
Effect of Different Ambient Temperatures on Reproductive Outcome and Stress Level of Lactating Females in Two Mouse Strains

 , , and
, , and
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Husbandry Conditions
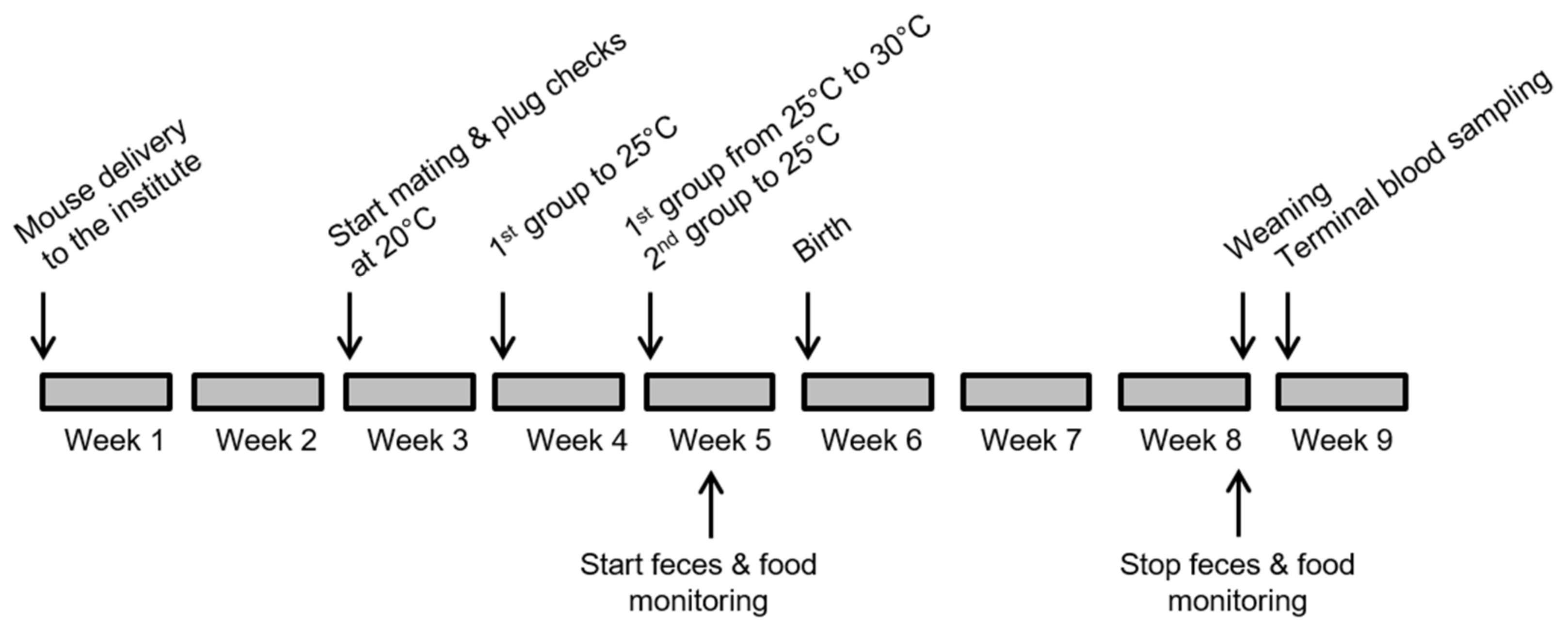
2.2. Experimental Temperature Groups
2.3. Experimental Measurements
2.4. Implantation Sites
2.5. Analysis of Faecal Corticosterone Metabolites and Plasma Corticosterone
2.6. Statistical Procedures
3. Results
3.1. Cage Temperature and Room Humidity
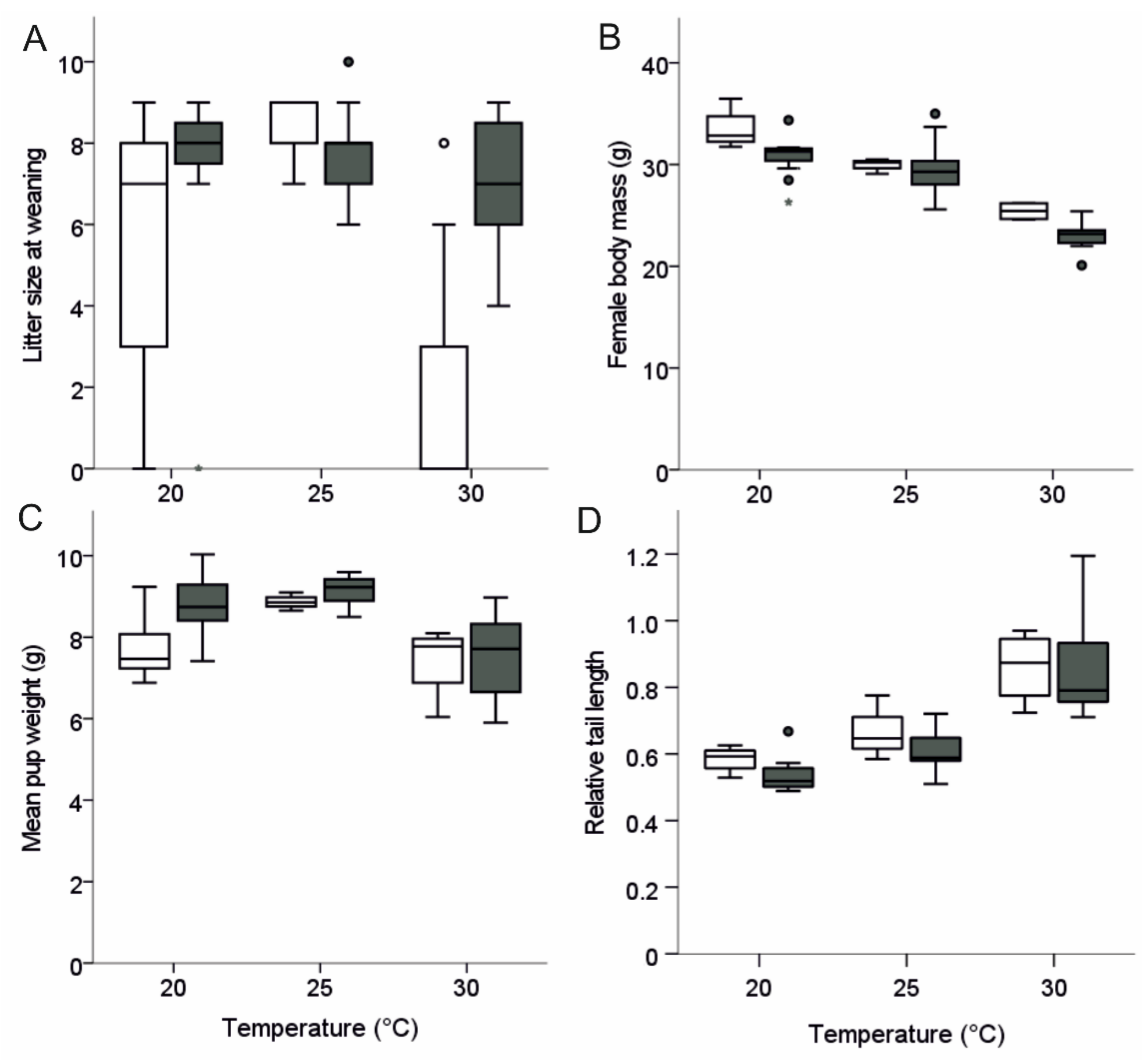
3.2. Reproductive Parameters
3.3. Weight and Tail Length
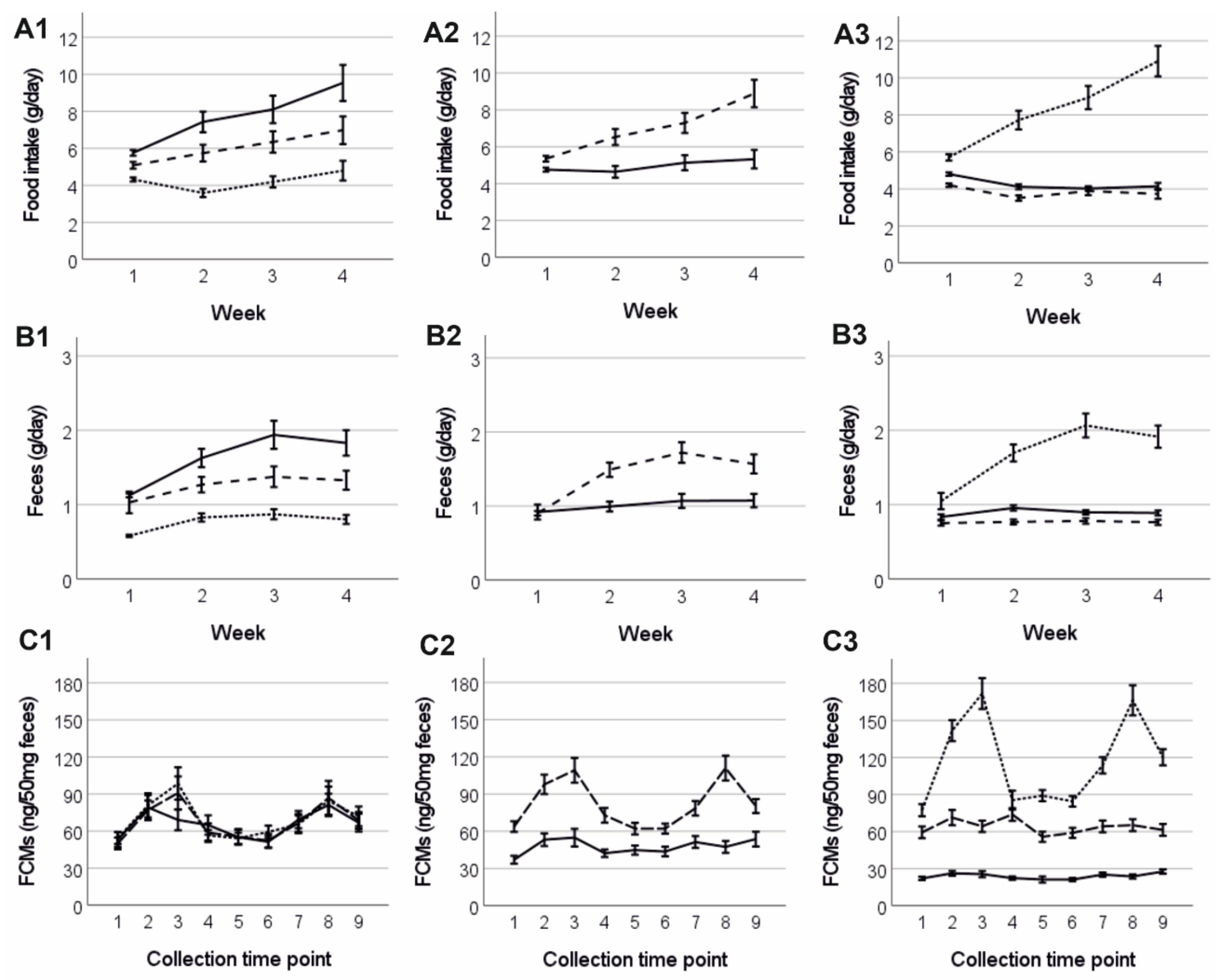
3.4. Food Consumption and Amount of Faeces
3.5. Faecal Corticosterone Metabolites (FCMs) and Plasma Corticosterone
4. Discussion
4.1. Reproduction
4.2. Physiological and Morphological Changes
4.3. Glucocorticoids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gordon, C.J.; Becker, P.; Ali, J.S. Behavioral thermoregulatory responses of single- and group-housed mice. Physiol. Behav. 1998, 65, 255–262. [Google Scholar] [CrossRef]
- Hylander, B.L.; Repasky, E.A. Thermoneutrality, mice, and cancer: A heated opinion. Trends Cancer 2016, 2, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Jhaveri, K.A.; Trammell, R.A.; Toth, L.A. Effect of environmental temperature on sleep, locomotor activity, core body temperature and immune responses of C57BL/6J mice. Brain Behav. Immun. 2007, 21, 975–987. [Google Scholar] [CrossRef] [PubMed]
- Repasky, E.A.; Evans, S.S.; Dewhirst, M.W. Temperature matters! And why it should matter to tumor immunologists. Cancer Immunol. Res. 2013, 1, 210–216. [Google Scholar] [CrossRef]
- Giles, D.A.; Ramkhelawon, B.; Donelan, E.M.; Stankiewicz, T.E.; Hutchison, S.B.; Mukherjee, R.; Cappelletti, M.; Karns, R.; Karp, C.L.; Moore, K.J.; et al. Modulation of ambient temperature promotes inflammation and initiates atherosclerosis in wild type C57BL/6 mice. Mol. Metab. 2016, 5, 1121–1130. [Google Scholar] [CrossRef]
- Hylander, B.L.; Gordon, C.J.; Repasky, E.A. Manipulation of ambient housing temperature to study the impact of chronic stress on immunity and cancer in mice. J. Immunol. 2019, 202, 631–636. [Google Scholar] [CrossRef]
- Swoap, S.K.; Overton, J.M.; Garber, G. Effect of ambient temperature on cardiovascular parameters in rats and mice: A comparative approach. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R391–R396. [Google Scholar] [CrossRef]
- Ostberg, J.R.; Taylor, S.L.; Baumann, H.; Repasky, E.A. Regulatory effects of fever-range whole-body hyperthermia on the LPS-induced acute inflammatory response. J. Leukoc. Biol. 2000, 68, 815–820. [Google Scholar] [CrossRef]
- Ostberg, J.R.; Gellin, C.; Patel, R.; Repasky, E.A. Regulatory potential of fever-range whole body hyperthermia on Langerhans cells and lymphocytes in an antigen-dependent cellular immune response. J. Immunol. 2001, 167, 2666–2670. [Google Scholar] [CrossRef]
- Ostberg, J.R.; Kaplan, K.C.; Repasky, E.A. Induction of stress proteins in a panel of mouse tissues by fever-range whole body hyperthermia. Int. J. Hyperthermia 2002, 18, 552–562. [Google Scholar] [CrossRef]
- Eng, J.W.-L.; Reed, C.B.; Kokolus, K.M.; Repasky, E.A. Housing temperature influences the pattern of heat shock protein induction in mice following mild whole body hyperthermia. Int. J. Hyperthermia 2014, 30, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Zynda, E.R.; Grimm, M.J.; Yuan, M.; Zhong, L.; Mace, T.A.; Capitano, M.; Ostberg, J.R.; Lee, K.P.; Pralle, A.; Repasky, E.A. A role for the thermal environment in defining co-stimulation requirements for CD4(+) T cell activation. Cell Cycle 2015, 14, 2340–2354. [Google Scholar] [CrossRef] [PubMed]
- Sareh, H.; Tulapurkar, M.E.; Shah, N.G.; Singh, I.S.; Hasday, J.D. Response of mice to continuous 5-day passive hyperthermia resembles human heat acclimation. Cell Stress Chaperones 2011, 16, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Cannon, B.; Nedergaard, J. Nonshivering thermogenesis and its adequate measurement in metabolic studies. J. Exp. Biol. 2011, 214, 242–253. [Google Scholar] [CrossRef] [PubMed]
- Skop, V.; Guo, J.; Liu, N.; Xiao, C.; Hall, K.D.; Gavrilova, O.; Reitman, M.L. Mouse thermoregulation: Introducing the concept of the thermoneutral point. Cell Rep. 2020, 31, 107501. [Google Scholar] [CrossRef]
- Gordon, C.J.; Aydin, C.; Repasky, E.A.; Kokolus, K.M.; Dheyongera, G.; Johnstone, A.F. Behaviorally mediated, warm adaptation: A physiological strategy when mice behaviorally thermoregulate. J. Therm. Biol. 2014, 44, 41–46. [Google Scholar] [CrossRef]
- Gaskill, B.N.; Rohr, S.A.; Pajor, E.A.; Lucas, J.R.; Garner, J.P. Some like it hot: Mouse temperature preferences in laboratory housing. Appl. Anim. Behav. Sci. 2009, 116, 279–285. [Google Scholar] [CrossRef]
- Gaskill, B.N.; Gordon, C.J.; Pajor, E.A.; Lucas, J.R.; Davis, J.K.; Garner, J.P. Heat or insulation: Behavioral titration of mouse preference for warmth or access to a nest. PLoS ONE 2012, 7, e32799. [Google Scholar] [CrossRef]
- Gordon, C.J. Effect of cage bedding on temperature regulation and metabolism of group-housed female mice. Comp. Med. 2004, 54, 63–68. [Google Scholar]
- Gaskill, B.N.; Gordon, C.J.; Pajor, E.A.; Lucas, J.R.; Davis, J.K.; Garner, J.P. Impact of nesting material on mouse body temperature and physiology. Physiol. Behav. 2013, 110–111, 87–95. [Google Scholar] [CrossRef]
- Gaskill, B.N.; Karas, A.Z.; Garner, J.P.; Pritchett-Corning, K.R. Nest building as an indicator of health and welfare in laboratory mice. J. Vis. Exp. 2013, 82, 51012. [Google Scholar] [CrossRef] [PubMed]
- Gaskill, B.N.; Winnicker, C.; Garner, J.P.; Pritchett-Corning, K.R. The naked truth: Breeding performance in nude mice with and without nesting material. Appl. Anim. Behav. Sci. 2013, 143, 110–116. [Google Scholar] [CrossRef]
- Gaskill, B.N.; Pritchett-Corning, K.R.; Gordon, C.J.; Pajor, E.A.; Lucas, J.R.; Davis, J.K.; Garner, J.P. Energy reallocation to breeding performance through improved nest building in laboratory mice. PLoS ONE 2013, 8, e74153. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.S.; Taylor, D.J.; Green, A.R.; Gaskill, B.N. Effects of nesting material on energy homeostasis in BALB/cAnNCrl, C57BL/6NCrl, and Crl:CD1(ICR) mice housed at 20 °C. J. Am. Assoc. Lab. Anim. Sci. 2017, 56, 254–259. [Google Scholar]
- Maher, R.L.; Barbash, S.M.; Lynch, D.V.; Swoap, S.J. Group housing and nest building only slightly ameliorate the cold stress of typical housing in female C57BL/6J mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R1070–R1079. [Google Scholar] [CrossRef]
- Kingma, B.R.M.; Frijns, A.J.H.; Schellen, L.; Van Marken Lichtenbelt, W.D. Beyond the classic thermoneutral zone: Including thermal comfort. Temperature 2014, 1, 142–149. [Google Scholar] [CrossRef]
- Speakman, J.R.; Keijer, J. Not so hot: Optimal housing temperatures for mice to mimic the thermal environment of humans. Mol. Metab. 2012, 2, 5–9. [Google Scholar] [CrossRef]
- Oufara, S.; Barré, H.; Rouanet, J.L.; Chatonnet, J. Adaptation to extreme ambient temperatures in cold-acclimated gerbils and mice. Am. J. Physiol. 1987, 253, R39–R45. [Google Scholar] [CrossRef]
- Van Bogaert, M.J.; Groenink, L.; Oosting, R.S.; Westphal, K.G.; van der Gugten, J.; Olivier, B. Mouse strain differences in autonomic responses to stress. Genes Brain Behav. 2006, 5, 139–149. [Google Scholar] [CrossRef]
- Van Loo, P.L.; Kuin, N.; Sommer, R.; Avsaroglu, H.; Pham, T.; Baumans, V. Impact of ‘living apart together’ on postoperative recovery of mice compared with social and individual housing. Lab. Anim. 2007, 41, 441–455. [Google Scholar] [CrossRef]
- Abreu-Vieira, G.; Xiao, C.; Gavrilova, O.; Reitman, M.L. Integration of body temperature into the analysis of energy expenditure in the mouse. Mol. Metab. 2015, 4, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Toth, L.A.; Trammell, R.A.; Ilsley-Woods, M. Interactions between housing density and ambient temperature in the cage environment: Effects on mouse physiology and behavior. J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 708–717. [Google Scholar] [PubMed]
- Habicht, G.S. Body temperature in normal and endotoxin-treated mice of different ages. Mech. Ageing Dev. 1981, 16, 97–104. [Google Scholar] [CrossRef]
- Moragues, V.; Pinkerton, H. Variation in morbidity and mortality of murine typhus infection in mice with changes in the environmental temperature. J. Exp. Med. 1944, 79, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Kokolus, K.M.; Capitano, M.L.; Lee, C.-T.; Eng, J.W.-L.; Waight, J.D.; Hylander, B.L.; Sexton, S.; Hong, C.-C.; Gordon, C.J.; Abrams, S.I.; et al. Baseline tumor growth and immune control in laboratory mice are significantly influenced by subthermoneutral housing temperature. Proc. Natl. Acad. Sci. USA 2013, 110, 20176–20181. [Google Scholar] [CrossRef] [PubMed]
- Kaikaew, K.; Steenbergen, J.; Themmen, A.P.N.; Visser, J.A.; Grefhorst, A. Sex difference in thermal preference of adult mice does not depend on presence of the gonads. Biol. Sex Differ. 2017, 8, 24. [Google Scholar] [CrossRef]
- Yamauchi, C.; Fujita, S.; Obara, T.; Ueda, T. Effects of room temperature on reproduction, body and organ weights, food and water intakes, and hematology in mice. Exp. Anim. 1983, 32, 1–11. [Google Scholar] [CrossRef]
- Helppi, J.; Schreier, D.; Naumann, R.; Zierau, O. Mouse reproductive fitness is maintained up to an ambient temperature of 28 when housed in individually ventilated cages. Lab. Anim. 2016, 50, 254–263. [Google Scholar] [CrossRef]
- Berteotti, C.; Lo Martire, V.; Alvente, S.; Bastianini, S.; Matteoli, G.; Silvani, A.; Zoccoli, G. Effect of ambient temperature on sleep breathing phenotype in mice: The role of orexins. J. Exp. Biol. 2020, 223, jeb.219485. [Google Scholar] [CrossRef]
- Greenberg, G. The effects of ambient temperature and population density on aggression in two inbred strains of mice, Mus musculus. Behaviour 1972, 42, 119–130. [Google Scholar] [CrossRef]
- Kolbe, T.; Palme, R.; Tichy, A.; Rülicke, T. Lifetime dependent variation of stress hormone metabolites in faeces of two laboratory mouse strains. PLoS ONE 2015, 10, e0136112. [Google Scholar] [CrossRef] [PubMed]
- Kolbe, T.; Sheety, S.; Walter, I.; Palme, R.; Rülicke, T. Impact of superovulation and mating on the wellbeing of juvenile and adult C57BL/6N mice. Reprod. Fert. Dev. 2016, 28, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Razzoli, M.; Nyuyki-Dufe, K.; Gurney, A.; Erickson, C.; McCallum, J.; Spielman, N.; Marzullo, M.; Patricelli, J.; Kurata, M.; Pope, E.A.; et al. Social stress shortens lifespan in mice. Aging Cell 2018, 17, e12778. [Google Scholar] [CrossRef] [PubMed]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef]
- Gamo, Y.; Bernard, A.; Mitchell, S.E.; Hambly, C.; Jothery, A.A.; Vaanholt, L.M.; Król, E.; Speakman, J.R. Limits to sustained energy intake. XVI. Body temperature and physical activity of female mice during pregnancy. J. Exp. Biol. 2013, 216, 2328–2338. [Google Scholar] [CrossRef]
- Gamo, Y.; Troup, C.; Mitchell, S.E.; Hambly, C.; Vaanholt, L.M.; Speakman, J.R. Limits to sustained energy intake. XX. Body temperatures and physical activity of female mice during lactation. J. Exp. Biol. 2013, 216, 3751–3761. [Google Scholar] [CrossRef]
- Salewski, E. Färbemethode zum makroskopischen Nachweis von Implantationsstellen am Uterus der Ratte. Naunyn-Schmiedebergs Archiv für experimentelle Pathologie und Pharmakologie 1964, 247, 367. [Google Scholar] [CrossRef]
- Touma, C.; Sachser, N.; Mostl, E.; Palme, R. Effects of sex and time of day on metabolism and excretion of corticosterone in urine and faeces of mice. Gen. Comp. Endocrinol. 2003, 130, 267–278. [Google Scholar] [CrossRef]
- Touma, C.; Palme, R.; Sachser, N. Analyzing corticosterone metabolites in fecal samples of mice: A noninvasive technique to monitor stress hormones. Horm. Behav. 2004, 45, 10–22. [Google Scholar] [CrossRef]
- Palme, R.; Möstl, E. Measurement of cortisol metabolites in faeces of sheep as a parameter of cortisol concentration in blood. Z. Säugetierkd. Int. J. Mammal. Biol. 1997, 62, 192–197. [Google Scholar]
- Zhao, Z.-J.; Hambly, C.; Shi, L.-L.; Bi, Z.-Q.; Cao, J.; Speakman, J.R. Late lactation in small mammals is a critically sensitive window of vulnerability to elevated ambient temperature. Proc. Natl. Acad. Sci. USA 2020, 117, 24352–24358. [Google Scholar] [CrossRef] [PubMed]
- Hamid, H.Y.; Zakaria, M.Z.A.B.; Meng, G.Y.; Haron, A.W.; Mustapha, N.M. Effects of elevated ambient temperature on reproductive outcomes and offspring growth depend on exposure time. Sci. World J. 2012, 2012, 359134. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ohrnberger, S.A.; Monclus, R.; Rödel, H.G.; Valencak, T.G. Ambient temperature affects postnatal litter size reduction in golden hamsters. Front. Zool. 2016, 13, 51. [Google Scholar] [CrossRef] [PubMed]
- Ohrnberger, S.A.; Brinkmann, K.; Palme, R.; Valencak, T.G. Dorsal shaving affects concentrations of faecal cortisol metabolites in lactating golden hamsters. Naturwissenschaften 2018, 105, 13. [Google Scholar] [CrossRef]
- Ramin, M.; Denk, N.; Schenkel, J. The role of diet and housing-temperature in the production of genetically modified mouse embryos and their developmental capacity after cryopreservation. Theriogenology 2015, 84, 1306–1313. [Google Scholar] [CrossRef]
- Krol, E.; Speakman, J.R. Limits to sustained energy intake VI. Energetics of lactation in laboratory mice at thermoneutrality. J. Exp. Biol. 2003, 206, 4255–4266. [Google Scholar] [CrossRef]
- Yang, D.-B.; Li, L.; Wang, L.-P.; Chi, Q.-S.; Hambly, C.; Wang, D.-H.; Speakman, J.R. Limits to sustained energy intake. XIX. A test of the heat dissipation limitation hypothesis in Mongolian gerbils (Meriones unguiculatus). J. Exp. Biol. 2013, 216, 3358–3368. [Google Scholar] [CrossRef]
- Wen, J.; Tan, S.; Qiao, Q.-G.; Fan, W.-J.; Huang, Y.-X.; Cao, J.; Liu, J.-S.; Wang, Z.-X.; Zhao, Z.-J. Sustained energy intake in lactating Swiss mice: A dual modulation Process. J. Exp. Biol. 2017, 220, 2277–2286. [Google Scholar] [CrossRef]
- Valencak, T.G.; Wright, P.; Weir, A.; Mitchell, S.E.; Vaanholt, L.M.; Hambly, C.; Król, E.; Speakman, J.R. Limits to sustained energy intake. XXI. Effect of exposing the mother, but not her pups, to a cold environment during lactation in mice. J. Exp. Biol. 2013, 216, 4326–4333. [Google Scholar] [CrossRef]
- Zhao, Z.J.; Song, D.G.; Su, Z.C.; Wei, W.B.; Liu, X.B.; Speakman, J.R. Limits to sustained energy intake. XVIII. Energy intake and reproductive output during lactation in Swiss mice raising small litters. J. Exp. Biol. 2013, 216, 2349–2358. [Google Scholar] [CrossRef]
- Ohrnberger, S.A.; Hambly, C.; Speakman, J.R.; Valencak, T.G. Limits to sustained energy intake. XXXII. Hot again: Dorsal shaving increases energy intake and milk output in golden hamsters (Mesocricetus auratus). J. Exp. Biol. 2020, 223, jeb230383. [Google Scholar] [CrossRef] [PubMed]
- Król, E.; Murphy, M.; Speakman, J.R. Limits to sustained energy intake. X. Effects of fur removal on reproductive performance in laboratory mice. J. Exp. Biol. 2007, 210, 4233–4243. [Google Scholar] [CrossRef] [PubMed]
- Sadowska, J.; Gebczynski, A.K.; Lewoc, M.; Konarzewski, M. Not that hot after all: No limits to heat dissipation in lactating mice selected for high or low BMR. J. Exp. Biol. 2019, 222, jeb204669. [Google Scholar] [CrossRef] [PubMed]
- Krol, E.; Johnson, M.S.; Speakman, J.R. Limits to sustained energy intake VIII. Resting metabolic rate and organ morphology of laboratory mice lactating at thermoneutrality. J. Exp. Biol. 2003, 206, 4283–4291. [Google Scholar] [CrossRef][Green Version]
- Krol, E.; Speakman, J.R. Limits to sustained energy intake VII. Milk energy output in laboratory mice at thermoneutrality. J. Exp. Biol. 2003, 206, 4267–4281. [Google Scholar] [CrossRef]
- Zhao, Z.-J.; Li, L.; Yang, D.-B.; Chi, Q.-S.; Hambly, C.; Spaekman, J.R. Limits to sustained energy intake XXV: Milk energy output and thermogenesis in Swiss mice lactating at thermoneutrality. Sci. Rep. 2016, 6, 31626. [Google Scholar] [CrossRef]
- Harrison, G.A.; Morton, R.J.; Weiner, J.S. The growth in weight and tail length of inbred and hybrid mice reared at two different temperatures i. Growth in weight ii. Tail length. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1959, 242, 479–516. [Google Scholar]
- Skop, V.; Liu, N.; Guo, J.; Gavrilova, O.; Reitman, M.L. The contribution of the mouse tail to thermoregulation is modest. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E438–E446. [Google Scholar] [CrossRef]
- Möstl, E.; Palme, R. Hormones as indicators of stress. Dom. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef]
- Rabiee, A.R.; Dalley, D.; Borman, J.M.; Macmillan, K.L.; Schwarzenberger, F. Progesterone clearance rate in lactating dairy cows with two levels of dry matter and metabolisable energy intakes. Anim. Reprod. Sci. 2002, 72, 11–25. [Google Scholar] [CrossRef]
- Lepschy, M.; Touma, C.; Palme, R. Faecal glucocorticoid metabolites: How to express yourself–comparison of absolute amounts versus concentrations in samples from a study in laboratory rats. Lab. Anim. 2010, 44, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Kokolus, K.M.; Spangler, H.M.; Povinelli, B.J.; Farren, M.R.; Lee, K.P.; Repasky, E.A. Stressful presentations: Mild cold stress in laboratory mice influences phenotype of dendritic cells in naïve and tumor-bearing mice. Front. Immunol. 2014, 5, 23. [Google Scholar] [CrossRef]
- Leigh, N.D.; Kokolus, K.M.; O’Neill, R.E.; Du, W.; Eng, J.W.-L.; Qiu, J.; Chen, G.L.; McCarthy, P.L.; Farrar, J.D.; Cao, X.; et al. Housing temperature-induced stress is suppressing murine graft-versus-host disease through β2-adrenergic receptor signaling. J. Immunol. 2015, 195, 5045–5054. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.G.; Parrish, J.L.; Hughes, L.F.; Toth, L.A.; Caspary, D.M. Hearing in laboratory animals: Strain differences and nonauditory effects of noise. Comp. Med. 2005, 55, 12–23. [Google Scholar] [PubMed]
- Wersinger, S.R.; Martin, L.B. Optimization of laboratory conditions for the study of social behavior. ILAR J. 2009, 50, 64–80. [Google Scholar] [CrossRef]
- Castelhano-Carlos, M.J.; Baumans, V. The impact of light, noise, cage cleaning and in-house transport on welfare and stress of laboratory rats. Lab. Anim. 2009, 43, 311–327. [Google Scholar] [CrossRef]
- Mansfield, K.G.; Riley, L.K.; Kent, M.L. Detection, impact, and control of specific pathogens in animal resource facilities. ILAR J. 2010, 51, 171–179. [Google Scholar] [CrossRef]
- Gordon, C.J.; Puckett, E.T.; Repasky, E.S.; Johnstone, A.F.M. A device that allows rodents to behaviorally thermoregulate when housed in vivariums. JAALAS 2017, 56, 173–176. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| 20 °C | 25 °C | 30 °C | |
|---|---|---|---|
| B6N | 7/13 | 3/12 | 9/12 |
| B6D2F1 | 11/13 | 11/12 | 11/12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolbe, T.; Lassnig, C.; Poelzl, A.; Palme, R.; Auer, K.E.; Rülicke, T. Effect of Different Ambient Temperatures on Reproductive Outcome and Stress Level of Lactating Females in Two Mouse Strains. Animals 2022, 12, 2141. https://doi.org/10.3390/ani12162141
Kolbe T, Lassnig C, Poelzl A, Palme R, Auer KE, Rülicke T. Effect of Different Ambient Temperatures on Reproductive Outcome and Stress Level of Lactating Females in Two Mouse Strains. Animals. 2022; 12(16):2141. https://doi.org/10.3390/ani12162141
Chicago/Turabian StyleKolbe, Thomas, Caroline Lassnig, Andrea Poelzl, Rupert Palme, Kerstin E. Auer, and Thomas Rülicke. 2022. "Effect of Different Ambient Temperatures on Reproductive Outcome and Stress Level of Lactating Females in Two Mouse Strains" Animals 12, no. 16: 2141. https://doi.org/10.3390/ani12162141
APA StyleKolbe, T., Lassnig, C., Poelzl, A., Palme, R., Auer, K. E., & Rülicke, T. (2022). Effect of Different Ambient Temperatures on Reproductive Outcome and Stress Level of Lactating Females in Two Mouse Strains. Animals, 12(16), 2141. https://doi.org/10.3390/ani12162141