.697408563 | 0.3983954436 | 1.750544 | 0.080024571 | |
| Orientation | Intercept | 0.8690858410 | 0.3574207691 | −2.4315482 | 0.01503445 |
| Duration | 0.0002375434 | 0.0008119219 | 0.2925692 | 0.76985144 | |
| Parting type (social) | 1.0857820340 | 0.4292195876 | 2.5296656 | 0.01141713 * | |
| Parting type (solo) | 0.6328640846 | 0.3964570247 | 1.5962993 | 0.11042198 |
| OR | 2.5% | 97.5% | p-Value | |
|---|---|---|---|---|
| Solo | 1.000 | 0.999 | 1.002 | 0.760 |
| Proximity | 1.706 | 0.761 | 3.828 | 0.195 |
| Social | 1.233 | 0.594 | 2.559 | 0.575 |
| Effects | Estimate | SE | z-Statistics | p-Value | |
|---|---|---|---|---|---|
| Eye gaze | Intercept | 2.87 × 10−4 | 0.1913975773 | −1.50156733 | 0.1332089 |
| Duration | 1.35× 10−5 | 0.0004388482 | 0.03064973 | 0.9755489 | |
| Parting type (social) | 2.12 × 10−1 | 0.2297794091 | 0.92129348 | 0.3568972 | |
| Parting type (solo) | 8.94 × 10−2 | 0.2161082851 | 0.41380489 | 0.6790170 | |
| Self-scratch | Intercept | −1.2261484383 | 0.3109283951 | −3.943507 | 0.0000802985 |
| Duration | 0.0008138371 | 0.0004770604 | 1.705942 | 0.0880189587 | |
| Parting type (social) | 0.5824369726 | 0.3587461767 | 1.623535 | 0.1044750885 | |
| Parting type (solo) | 0.8417835343 | 0.3312493396 | 2.541238 | 0.0110460580 * | |
| Orientation | Intercept | 0.8690858410 | −0.3574207691 | −2.4315482 | 0.01503445 |
| Duration | 0.0002375434 | 0.0008119219 | 0.2925692 | 0.76985144 | |
| Parting type (social) | 1.0857820340 | 0.4292195876 | 2.5296656 | 0.01141713 * | |
| Parting type (solo) | 0.6328640846 | 0.3964570247 | 1.5962993 | 0.11042198 |
| OR | 2.5% | 97.5% | p-Value | |
|---|---|---|---|---|
| Solo | 1.000 | 0.999 | 1.001 | 0.976 |
| Proximity | 1.236 | 0.788 | 1.939 | 0.357 |
| Social | 1.094 | 0.716 | 1.670 | 0.679 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baehren, L.; Carvalho, S. Yet Another Non-Unique Human Behaviour: Leave-Taking in Wild Chacma Baboons (Papio ursinus). Animals 2022, 12, 2577. https://doi.org/10.3390/ani12192577
Baehren L, Carvalho S. Yet Another Non-Unique Human Behaviour: Leave-Taking in Wild Chacma Baboons (Papio ursinus). Animals. 2022; 12(19):2577. https://doi.org/10.3390/ani12192577
Chicago/Turabian StyleBaehren, Lucy, and Susana Carvalho. 2022. "Yet Another Non-Unique Human Behaviour: Leave-Taking in Wild Chacma Baboons (Papio ursinus)" Animals 12, no. 19: 2577. https://doi.org/10.3390/ani12192577
APA StyleBaehren, L., & Carvalho, S. (2022). Yet Another Non-Unique Human Behaviour: Leave-Taking in Wild Chacma Baboons (Papio ursinus). Animals, 12(19), 2577. https://doi.org/10.3390/ani12192577