
Generation and Characterization of Bovine Testicular Organoids Derived from Primary Somatic Cell Populations
 , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Study Design
2.3. Bovine Testis Collection
2.4. Isolation of Bovine Leydig, Sertoli and Peritubular Myoid Cells
2.5. Three-Dimensional Co-Culture of Testicular Cells
2.6. Testicular Organoid Culture Conditions
2.7. Testicular Organoid Whole-Mounting Staining
2.8. Digital Image Acquisition and Processing
2.9. Q-PCR Analysis
2.10. Fluorescence Activated Cell Sorting (FACS) Analysis
2.11. Testosterone Production Assay
2.12. Statistical Analysis
3. Results
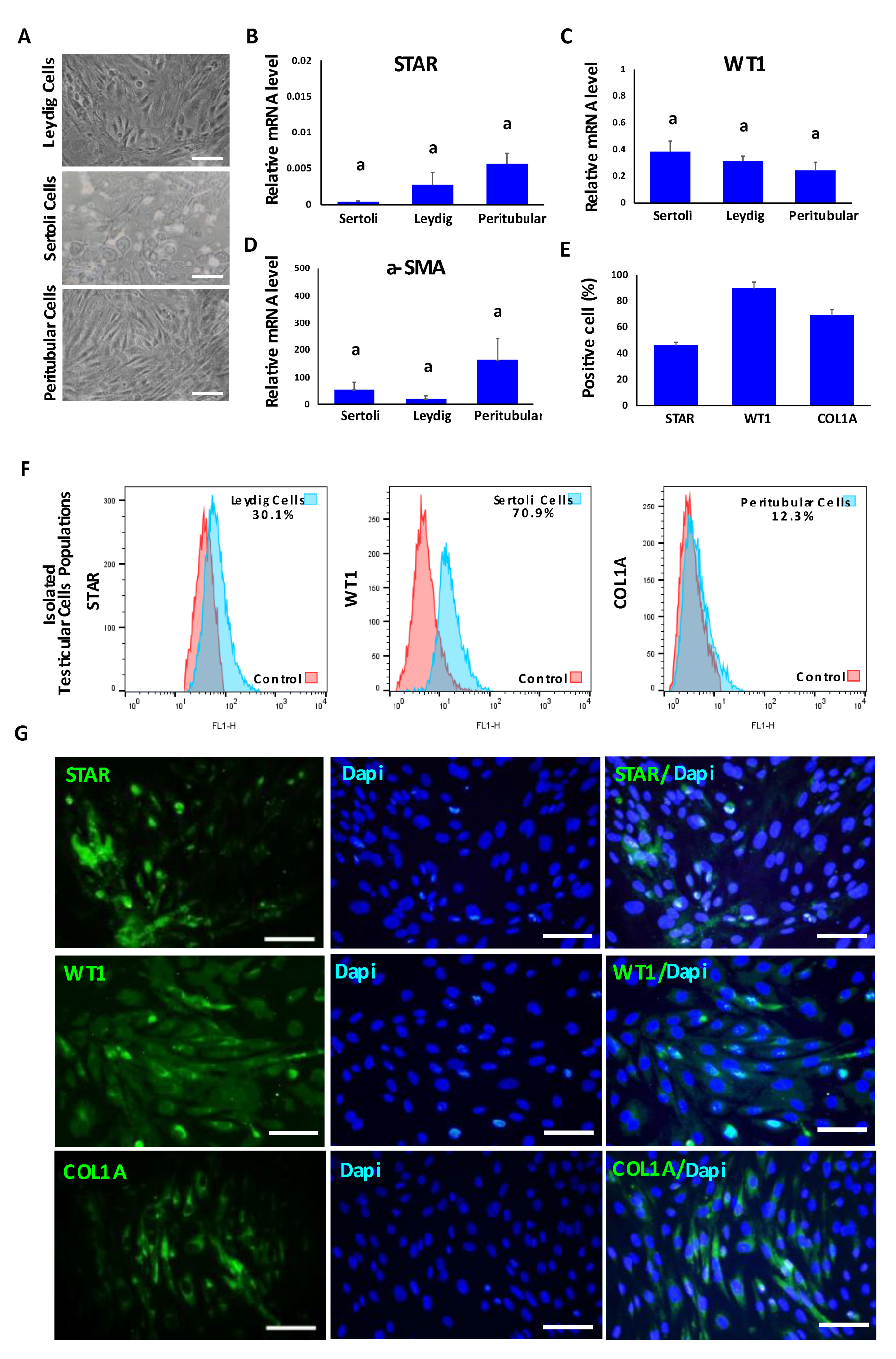
3.1. Isolation and Characterization of Bovine Leydig, Sertoli and Peritubular Myoid Cells
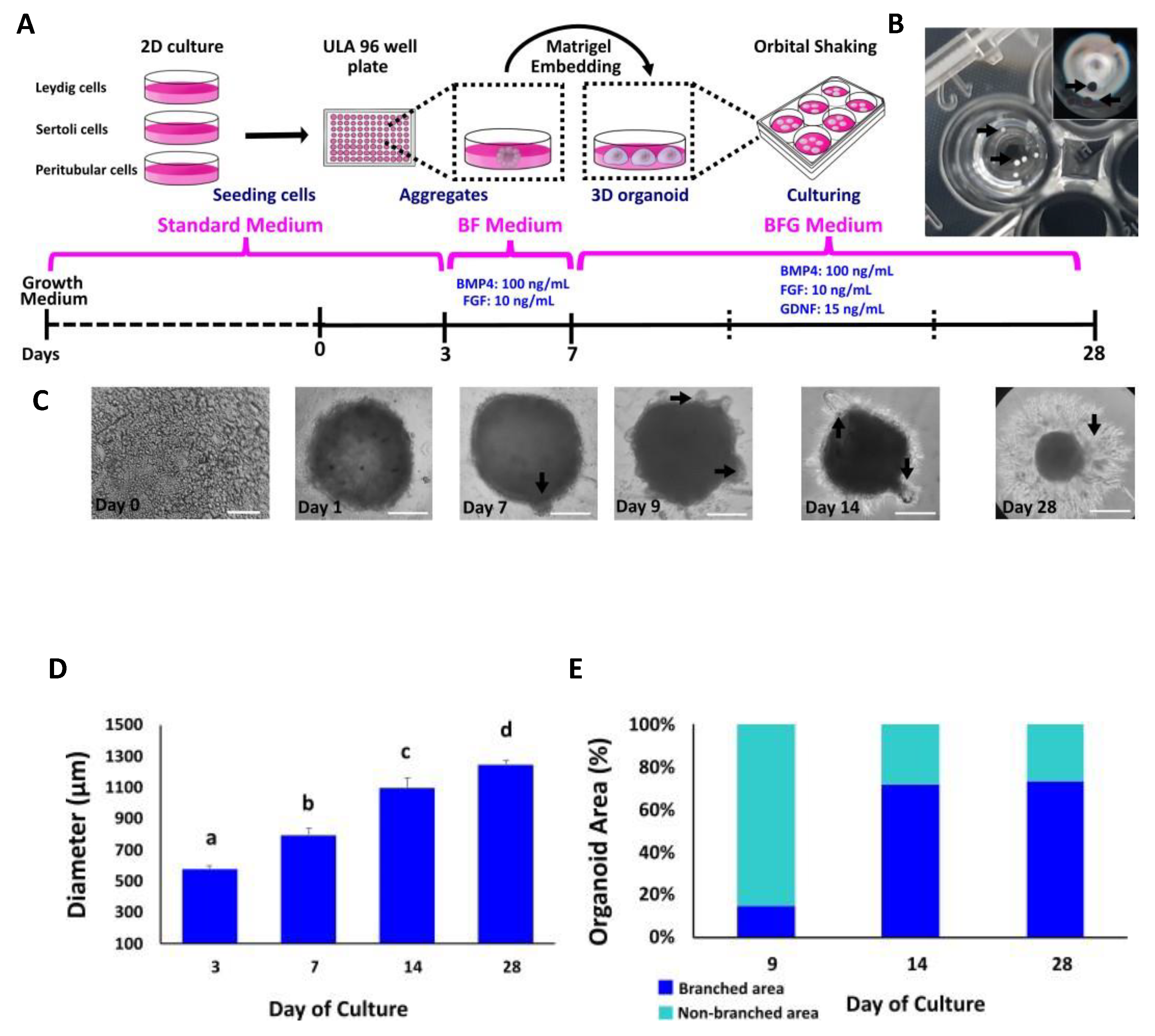
3.2. Formation and Morphological Characterization of Multicellular Bovine TOs
3.3. Testicular Cell Marker Evaluation by QPCR in Bovine TOs
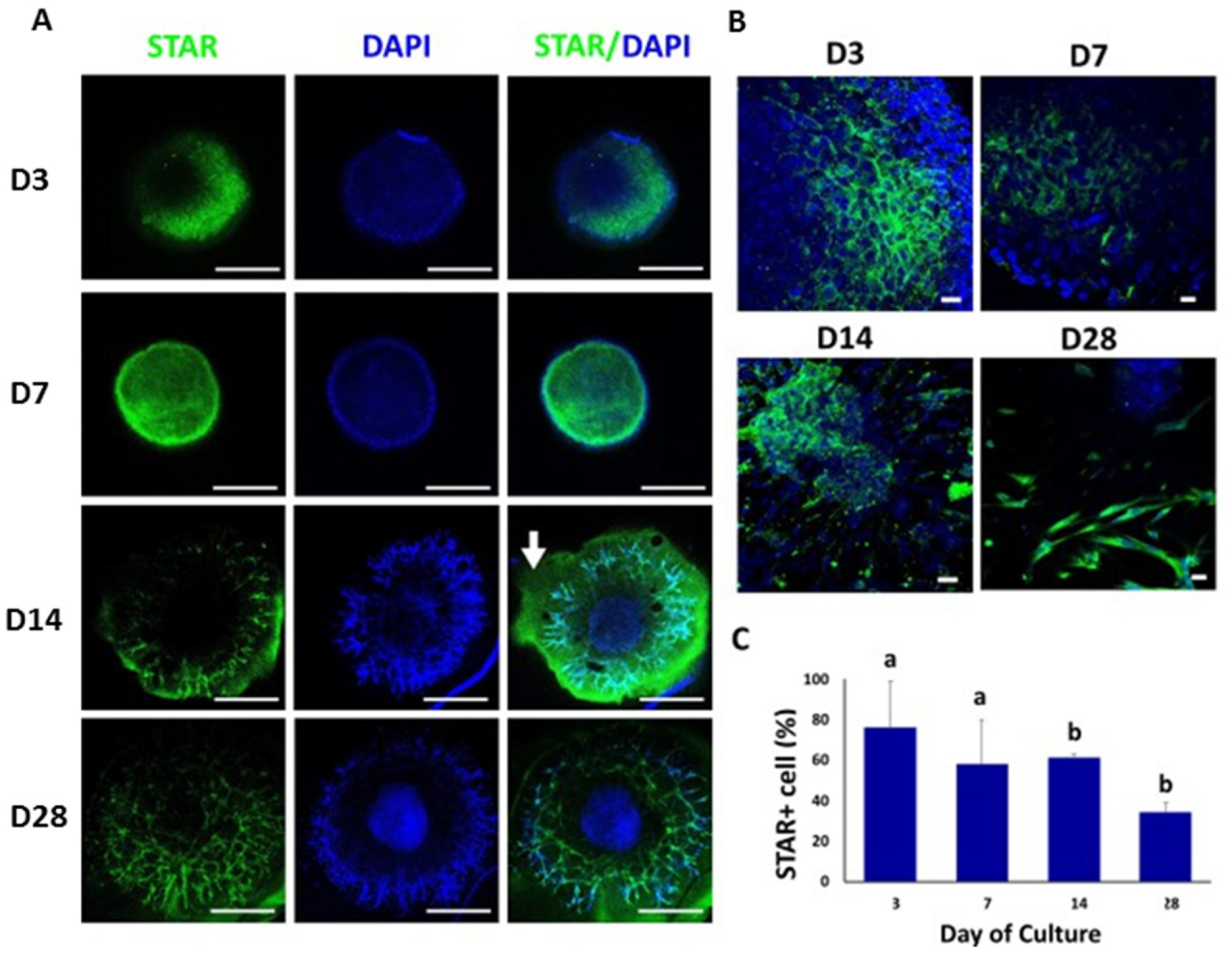
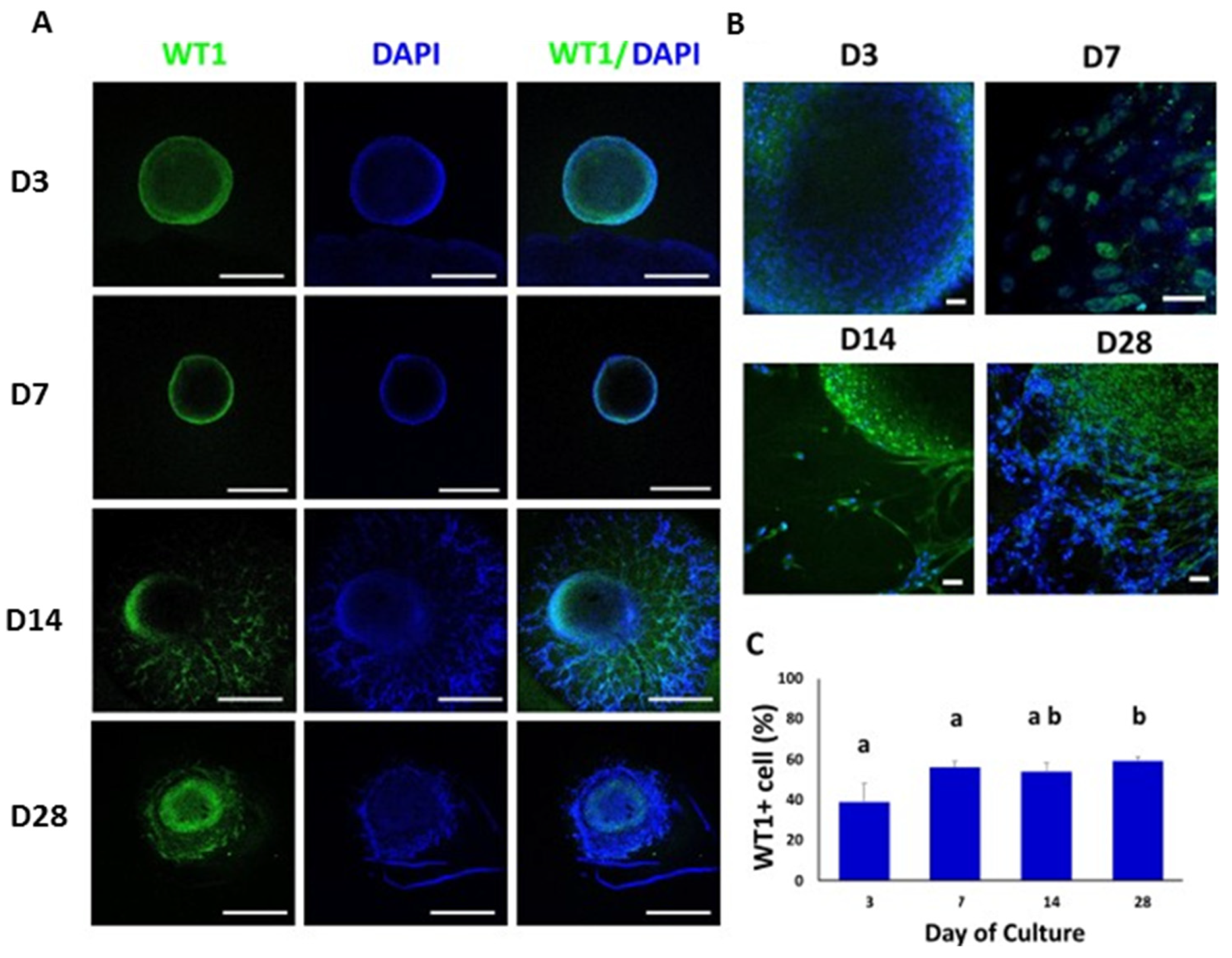
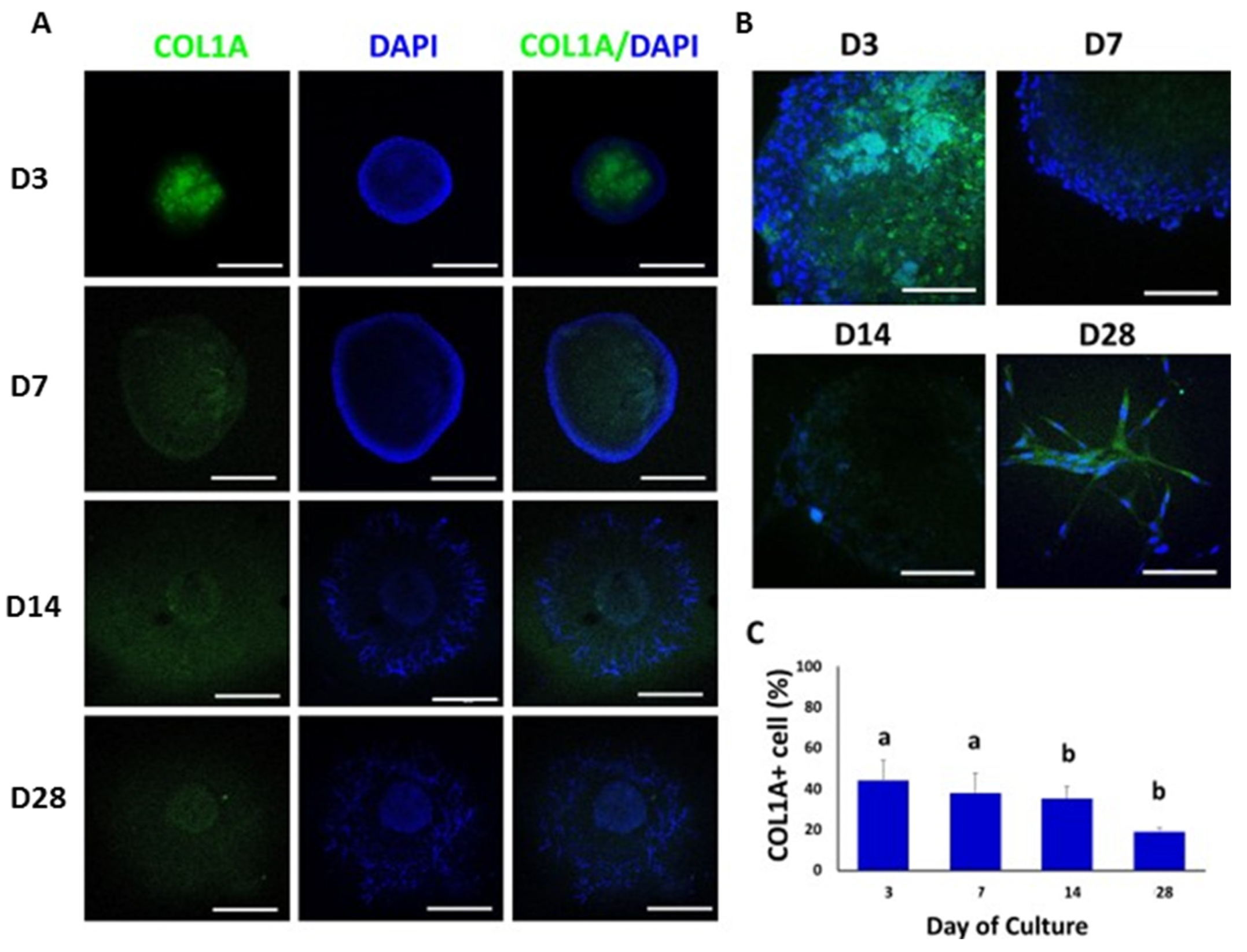
3.4. Confocal Immunofluorescence Analysis of Cell-Specific Testicular Markers in Bovine TOs
3.5. Testosterone Measurements in Testicular Organoids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lancaster, M.A.; Knoblich, J.A. Generation of cerebral organoids from human pluripotent stem cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef] [PubMed]
- Goldsmith, T.M.; Sakib, S.; Webster, D.; Carlson, D.F.; Van der Hoorn, F.; Dobrinski, I. A reduction of primary cilia but not hedgehog signaling disrupts morphogenesis in testicular organoids. Cell Tissue Res. 2020, 380, 191–200. [Google Scholar] [CrossRef]
- Dang, J.; Tiwari, S.K.; Lichinchi, G.; Qin, Y.; Patil, V.S.; Eroshkin, A.M.; Rana, T.M. Zika virus depletes neural progenitors in human cerebral organoids through activation of the innate immune receptor TLR3. Cell Stem Cell 2016, 19, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Fujii, M.; Shimokawa, M.; Date, S.; Takano, A.; Matano, M.; Nanki, K.; Ohta, Y.; Toshimitsu, K.; Nakazato, Y.; Kawasaki, K.; et al. A Colorectal tumor organoid library demonstrates progressive loss of niche factor requirements during tumorigenesis. Cell Stem Cell 2016, 18, 827–838. [Google Scholar] [CrossRef]
- Ettayebi, K.; Crawford, S.E.; Murakami, K.; Broughman, J.R.; Karandikar, U.; Tenge, V.R.; Neill, F.H.; Blutt, S.E.; Zeng, X.L.; Qu, L.; et al. Replication of human noroviruses in stem cell-derived human enteroids. Science 2016, 353, 1387–1393. [Google Scholar] [CrossRef] [PubMed]
- Baert, Y.; Dvorakova-Hortova, K.; Margaryan, H.; Goossens, E. Mouse in vitro spermatogenesis on alginate-based 3D bioprinted scaffolds. Biofabrication 2019, 11, 035011. [Google Scholar] [CrossRef] [PubMed]
- Strange, D.P.; Zarandi, N.P.; Trivedi, G.; Atala, A.; Bishop, C.E.; Sadri-Ardekani, H.; Verma, S. Human testicular organoid system as a novel tool to study Zika virus pathogenesis. Emerg. Microbes Infect. 2018, 7, 82. [Google Scholar] [CrossRef]
- Cham, T.C.; Ibtisham, F.; Fayaz, M.A.; Honaramooz, A. Generation of a highly Biomimetic organoid, including vasculature, resembling the native immature testis tissue. Cells 2021, 10, 1696. [Google Scholar] [CrossRef]
- AbuMadighem, A.; Shuchat, S.; Lunenfeld, E.; Yossifon, G.; Huleihel, M. Testis on a chip-a microfluidic three-dimensional culture system for the development of spermatogenesis in-vitro. Biofabrication 2022, 14, 3. [Google Scholar] [CrossRef]
- Sun, M.; Yuan, Q.; Niu, M.; Wang, H.; Wen, L.; Yao, C.; Hou, J.; Chen, Z.; Fu, H.; Zhou, F.; et al. Efficient generation of functional haploid spermatids from human germline stem cells by three-dimensional-induced system. Cell Death Differ. 2018, 25, 749–766. [Google Scholar] [CrossRef]
- Sadri-Ardekani, H.; Atala, A. Regenerative medicine for the treatment of reproductive system disorders: Current and potential options. Adv. Drug Deliv. Rev. 2015, 82–83, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.F.; Yango, P.; Altman, E.; Choudhry, S.; Poelzl, A.; Zamah, A.M.; Rosen, M.; Klatsky, P.C.; Tran, N.D. Testicular niche required for human spermatogonial stem cell expansion. Stem Cells Transl. Med. 2014, 3, 1043–1054. [Google Scholar] [CrossRef] [PubMed]
- Bhang, D.H.; Kim, B.J.; Kim, B.G.; Schadler, K.; Baek, K.H.; Kim, Y.H.; Hsiao, W.; Ding, B.S.; Rafii, S.; Weiss, M.J.; et al. Testicular endothelial cells are a critical population in the germline stem cell niche. Nat. Commun. 2018, 9, 4379. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Y.; Brown, P.R.; Willis, W.B.; Eddy, E.M. Peritubular myoid cells participate in male mouse spermatogonial stem cell maintenance. Endocrinology 2014, 155, 4964–4974. [Google Scholar] [CrossRef]
- Konrad, L.; Weber, M.A.; Groos, S.; Albrecht, M.; Aumüller, G. Paracrine interaction in testicular somatic cells. Ital. J. Anat. Embryol. 1998, 103, 139–152. [Google Scholar]
- Bernardino, R.L.; Alves, M.G.; Oliveira, P.F. Establishment of primary culture of Sertoli cells. Methods Mol. Biol. 2018, 1748, 1–8. [Google Scholar] [CrossRef]
- Crisóstomo, L.; Alves, M.G.; Gorga, A.; Sousa, M.; Riera, M.F.; Galardo, M.N.; Meroni, S.B.; Oliveira, P.F. Molecular mechanisms and signaling pathways involved in the nutritional support of spermatogenesis by Sertoli cells. Methods Mol. Biol. 2018, 1748, 129–155. [Google Scholar] [CrossRef]
- Shima, Y.; Miyabayashi, K.; Haraguchi, S.; Arakawa, T.; Otake, H.; Baba, T.; Matsuzaki, S.; Shishido, Y.; Akiyama, H.; Tachibana, T.; et al. Contribution of Leydig and Sertoli cells to testosterone production in mouse fetal testes. Mol. Endocrinol. 2013, 27, 63–73. [Google Scholar] [CrossRef]
- Paduch, D.A.; Hilz, S.; Grimson, A.; Schlegel, P.N.; Jedlicka, A.E.; Wright, W.W. Aberrant gene expression by Sertoli cells in infertile men with Sertoli cell-only syndrome. PLoS ONE 2019, 14, e0216586. [Google Scholar] [CrossRef]
- Bhushan, S.; Aslani, F.; Zhang, Z.; Sebastian, T.; Elsässer, H.P.; Klug, J. Isolation of Sertoli cells and peritubular cells from rat testes. J. Vis. Exp. 2016, 8, e53389. [Google Scholar] [CrossRef]
- Russell, L.D.; Bartke, A.; Goh, J.C. Postnatal development of the Sertoli cell barrier, tubular lumen, and cytoskeleton of Sertoli and myoid cells in the rat, and their relationship to tubular fluid secretion and flow. Am. J. Anat. 1989, 184, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Welsh, M.; Moffat, L.; Belling, K.; de França, L.R.; Segatelli, T.M.; Saunders, P.T.; Sharpe, R.M.; Smith, L.B. Androgen receptor signaling in peritubular myoid cells is essential for normal differentiation and function of adult Leydig cells. Int. J. Androl. 2012, 35, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Nistal, M.; Paniagua, R.; Regadera, J.; Santamarìa, L.; Amat, P. A quantitative morphological study of human Leydig cells from birth to adulthood. Cell Tissue Res. 1986, 246, 229–236. [Google Scholar] [CrossRef]
- O’Donnell, L.; McLachlan, R.I.; Wreford, N.G.; de Kretser, D.M.; Robertson, D.M. Testosterone withdrawal promotes stage-specific detachment of round spermatids from the rat seminiferous epithelium. Biol. Reprod. 1996, 55, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Sun, J.; Liu, G.H.; Zhu, Y.J.; Zhu, J. Research on the steroidogenesis of proliferated Leydig cells in vitro. J. Artif. Organs 2013, 16, 229–233. [Google Scholar] [CrossRef]
- Ishiguro, K.I.; Matsuura, K.; Tani, N.; Takeda, N.; Usuki, S.; Yamane, M.; Sugimoto, M.; Fujimura, S.; Hosokawa, M.; Chuma, S.; et al. Meiosin directs the switch from mitosis to meiosis in mammalian germ cells. Dev. Cell 2020, 52, 429–445. [Google Scholar] [CrossRef]
- Guo, J.; Sosa, E.; Chitiashvili, T.; Nie, X.; Rojas, E.J.; Oliver, E.; Plath, K.; Hotaling, J.M.; Stukenborg, J.B.; Clark, A.T.; et al. Single-cell analysis of the developing human testis reveals somatic niche cell specification and fetal germline stem cell establishment. Cell Stem Cell 2021, 28, 764–778. [Google Scholar] [CrossRef]
- Guo, J.; Grow, E.J.; Mlcochova, H.; Maher, G.J.; Lindskog, C.; Nie, X.; Guo, Y.; Takei, Y.; Yun, J.; Cai, L.; et al. The adult human testis transcriptional cell atlas. Cell Res. 2018, 28, 1141–1157. [Google Scholar] [CrossRef]
- Saitou, M. Mammalian germ cell development: From mechanism to in vitro reconstitution. Stem Cell Rep. 2021, 16, 669–680. [Google Scholar] [CrossRef]
- Sakib, S.; Uchida, A.; Valenzuela-Leon, P.; Yu, Y.; Valli-Pulaski, H.; Orwig, K.; Ungrin, M.; Dobrinski, I. Formation of organotypic testicular organoids in microwell culture. Biol. Reprod. 2019, 100, 1648–1660. [Google Scholar] [CrossRef]
- Alves-Lopes, J.P.; Söder, O.; Stukenborg, J.B. Testicular organoid generation by a novel in vitro three-layer gradient system. Biomaterials 2017, 130, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Sakib, S.; Yu, Y.; Voigt, A.; Ungrin, M.; Dobrinski, I. Generation of Porcine Testicular Organoids with testis specific architecture using microwell culture. J. Vis. Exp. 2019, 3, 152. [Google Scholar] [CrossRef] [PubMed]
- Pendergraft, S.S.; Sadri-Ardekani, H.; Atala, A.; Bishop, C.E. Three-dimensional testicular organoid: A novel tool for the study of human spermatogenesis and gonadotoxicity in vitro. Biol. Reprod. 2017, 96, 720–732. [Google Scholar] [CrossRef]
- Sakib, S.; Lara, N.L.E.M.; Huynh, B.C.; Dobrinski, I. Organotypic rat testicular organoids for the study of testicular maturation and toxicology. Front. Endocrinol. 2022, 13, 892342. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, G.; Cadoret, V.; Charpigny, G.; Couturier-Tarrade, A.; Dalbies-Tran, R.; Flores, M.J.; Froment, P.; Raliou, M.; Reynaud, K.; Saint-Dizier, M.; et al. Progress and challenges in developing organoids in farm animal species for the study of reproduction and their applications to reproductive biotechnologies. Vet. Res. 2021, 52, 42. [Google Scholar] [CrossRef] [PubMed]
- Aponte, P.M. Isolation, culture, cryopreservation, and identification of bovine, murine, and human spermatogonial stem cells. Methods Mol. Biol. 2020, 2155, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Raval, N.P.; Shah, T.M.; George, L.B.; Joshi, C.G. Insight into bovine (Bos indicus) spermatozoal whole transcriptome profile. Theriogenology 2019, 129, 8–13. [Google Scholar] [CrossRef]
- Aponte, P.M.; Soda, T.; van de Kant, H.J.; de Rooij, D.G. Basic features of bovine spermatogonial culture and effects of glial cell line-derived neurotrophic factor. Theriogenology 2006, 65, 1828–1847. [Google Scholar] [CrossRef]
- Gaur, M.; Ramathal, C.; Reijo Pera, R.A.; Turek, P.J.; John, C.M. Isolation of human testicular cells and co-culture with embryonic stem cells. Reproduction 2018, 155, 153–166. [Google Scholar] [CrossRef]
- Drost, J.; Karthaus, W.R.; Gao, D.; Driehuis, E.; Sawyers, C.L.; Chen, Y.; Clevers, H. Organoid culture systems for prostate epithelial and cancer tissue. Nat. Protoc. 2016, 11, 347–358. [Google Scholar] [CrossRef]
- Sahu, S.; Albaugh, M.E.; Martin, B.K.; Patel, N.L.; Riffle, L.; Mackem, S.; Kalen, J.D.; Sharan, S.K. Growth factor dependency in mammary organoids regulates ductal morphogenesis during organ regeneration. Sci. Rep. 2022, 12, 7200. [Google Scholar] [CrossRef] [PubMed]
- Kiani, M.; Movahedin, M.; Halvaei, I.; Soleimani, M. Formation of organoid-like structures in the decellularized rat testis. Iran. J. Basic Med. Sci. 2021, 24, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, L.; Guo, Y.; Zhu, Y.; Qin, J. Engineering stem cell-derived 3D brain organoids in a perfusable organ-on-a-chip system. RSC Adv. 2018, 8, 1677–1685. [Google Scholar] [CrossRef] [PubMed]
- Takasato, M.; Er, P.X.; Chiu, H.S.; Little, M.H. Generation of kidney organoids from human pluripotent stem cells. Nat. Protoc. 2016, 11, 1681–1692. [Google Scholar] [CrossRef]
- Sato, T.; Vries, R.G.; Snippert, H.J.; van de Wetering, M.; Barker, N.; Stange, D.E.; van Es, J.H.; Abo, A.; Kujala, P.; Peters, P.J.; et al. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature 2009, 459, 262–265. [Google Scholar] [CrossRef] [PubMed]
- Vishwakarma, A.; Rouwkema, J.; Jones, P.A.; Karp, J.M. The need to study, mimic, and target stem cell niches. In Biology and Engineering of Stem Cell Niches; Vishwakarma, A., Karp, J.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 3–13. [Google Scholar] [CrossRef]
- Vyas, D.; Baptista, P.M.; Brovold, M.; Moran, E.; Gaston, B.; Booth, C.; Samuel, M.; Atala, A.; Soker, S. Self-assembled liver organoids recapitulate hepatobiliary organogenesis in vitro. Hepatology 2018, 67, 750–761. [Google Scholar] [CrossRef]
- Baert, Y.; Rombaut, C.; Goossens, E. Scaffold-Based and Scaffold-Free Testicular organoids from primary human testicular cells. Methods Mol. Biol. 2019, 1576, 283–290. [Google Scholar] [CrossRef]
- Cham, T.C.; Chen, X.; Honaramooz, A. Current progress, challenges, and future prospects of testis organoids. Biol. Reprod. 2021, 104, 942–961. [Google Scholar] [CrossRef] [PubMed]
- Topraggaleh, T.; Rezazadeh Valojerdi, M.; Montazeri, L.; Baharvand, H. A testis-derived macroporous 3D scaffold as a platform for the generation of mouse testicular organoids. Biomater. Sci. 2019, 7, 1422–1436. [Google Scholar] [CrossRef]
- Mall, E.M.; Rotte, N.; Yoon, J.; Sandhowe-Klaverkamp, R.; Röpke, A.; Wistuba, J.; Hübner, K.; Schöler, H.R.; Schlatt, S. A novel xeno-organoid approach: Exploring the crosstalk between human iPSC-derived PGC-like and rat testicular cells. Mol. Hum. Reprod. 2020, 26, 879–893. [Google Scholar] [CrossRef]
- Banerjee, A.; Arha, M.; Choudhary, S.; Ashton, R.S.; Bhatia, S.R.; Schaffer, D.V.; Kane, R.S. The influence of hydrogel modulus on the proliferation and differentiation of encapsulated neural stem cells. Biomaterials 2009, 30, 4695–4699. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, M.; Del Vento, F.; Kanbar, M.; Pyr Dit Ruys, S.; Vertommen, D.; Poels, J.; Wyns, C. Generation of organized porcine testicular organoids in solubilized hydrogels from decellularized extracellular matrix. Int. J. Mol. Sci. 2019, 20, 5476. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, M.E.; Forshee, M.D.; Woodruff, T.K. Extra cellular matrix-based and extra cellular matrix-free generation of murine testicular organoids. J. Vis. Exp. 2020, 7, 164. [Google Scholar] [CrossRef] [PubMed]
- Edmonds, M.E.; Woodruff, T.K. Testicular organoid formation is a property of immature somatic cells, which self-assemble and exhibit long-term hormone-responsive endocrine function. Biofabrication 2020, 12, 045002. [Google Scholar] [CrossRef] [PubMed]
- McMurtrey, R.J. Analytic models of oxygen and nutrient diffusion, metabolism dynamics, and architecture optimization in three-dimensional tissue constructs with applications and insights in cerebral organoids. Tissue Eng. Part C Methods 2016, 22, 221–249. [Google Scholar] [CrossRef] [PubMed]
- Berger, E.; Magliaro, C.; Paczia, N.; Monzel, A.S.; Antony, P.; Linster, C.L.; Bolognin, S.; Ahluwalia, A.; Schwamborn, J.C. Millifluidic culture improves human midbrain organoid vitality and differentiation. Lab Chip 2018, 18, 3172–3183. [Google Scholar] [CrossRef]
- Richer, G.; Hobbs, R.M.; Loveland, K.L.; Goossens, E.; Baert, Y. Long-term maintenance and meiotic entry of early germ cells in murine testicular organoids functionalized by 3D printed scaffolds and air-medium interface cultivation. Front. Physiol. 2021, 24, 12. [Google Scholar] [CrossRef]
- Oliver, E.; Alves-Lopes, J.P.; Harteveld, F.; Mitchell, R.T.; Åkesson, E.; Söder, O.; Stukenborg, J.B. Self-organising human gonads generated by a Matrigel-based gradient system. BMC Biol. 2021, 19, 212. [Google Scholar] [CrossRef]
- Koo, B.; Choi, B.; Park, H.; Yoon, K.J. Past, present, and Ffuture of brain organoid technology. Mol. Cells 2019, 42, 617–627. [Google Scholar] [CrossRef]
- Sivitilli, A.A.; Gosio, J.T.; Ghoshal, B.; Evstratova, A.; Trcka, D.; Ghiasi, P.; Hernandez, J.J.; Beaulieu, J.M.; Wrana, J.L.; Attisano, L. Robust production of uniform human cerebral organoids from pluripotent stem cells. Life Sci. Alliance 2020, 3, 5. [Google Scholar] [CrossRef]
- Pellegrini, L.; Bonfio, C.; Chadwick, J.; Begum, F.; Skehel, M.; Lancaster, M.A. Human CNS barrier-forming organoids with cerebrospinal fluid production. Science 2020, 10, 369. [Google Scholar] [CrossRef]
- Choe, M.S.; Kim, S.J.; Oh, S.T.; Bae, C.M.; Choi, W.Y.; Baek, K.M.; Kim, J.S.; Lee, M.Y. A simple method to improve the quality and yield of human pluripotent stem cell-derived cerebral organoids. Heliyon 2021, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Nengzhuang, W.; Jiaming, S.; Minghua, L.I.U.; Long, M.A.; Lina, Q.I.N.; Xuemei, G.E.; Hongli, Y.A.N. A brief history of testicular organoids: From theory to the wards. J. Assist. Reprod. Genet. 2022, 39, 1423–1431. [Google Scholar] [CrossRef]
- Miller, W.L. Steroidogenic acute regulatory protein (StAR), a novel mitochondrial cholesterol transporter. Biochim. Biophys. Acta 2007, 1771, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Galano, M.; Li, Y.; Li, L.; Sottas, C.; Papadopoulos, V. Role of constitutive STAR in Leydig cells. Int. J. Mol. Sci. 2021, 22, 2021. [Google Scholar] [CrossRef]
- Chung, J.Y.; Brown, S.; Chen, H.; Liu, J.; Papadopoulos, V.; Zirkin, B. Effects of pharmacologically induced Leydig cell testosterone production on intratesticular testosterone and spermatogenesis. Biol. Reprod. 2020, 14, 489–498. [Google Scholar] [CrossRef]
- Lara, N.L.E.M.; Sakib, S.; Dobrinski, I. Regulation of Cell Types within Testicular Organoids. Endocrinology 2021, 162, bqab033. [Google Scholar] [CrossRef] [PubMed]
- Mincheva, M.; Sandhowe-Klaverkamp, R.; Wistuba, J.; Redmann, K.; Stukenborg, J.B.; Kliesch, S.; Schlatt, S. Reassembly of adult human testicular cells: Can testis cord-like structures be created in vitro? Mol. Hum. Reprod. 2018, 24, 55–63. [Google Scholar] [CrossRef]
- Baert, Y.; De Kock, J.; Alves-Lopes, J.P.; Söder, O.; Stukenborg, J.B.; Goossens, E. Primary human testicular cells self-organize into organoids with testicular properties. Stem Cell Rep. 2017, 8, 30–38. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Cell Marker | Cat.# | P/N Company | Dilution |
|---|---|---|---|---|
| Anti-COL1 | Peritubular | SC293182 | Santa Cruz Biotechonology | 1:50 |
| Anti-STAR | Leydig | AB96637 | Abcam | 1:100 |
| Anti-WT1 | Sertoli | AB89901 | Abcam | 1:100 |
| IgG FITC | - | AB97050 | Abcam | 1:100 |
| IgG Alexa Fluor 488 | - | A32766 | Thermo Fisher | 1:500 |
| Gene | Nucleotide Sequence (5′-3′) | Accession Number |
|---|---|---|
| β-ACTIN | Forward CGCACCACTGGCATTGTCAT Reverse TCCAAGGCGACGTAGCAGAG | NM_173979.3 |
| WT1 * | Forward AACCACACCACACCCATCC Reverse ACGCCGCACATCCTGAAT | XM_015466595.1 |
| STAR * | Forward GACACGGTCATCACTCACGA Reverse TACGCTCACAAAGTCTCGGG | NM_174189.3 |
| αSMA ** | Forward CAGCCGAGAACTTTCAGGGAC Reverse GGTGATGATGCCGTGCTCTA | NM_001034502.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortez, J.; Leiva, B.; Torres, C.G.; Parraguez, V.H.; De los Reyes, M.; Carrasco, A.; Peralta, O.A. Generation and Characterization of Bovine Testicular Organoids Derived from Primary Somatic Cell Populations. Animals 2022, 12, 2283. https://doi.org/10.3390/ani12172283
Cortez J, Leiva B, Torres CG, Parraguez VH, De los Reyes M, Carrasco A, Peralta OA. Generation and Characterization of Bovine Testicular Organoids Derived from Primary Somatic Cell Populations. Animals. 2022; 12(17):2283. https://doi.org/10.3390/ani12172283
Chicago/Turabian StyleCortez, Jahaira, Barbara Leiva, Cristian G. Torres, Víctor H. Parraguez, Mónica De los Reyes, Albert Carrasco, and Oscar A. Peralta. 2022. "Generation and Characterization of Bovine Testicular Organoids Derived from Primary Somatic Cell Populations" Animals 12, no. 17: 2283. https://doi.org/10.3390/ani12172283
APA StyleCortez, J., Leiva, B., Torres, C. G., Parraguez, V. H., De los Reyes, M., Carrasco, A., & Peralta, O. A. (2022). Generation and Characterization of Bovine Testicular Organoids Derived from Primary Somatic Cell Populations. Animals, 12(17), 2283. https://doi.org/10.3390/ani12172283