
Influences of Substrate Grain Size on the Burrowing Behavior of Juvenile Meretrix meretrix

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Acquisition of Experimental Animals and Substrates
2.2. Video Device
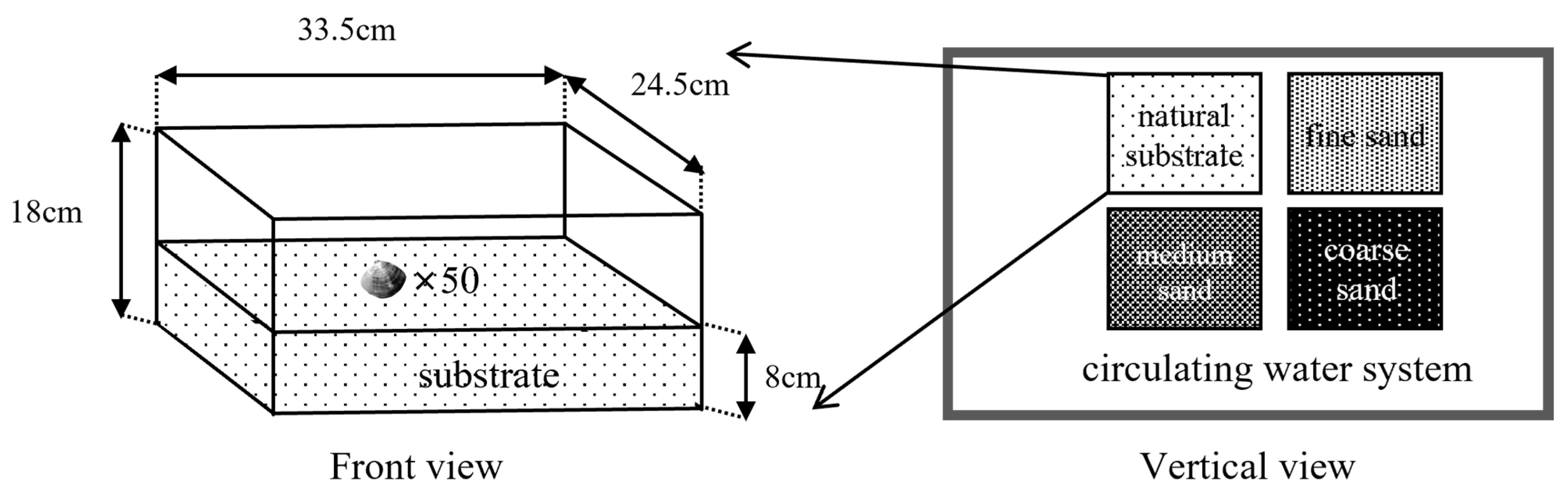
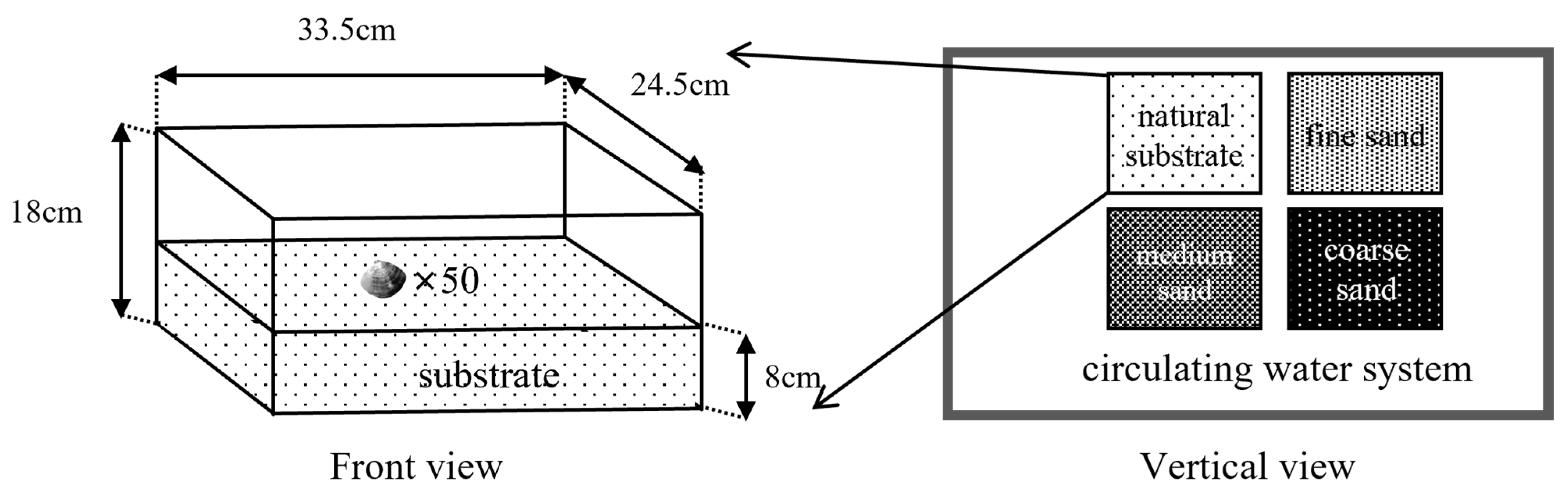
2.3. Experiment Ι
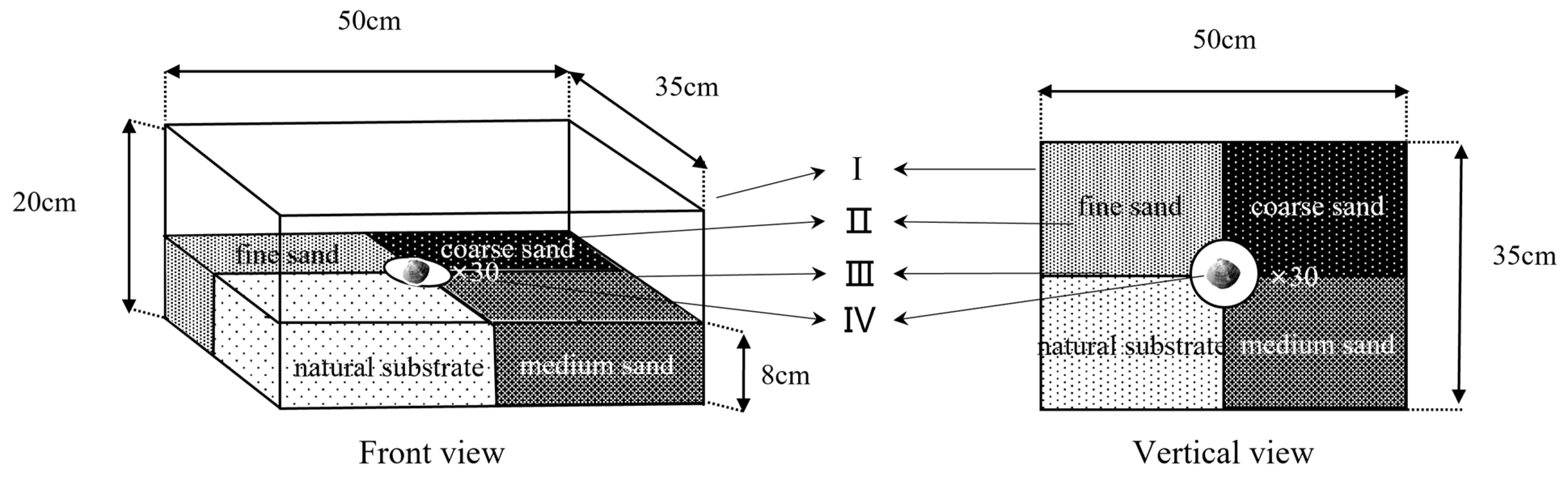
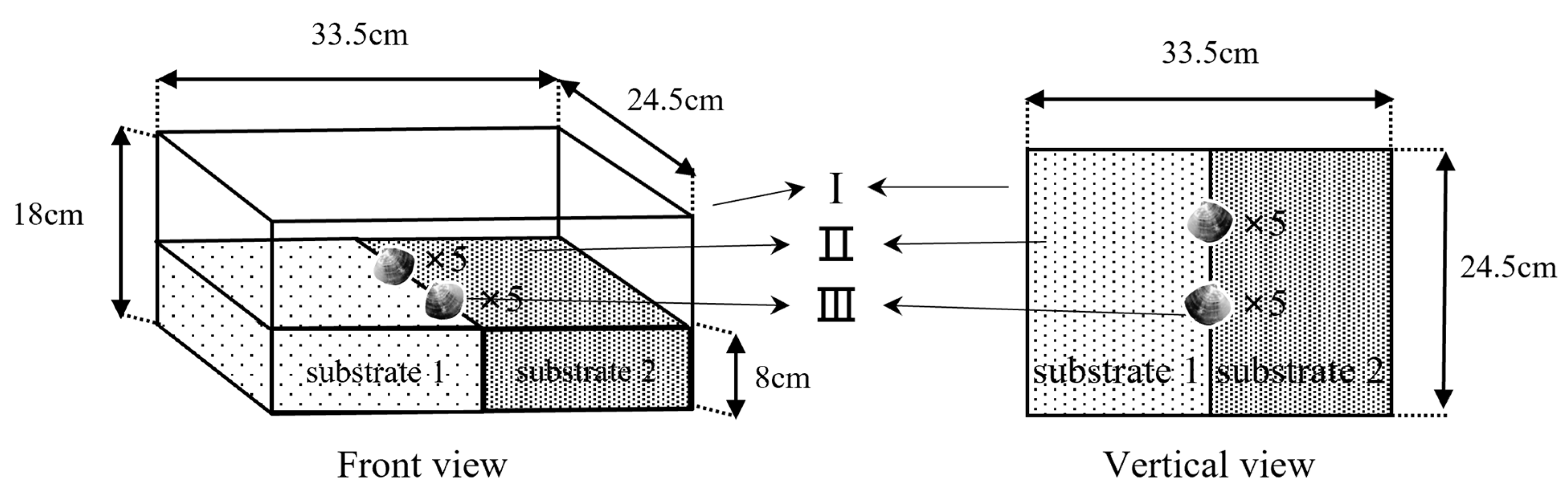
2.4. Experiment Ⅱ
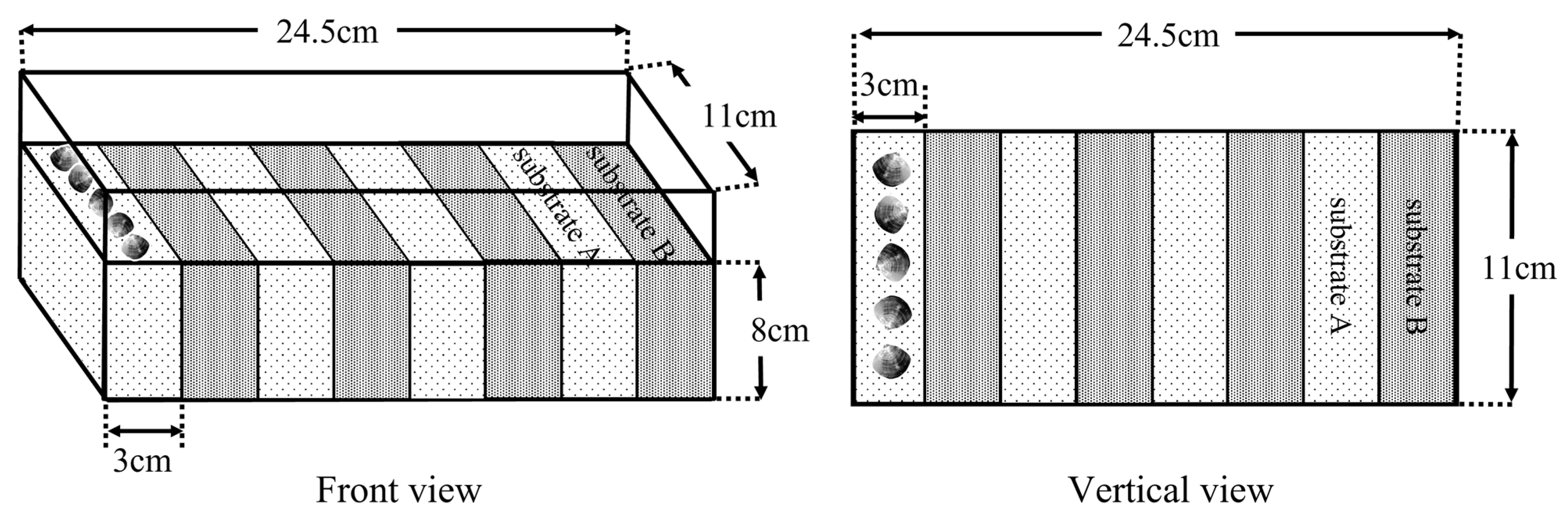
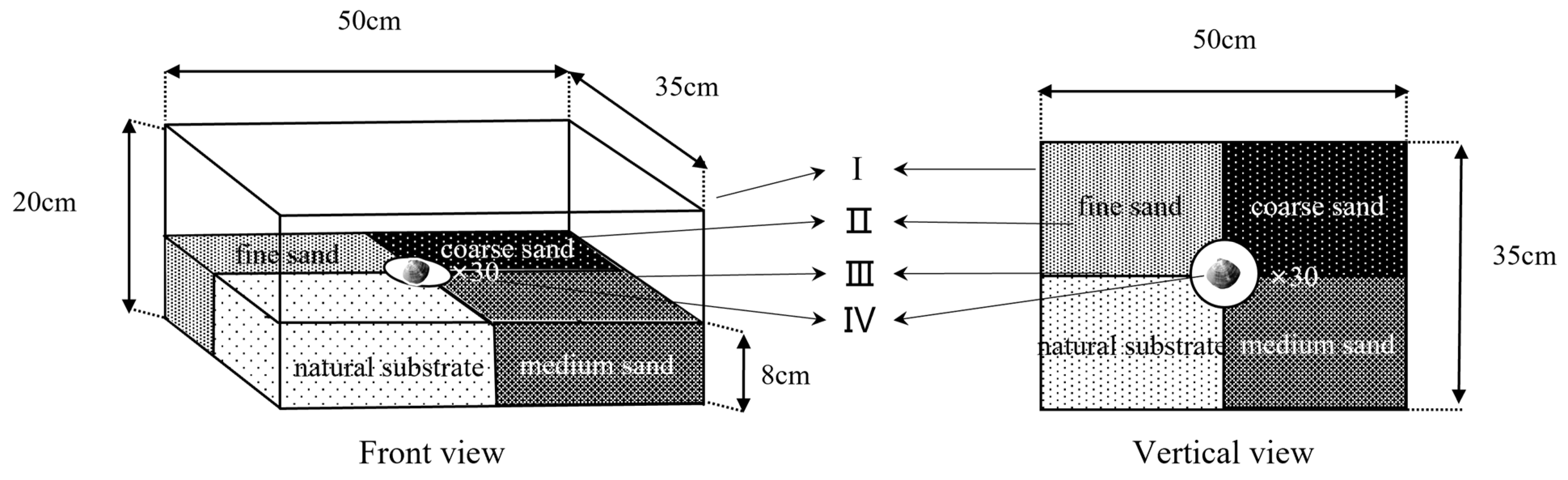
2.5. Experiment Ⅲ
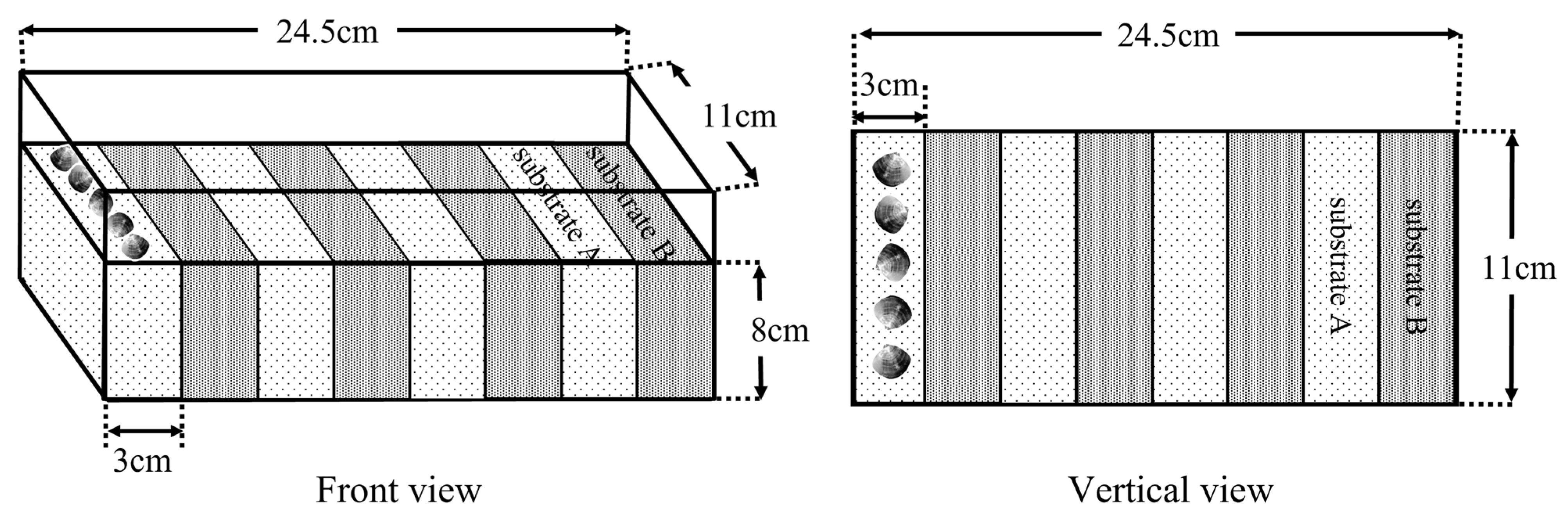
2.6. Experiment Ⅳ
2.7. Statistical Analyses
3. Results
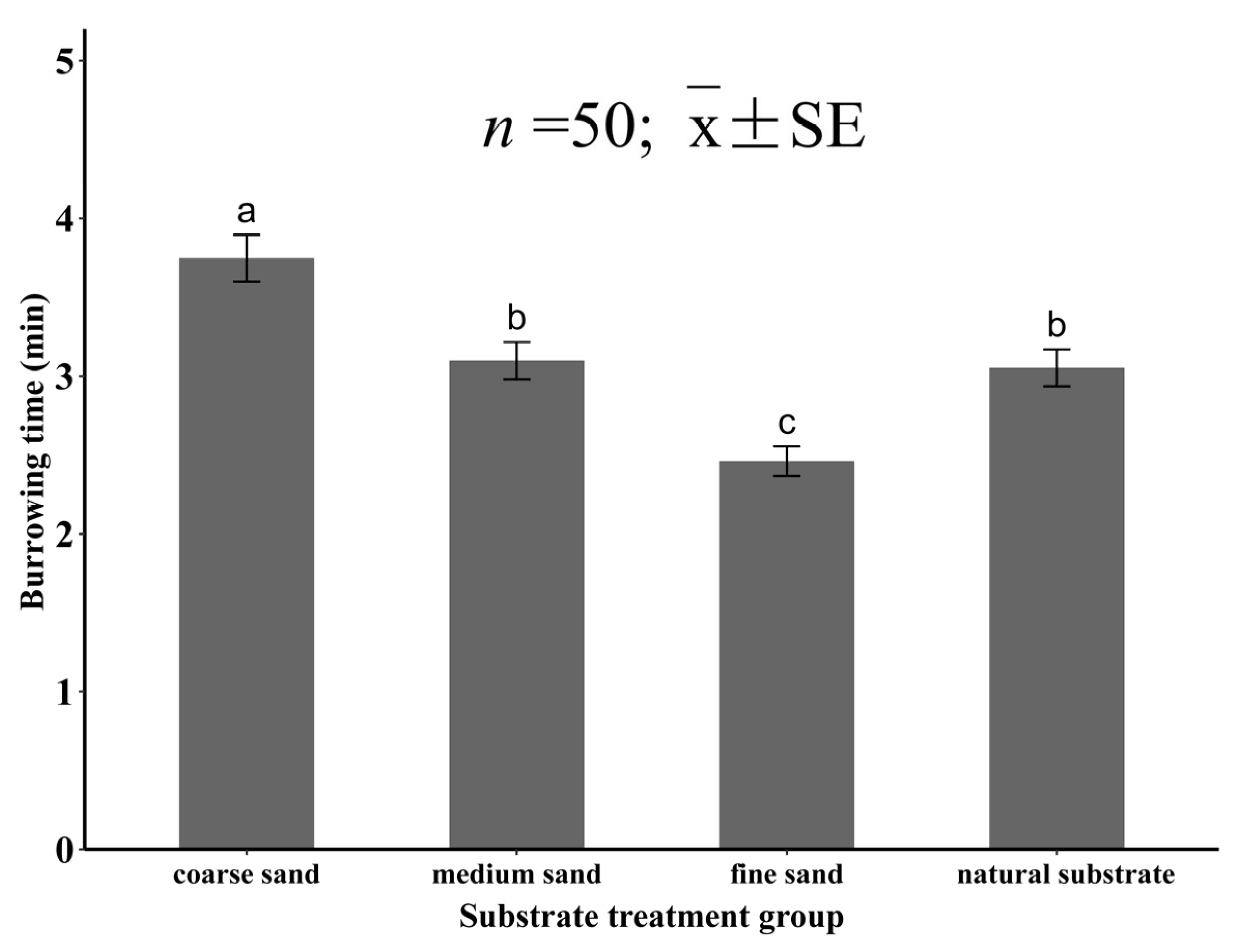
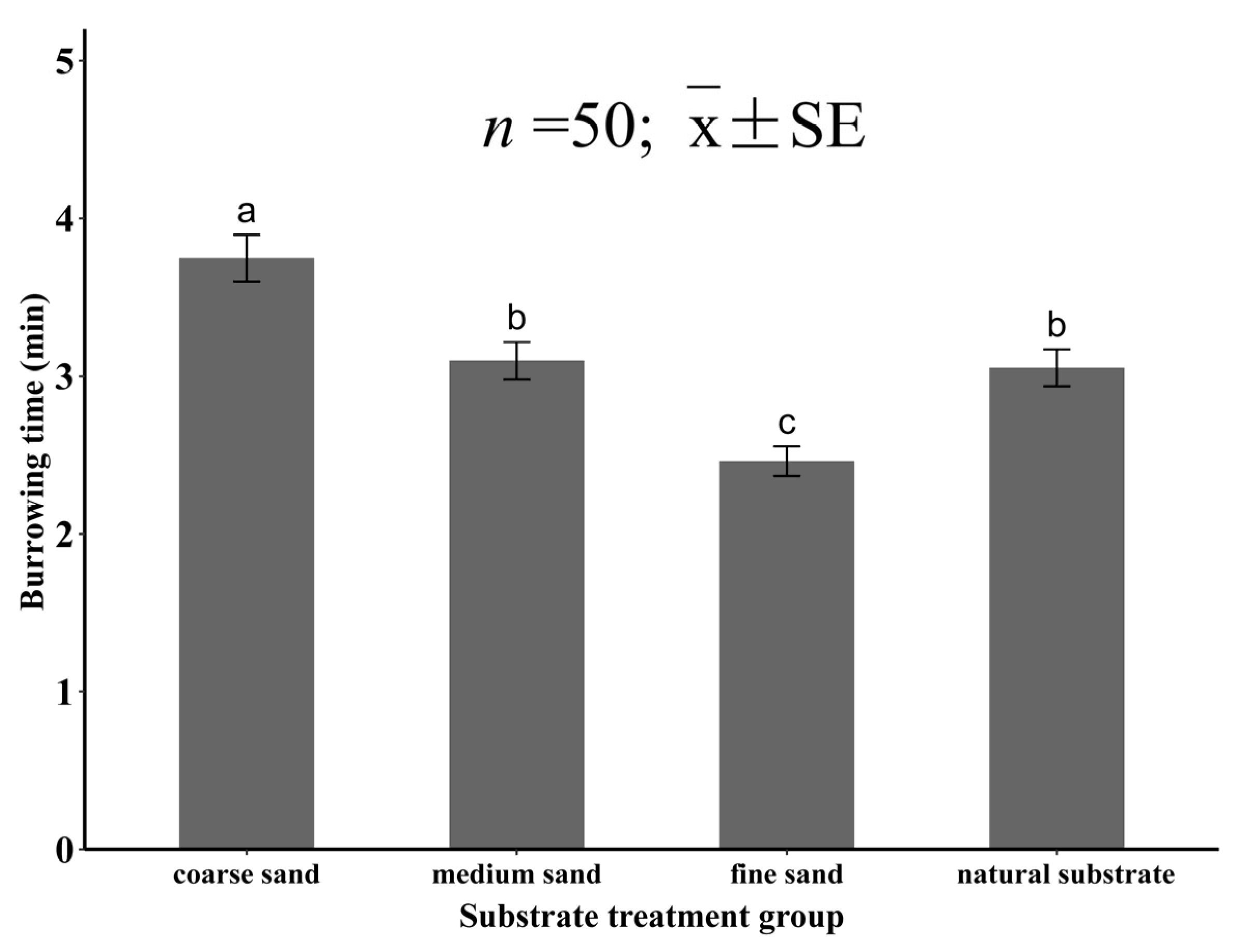
3.1. Burrowing Ability of Juvenile Meretrix meretrix in Different Grain Size Substrates
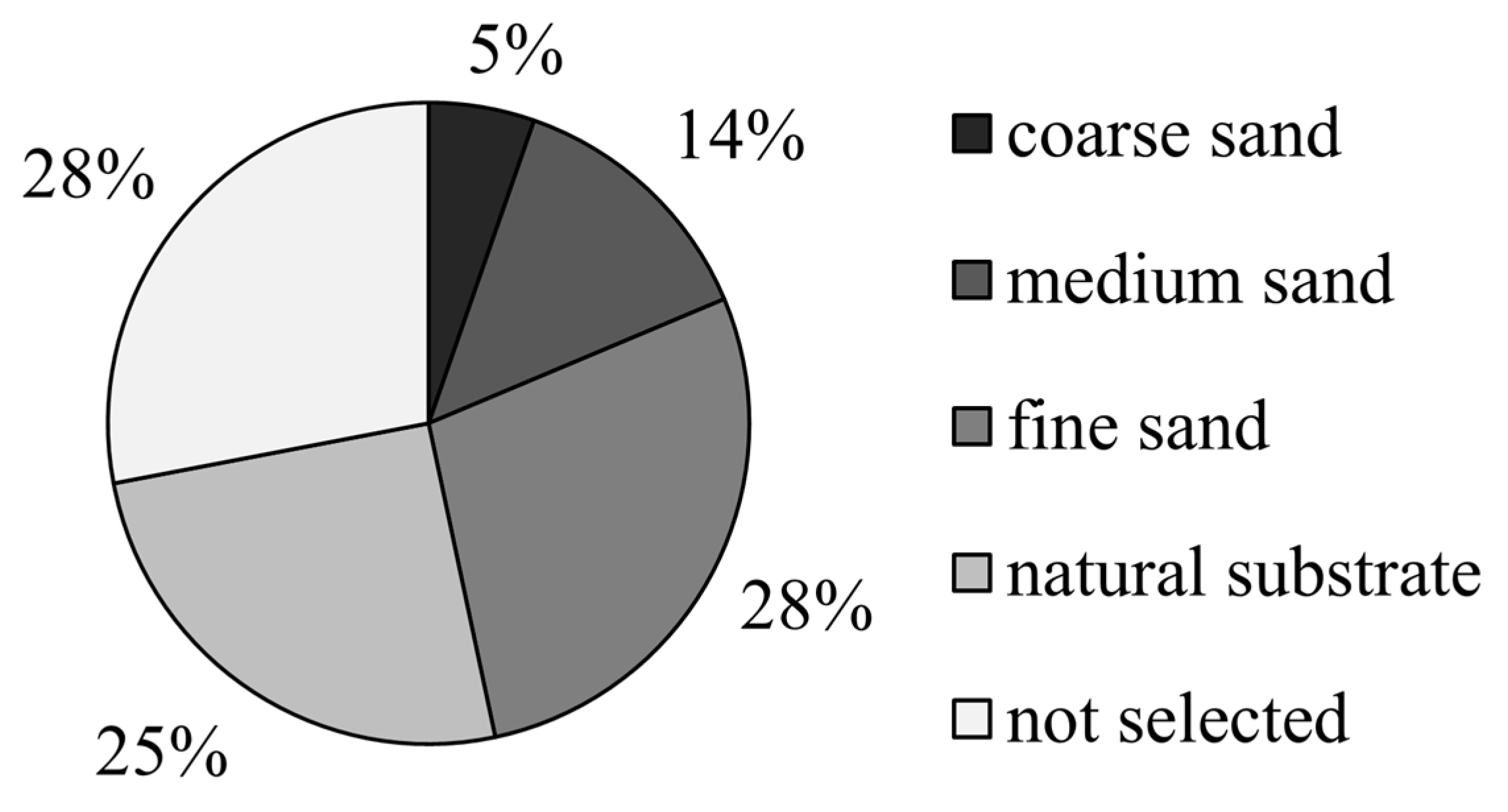
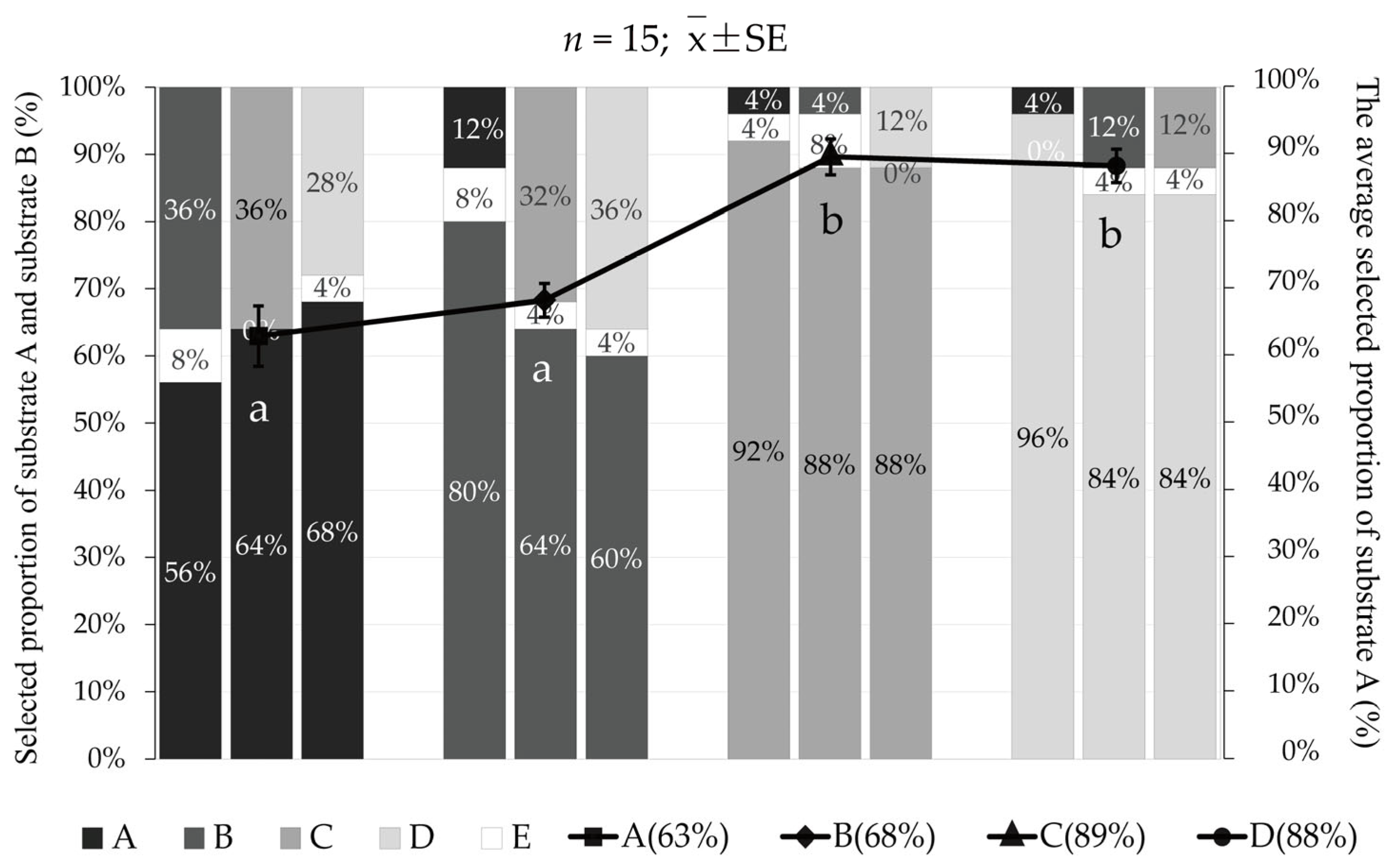
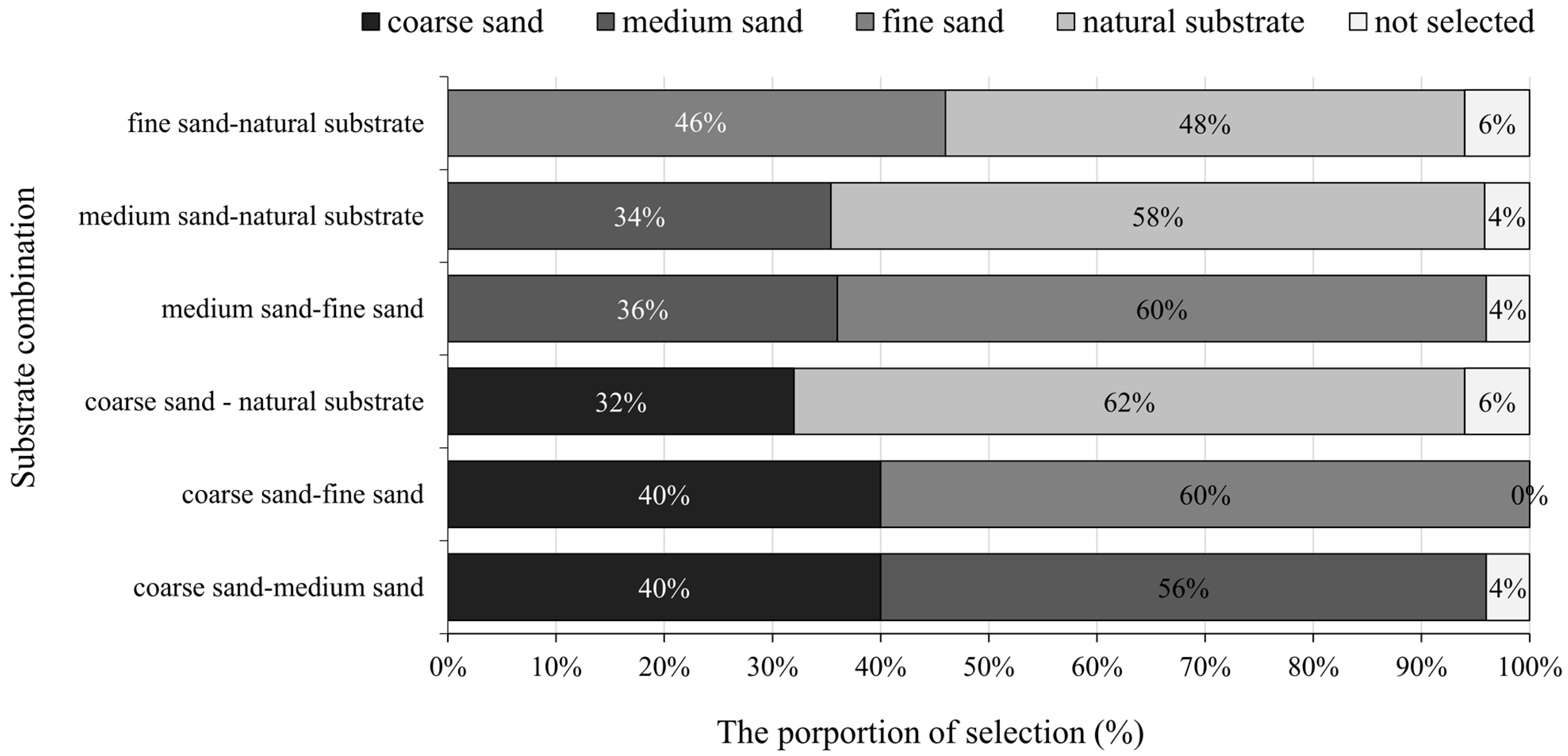
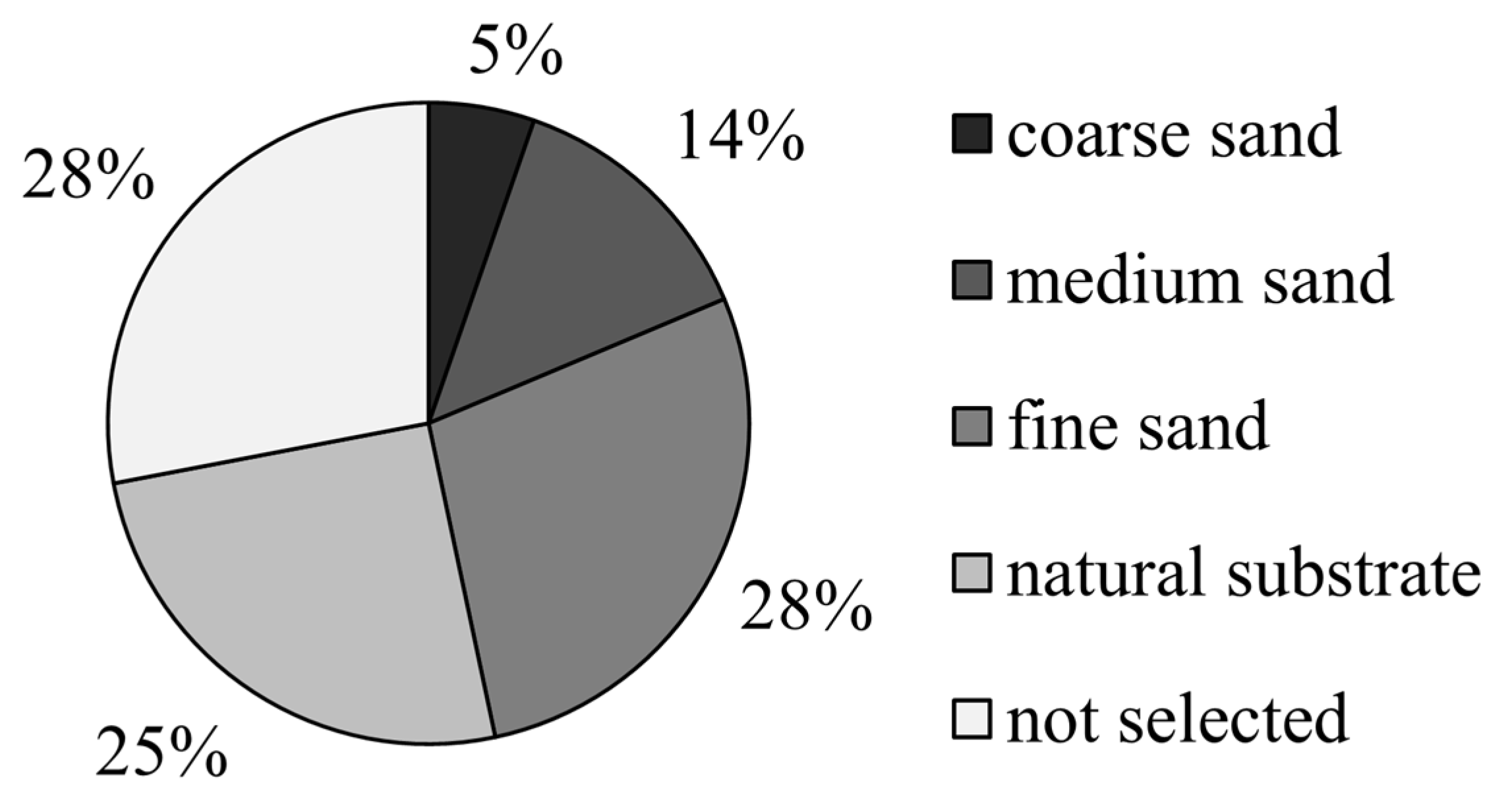
3.2. The Preference of the Juvenile Meretrix meretrix for Different Grain Size Substrates
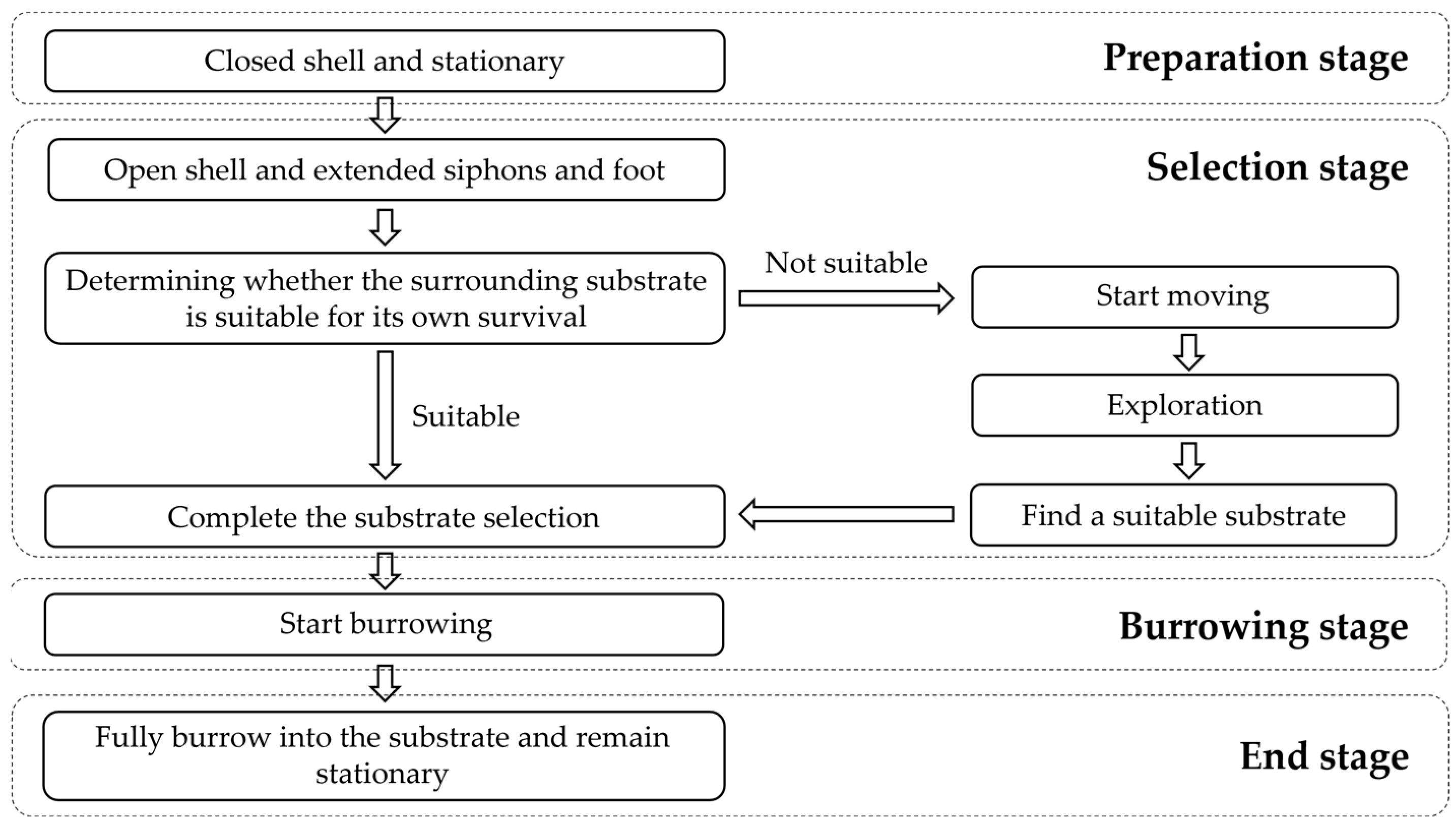
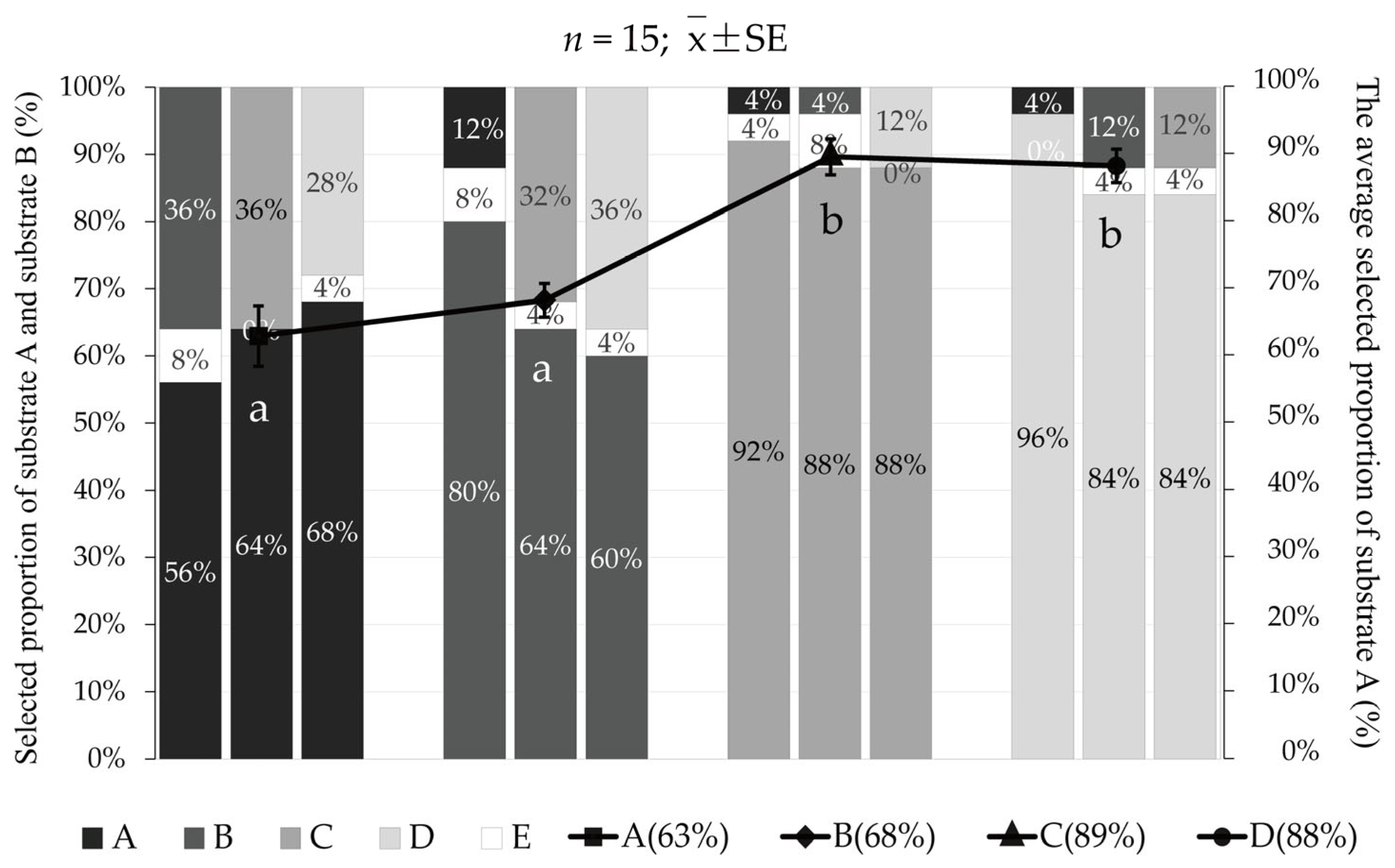
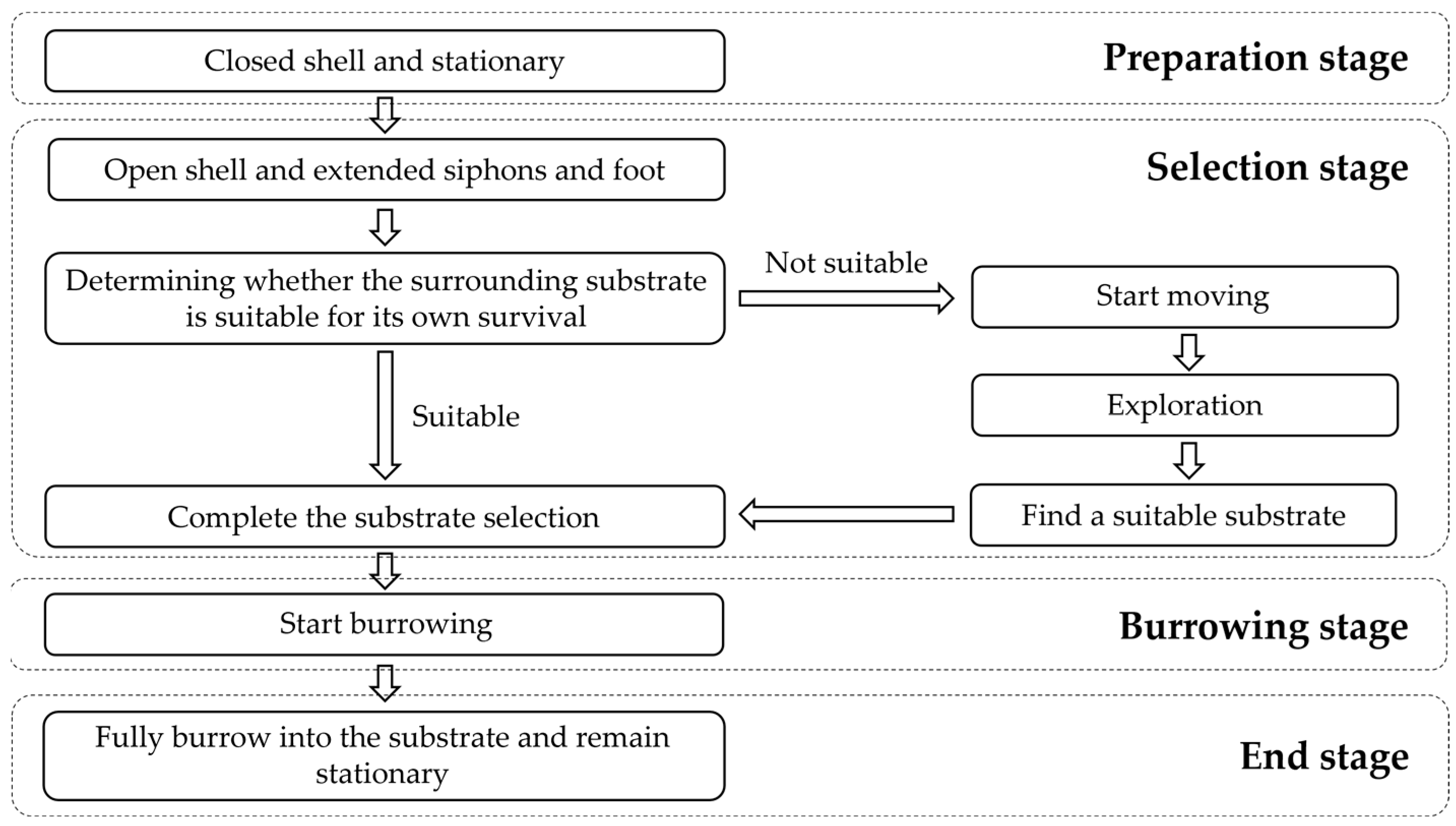
3.3. Behavioral Characteristics of Juvenile Meretrix meretrix in the Process of Substrate Selection
3.3.1. The Selection Process of the Substrate for Meretrix meretrix
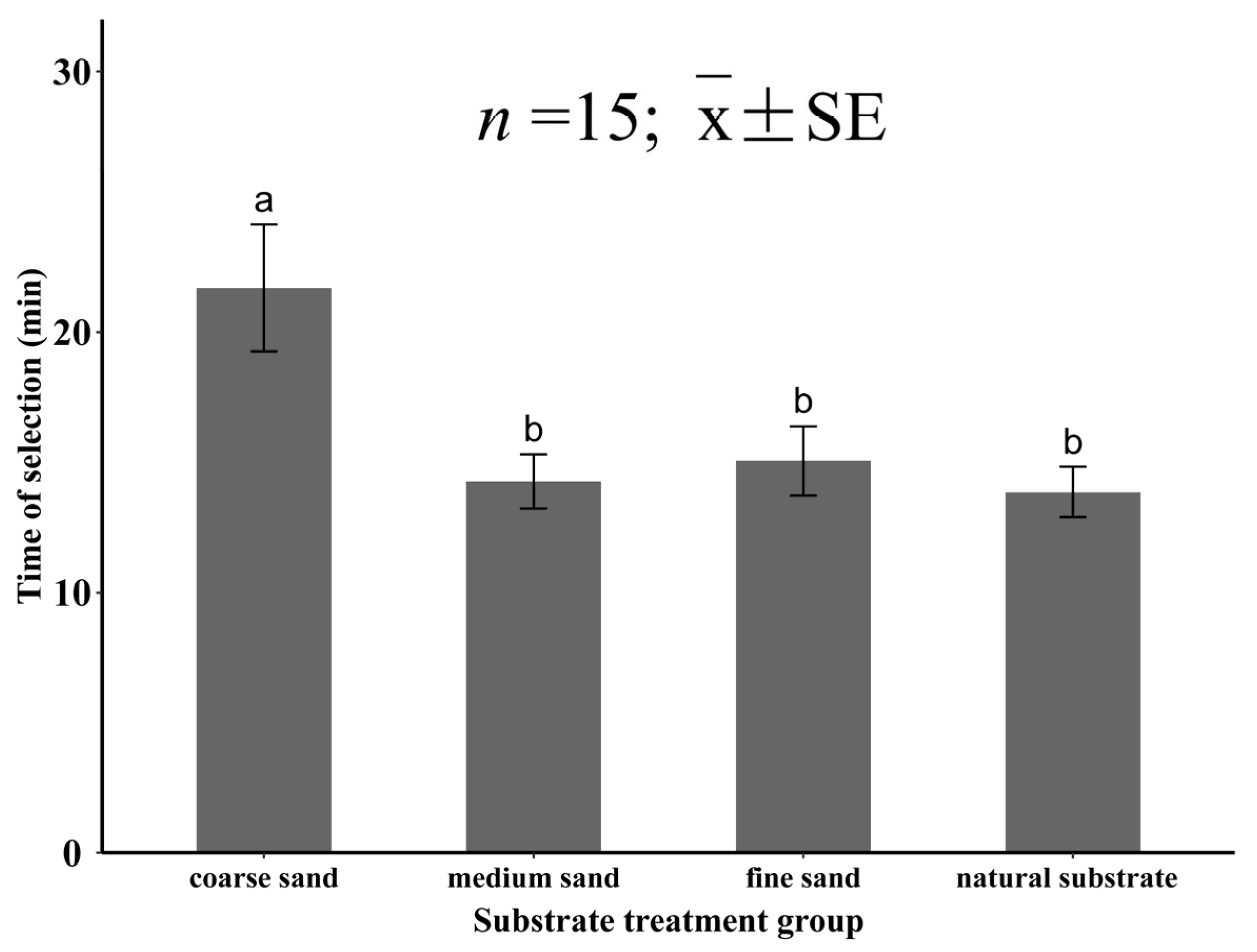
3.3.2. The Selection Time of Meretrix meretrix
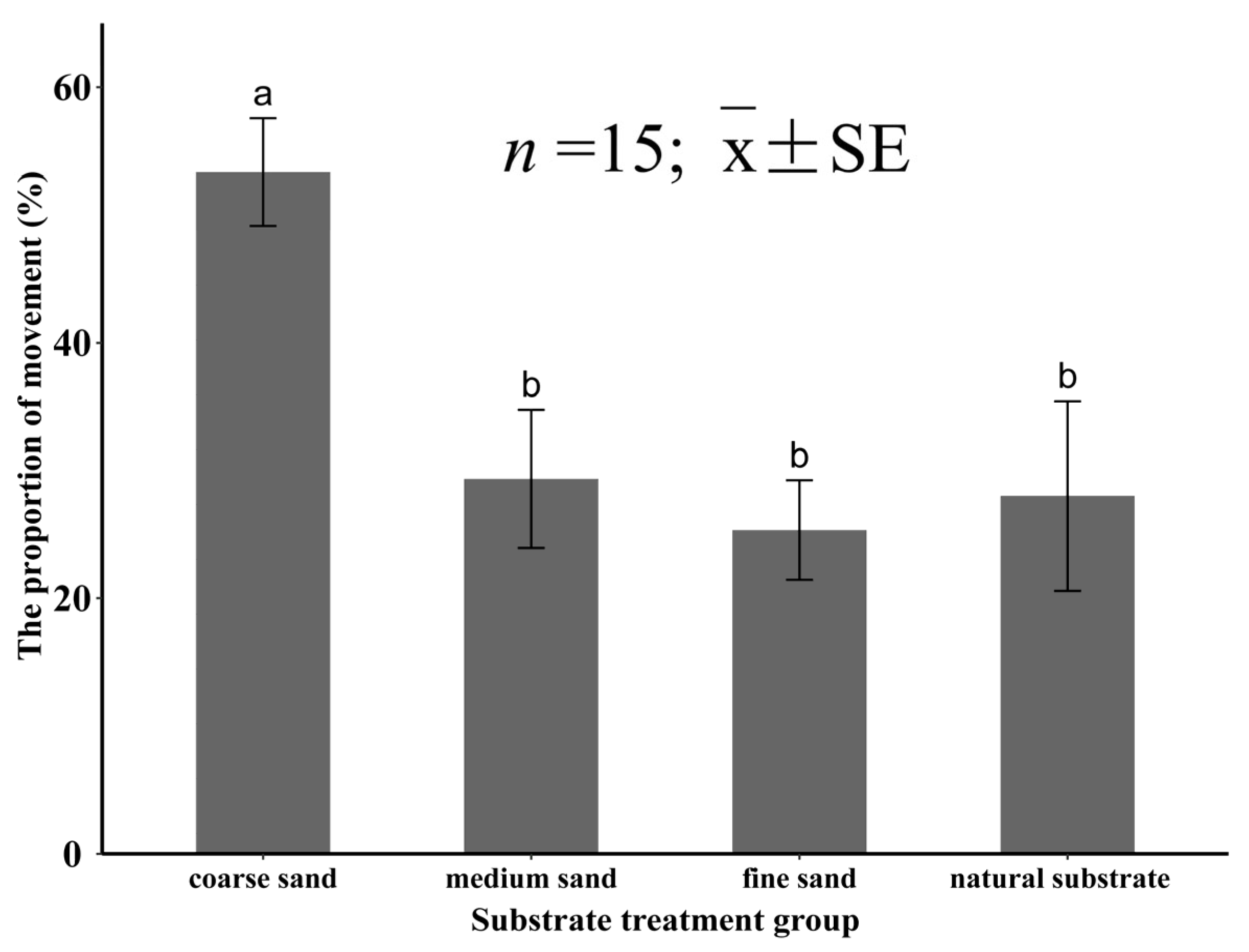
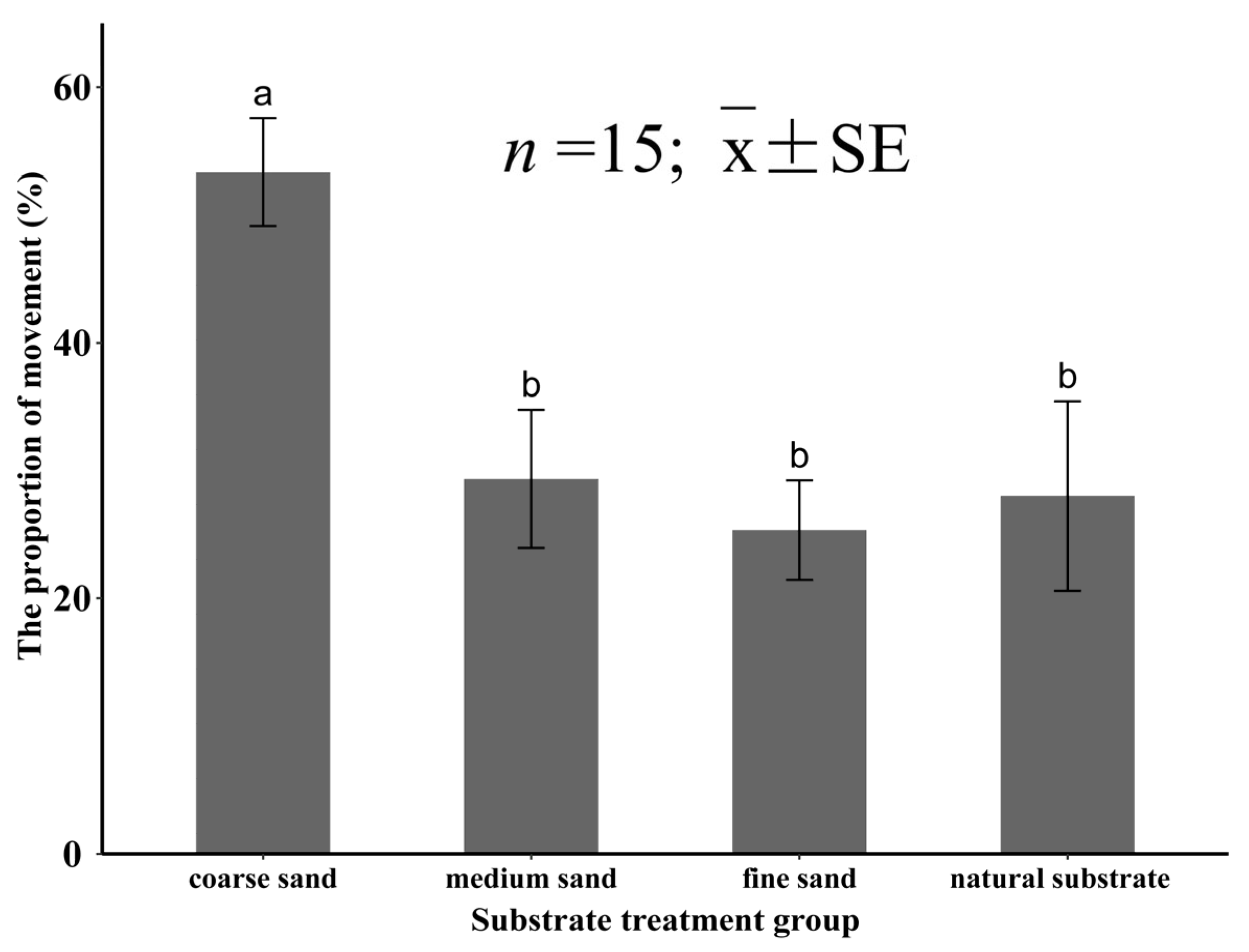
3.3.3. The Percentage of Movement for Meretrix meretrix
4. Discussion
4.1. Burrowing Ability
4.2. Substrate Preference
4.3. Behavioral Characteristics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zenone, A.; Badalamenti, F.; Giacalone, V.M.; Musco, L.; Pipitone, C.; Vega Fernández, T.; D’Anna, G. Substrate preference and settlement behaviour of the megalopa of the invasive crab Percnon gibbesi (Decapoda, Percnidae) in the Mediterranean Sea. Helgol. Mar. Res. 2016, 70, 1–7. [Google Scholar] [CrossRef]
- Zhang, X.; Li, Z.; Huo, Z.; Yan, X.; Yang, F.; Liu, H.; Zhang, X. Effect of substrate component on the growth and survival of juvenile sunray surf clam (Mactra chinensis Philippi). J. Ocean Univ. China 2016, 15, 676–680. [Google Scholar] [CrossRef]
- Nel, R.; McLachlan, A.; Winter, D.P.E. The effect of grain size on the burrowing of two Donax species. J. Exp. Mar. Biol. Ecol. 2001, 265, 219–238. [Google Scholar] [CrossRef]
- Moksnes, P.O.; Hedvall, O.; Reinwald, T. Settlement behavior in shore crabs Carcinus maenas: Why do postlarvae emigrate from nursery habitats? Mar. Ecol. Prog. Ser. 2003, 250, 215–230. [Google Scholar] [CrossRef]
- Yeager, M.M.; Cherry, D.S.; Neves, R.J. Feeding and Burrowing Behaviors of Juvenile Rainbow Mussels, Villosa iris (Bivalvia:Unionidae). J. N. Am. Benthol. Soc. 1994, 13, 217–222. [Google Scholar] [CrossRef]
- Joo, S.; Jo, K.; Bae, H.; Seo, H.; Kim, T. Optimal sediment grain size and sorting for survival and growth of juvenile Manila clams, Venerupis philippinarum. Aquaculture 2021, 543, 737010. [Google Scholar] [CrossRef]
- Martinez-Garcia, E.; Carlsson, M.S.; Sanchez-Jerez, P.; Sánchez-Lizaso, J.L.; Sanz-Lazaro, C.; Holmer, M. Effect of sediment grain size and bioturbation on decomposition of organic matter from aquaculture. Biogeochemistry 2015, 125, 133–148. [Google Scholar] [CrossRef]
- Meng, S.; Fang, J.; Jiang, Z.; Xu, Y.; Dong, S.; Yu, W.; Hou, X.; Gao, Y.; Du, M.; Lin, F. Experimental study of the choice behavior of Perinereis aibuhitensis Grube among different sediment types. Progress in Fishery Sciences. Prog. Fish. Sci. 2020, 41, 110–116. [Google Scholar] [CrossRef]
- Ueng, Y.-T.; Lin, F.-J.; Tsao, C.-W.; Chen, H.-C.; Chang, W.-C. Sand Grain Size and Organic Matter Concentration in the Intertidal Zones on the Southwest Coast of Taiwan. Nat. Resour. 2021, 12, 108–123. [Google Scholar] [CrossRef]
- Wong, M.C.; Peterson, C.H.; Kay, J. Prey size selection and bottom type influence multiple predator effects in a crab–bivalve system. Mar. Ecol. Prog. Ser. 2010, 409, 143–156. [Google Scholar] [CrossRef]
- de la Huz, R.; Lastra, M.; López, J. The influence of sediment grain size on burrowing, growth and metabolism of Donax trunculus L. (Bivalvia: Donacidae). J. Sea Res. 2002, 47, 85–95. [Google Scholar] [CrossRef]
- Jermacz, Ł.; Dzierżyńska, A.; Poznańska, M.; Kobak, J. Experimental evaluation of preferences of an invasive Ponto-Caspian gammarid Pontogammarus robustoides (Amphipoda, Gammaroidea) for mineral and plant substrata. Hydrobiologia 2014, 746, 209–221. [Google Scholar] [CrossRef]
- Pépino, M.; Rodríguez, M.A.; Magnan, P. Shifts in movement behavior of spawning fish under risk of predation by land-based consumers. Behav. Ecol. 2015, 26, 996–1004. [Google Scholar] [CrossRef]
- Zhou, S.; Zhang, X.; Liu, X.; Zhang, P. Substrate preference and burrowing ability assessment of the juvenile Scapharca broughtonii. J. Fish. China 2015, 39, 867–875. [Google Scholar] [CrossRef]
- Huehner, M.K. Field and Laboratory Determination of Substrate Preferences of Unionid Mussels. Ohio J. Sci. 1987, 87, 29–32. [Google Scholar]
- Downing, J.A.; Van Leeuwen, H.; Di Paolo, L.A. Substratum patch selection in the lacustrine mussels Elliptio complanata and Pyganodon grandis grandis. Freshwat. Biol. 2000, 44, 641–648. [Google Scholar] [CrossRef]
- Allen, D.C.; Vaughn, C.C. Burrowing behavior of freshwater mussels in experimentally manipulated communities. J. N. Am. Benthol. Soc. 2009, 28, 93–100. [Google Scholar] [CrossRef]
- Wang, X.; Song, L.; Chen, Y.; Ran, H.; Song, J. Impact of ocean acidification on the early development and escape behavior of marine medaka (Oryzias melastigma). Mar. Environ. Res. 2017, 131, 10–18. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, F.; Su, X.; Liu, D.; Dong, S. Size matters for predator-prey relationships: The case of swimming crab Portunus trituberculatus preying on Manila clam Ruditapes philippinarum. Aquaculture 2017, 479, 638–644. [Google Scholar] [CrossRef]
- Liu, D.; Guo, X.; Zhong, D.; Wang, F.; Li, L.; Hu, N. Prey density and a conspecific competitor influence multiple predator effects in a crab-clam foraging system. Aquaculture 2018, 493, 264–271. [Google Scholar] [CrossRef]
- Wang, R.; Wang, Z.; Zhang, J. Science of Marine Shellfish Culture; Qingdao Ocean University Press: Qingdao, China, 1993; pp. 396–410. [Google Scholar]
- Zhang, A.; Li, H.; Yang, X.; Wang, L.; Gao, Y.; Song, M.; Yuan, X. Stock Assessment of Hatchery-Released Clam Meretrix meretrix in an Estuary of China From the Perspectives of Population Ecology and Genetic Diversity. Front. Mar. Sci. 2021, 8, 725238. [Google Scholar] [CrossRef]
- Hashiguchi, M.; Yamaguchi, J.; Henmi, Y. Distribution and movement between habitats with growth of the hard clam Meretrix lusoria in the Shirakawa–Midorikawa estuary of the Ariake Sea. Fish. Sci. 2014, 80, 687–693. [Google Scholar] [CrossRef]
- Chen, L.; Wang, X.; Chen, D. Research on the development of hard clam market in China mainland. J. Fish. Univ. Shanghai 2004, 13, 283–287. [Google Scholar]
- Ji, B. Analysis of the cost benefit of exporting marine aquaculture products from Wenzhou with Meretrix meretrix as an example. J. Aquac. 2014, 35, 37–40. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, A.; Li, X.; Mao, Y. Spatial distribution of buried molluscs and their relationship with sediment factors in Geligang. Mar. Sci. 2016, 40, 32–39. [Google Scholar] [CrossRef]
- Lü, H.; Chapelsky, A.; Fu, M.; Xi, D.; Zhang, Z.; Zhang, X. Effect of sand grain size on substrate preference and burial behaviour in cultured Japanese flounder juvenile, Paralichthys olivaceus. Aquacult. Res. 2018, 49, 1664–1671. [Google Scholar] [CrossRef]
- Zhang, A.; Yuan, X.; Yang, F.; Wang, H.; Wang, L.; Zhao, K. Effects of temperature, salinity and sediment on the burrowing behavior of clam Meretrix meretrix. Chin. J. Ecol. 2015, 34, 1595–1601. [Google Scholar] [CrossRef]
- Sakakura, Y. Larval fish behavior can be a predictable indicator for the quality of Japanese flounder seedlings for release. Aquaculture 2006, 257, 316–320. [Google Scholar] [CrossRef]
- Chang, C.C.; Huang, J.F.; Schafferer, C.; Lee, J.M.; Ho, L.M. Impacts of culture survival rate on culture cost and input factors: Case study of the hard clam (Meretrix meretrix) culture in Yunlin County, Taiwan. J. World Aquacult. Soc. 2019, 51, 139–158. [Google Scholar] [CrossRef]
- Henmi, Y.; Kobayashi, S.; Yamaguchi, J.; Hashiguchi, M. Recruitment and movement of the hard clam Meretrix lusoria in a tidal river of northern Kyushu, Japan. Fish. Sci. 2014, 80, 705–714. [Google Scholar] [CrossRef]
- Zhang, A.; Li, T.; Su, X.; Liu, B. Current status and prospect of Meretrix meretrix culture. Fish. Sci. 2005, 24, 31–33. [Google Scholar] [CrossRef]
- Doeglas, D.J. Grain-Size Indices, Classification and Environment. Sedimentology 1968, 10, 83–100. [Google Scholar] [CrossRef]
- Liu, D.; Wang, F.; Lu, Y.; Hu, N.; Su, X.; Liu, J.; Zhu, B. Behavioral mechanisms of swimming crab (Portunus trituberculatus) preying on Manila clams (Ruditapes philippinarum): Effects of substrate and competition. Mar. Biol. 2019, 166, 15. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, F.; Yang, C.; Liu, D.; Wang, X.; Su, X. Predator size influences intraspecific multiple predator effects in swimming crab-Manila clam system. Aquaculture 2018, 488, 74–79. [Google Scholar] [CrossRef]
- Alexander, R.R.; Stanton, R.J.; Dodd, J.R. Influence of Sediment Grain Size on the Burrowing of Bivalves: Correlation with Distribution and Stratigraphic Persistence of Selected Neogene Clams. Palaios 1993, 8, 289–303. [Google Scholar] [CrossRef]
- Kranz, P.M. The Anastrophic Burial of Bivalves and its Paleoecological Significance. J. Geol. 1974, 82, 237–265. [Google Scholar] [CrossRef]
- Watters, G.T.; O’Dee, S.H.; Chordas, S. Patterns of Vertical Migration in Freshwater Mussels (Bivalvia: Unionoida). J. Freshwat. Ecol. 2011, 16, 541–549. [Google Scholar] [CrossRef]
- Sun, T.; Liu, C.e.; Li, X.; An, D.; Yu, H.; Ma, Z.; Liu, F. The effect of substrate grain size on burrowing ability and distribution characteristics of Perinereis aibuhitensis. Acta Oceanol. Sin. 2019, 38, 52–58. [Google Scholar] [CrossRef]
- Fiori, S.M.; Carcedo, M.C. Influence of Grain Size on Burrowing and Alongshore Distribution of the Yellow Clam (Amarilladesma mactroides). J. Shellfish Res. 2015, 34, 785–789. [Google Scholar] [CrossRef]
- Lewis, J.B.; Riebel, P.N. The effect of substrate on burrowing in freshwater mussels (Unionidae). Can. J. Zool. 1984, 62, 2023–2025. [Google Scholar] [CrossRef]
- Hyvärinen, H.; Saarinen-Valta, M.; Mäenpää, E.; Taskinen, J. Effect of substrate particle size on burrowing of the juvenile freshwater pearl mussel Margaritifera margaritifera. Hydrobiologia 2021, 848, 1137–1146. [Google Scholar] [CrossRef]
- McLachlan, A.; Jaramillo, E.; Defeo, O.; Dugan, J.; de Ruyck, A.; Coetzee, P. Adaptations of bivalves to different beach types. J. Exp. Mar. Biol. Ecol. 1995, 187, 147–160. [Google Scholar] [CrossRef]
- Morton, B.; Leung, S.F.; Leung, K.F. The biology and functional morphology of Meretrix cf. meretrix (Bivalvia: Veneridae: Meretricinae) at Tong Fuk Miu Wan, Shui Hau, Lantau Island, Hong Kong. Reg. Stud. Mar. Sci. 2021, 45, 101842. [Google Scholar] [CrossRef]
- Enderlein, P. Effect of substrate composition on burrowing depth and respiratory current in two spatangoids (Echinoidea). Sarsia 2007, 89, 190–195. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, F.; Hu, N.; Su, X.; Liu, Z.; Dircksen, H. Prey selection of the swimming crab Portunus trituberculatus (Miers, 1876) (Brachyura: Portunidae) foraging on bivalves. J. Crust. Biol. 2017, 37, 521–528. [Google Scholar] [CrossRef]
- Thach, T.T.; Harada, M.; Oniki, A.; Hiramatsu, K.; Tabata, T. Experimental study on the influence of dissolved organic matter in water and redox state of bottom sediment on water quality dynamics under anaerobic conditions in an organically polluted water body. Paddy Water Environ. 2017, 15, 889–906. [Google Scholar] [CrossRef]
- Taskinen, J.; Saarinen, M. Burrowing behaviour affects Paraergasilus rylovi abundance in Anodonta piscinalis. Parasitology 2006, 133, 623–629. [Google Scholar] [CrossRef]
- Nichols, S.J.; Wilcox, D.A. Burrowing saves Lake Erie clams. Nature 1997, 389, 921. [Google Scholar] [CrossRef]
- Haider, F.; Sokolov, E.P.; Sokolova, I.M. Effects of mechanical disturbance and salinity stress on bioenergetics and burrowing behavior of the soft-shell clam Mya arenaria. J. Exp. Biol. 2018, 221, 172643. [Google Scholar] [CrossRef]
- Sun, J.; Hamel, J.-F.; Stuckless, B.; Small, T.J.; Mercier, A. Effect of light, phytoplankton, substrate types and colour on locomotion, feeding behaviour and microhabitat selection in the sea cucumber Cucumaria frondosa. Aquaculture 2020, 526, 735369. [Google Scholar] [CrossRef]
- Zajac, K.; Zajac, T. The role of active individual movement in habitat selection in the endangered freshwater mussel Unio crassus Philipsson 1788. J. Conchol. 2011, 40, 446–461. [Google Scholar]
- Spyksma, A.J.; Taylor, R.B.; Shears, N.T. Predation cues rather than resource availability promote cryptic behaviour in a habitat-forming sea urchin. Oecologia 2017, 183, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Shinya, T.; Yasuhisa, H. Migration by Mucous Cord in the Hard Clam Meretrix lusoria. Jpn. J. Benthol. 2011, 65, 76–81. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grain Group Type | Fine Gravel | Coarse Sand | Medium Sand | Fine Sand | Silt | Mud |
|---|---|---|---|---|---|---|
| Particle diameter (μm) | >2000 | 2000–500 | 500–250 | 250–63 | 63–4 | 4–0 |
| Volume proportion (%) | 0.00 | 0.63 | 1.58 | 91.72 | 4.69 | 1.49 |
| Median particle size | 127 μm | |||||
| Substrate Name | Combination | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Substrate A | a | a | a | b | b | b | c | c | c | d | d | d |
| Substrate B | b | c | d | a | c | d | a | b | d | a | b | c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Xue, S.; Li, J.; Fang, J.; Liu, L.; Ma, Z.; Yu, W.; Zhuang, H.; Mao, Y. Influences of Substrate Grain Size on the Burrowing Behavior of Juvenile Meretrix meretrix. Animals 2022, 12, 2094. https://doi.org/10.3390/ani12162094
Zhang C, Xue S, Li J, Fang J, Liu L, Ma Z, Yu W, Zhuang H, Mao Y. Influences of Substrate Grain Size on the Burrowing Behavior of Juvenile Meretrix meretrix. Animals. 2022; 12(16):2094. https://doi.org/10.3390/ani12162094
Chicago/Turabian StyleZhang, Changsheng, Suyan Xue, Jiaqi Li, Jinghui Fang, Lulei Liu, Zhanfei Ma, Wenhan Yu, Haonan Zhuang, and Yuze Mao. 2022. "Influences of Substrate Grain Size on the Burrowing Behavior of Juvenile Meretrix meretrix" Animals 12, no. 16: 2094. https://doi.org/10.3390/ani12162094
APA StyleZhang, C., Xue, S., Li, J., Fang, J., Liu, L., Ma, Z., Yu, W., Zhuang, H., & Mao, Y. (2022). Influences of Substrate Grain Size on the Burrowing Behavior of Juvenile Meretrix meretrix. Animals, 12(16), 2094. https://doi.org/10.3390/ani12162094