
The Economic Burden of Chromosome Translocations and the Benefits of Enhanced Screening for Cattle Breeding

 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cytogenetic Preparation
2.2. Fluorescent In situ Hybridization
2.3. Calculations of the Benefits of Chromosome Screening
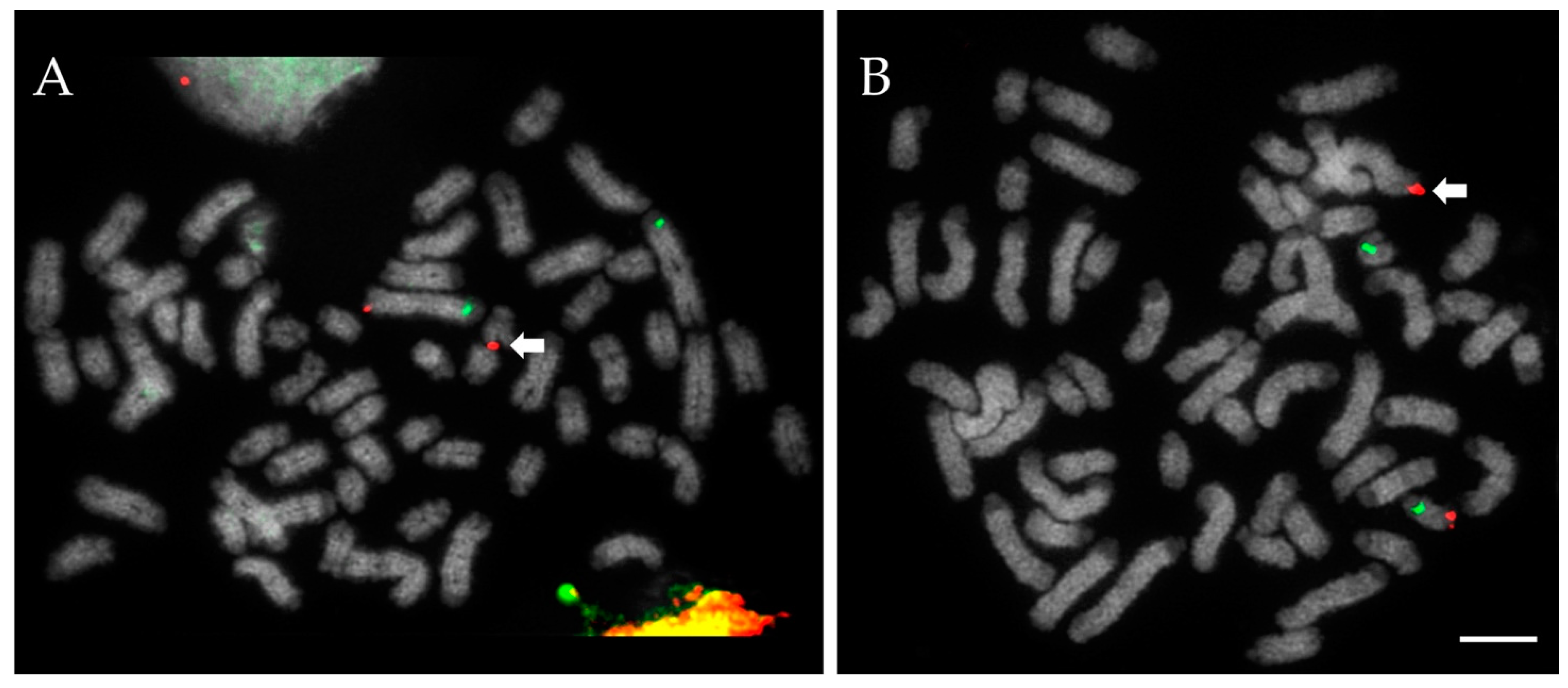
3. Results
3.1. Chromosome Screening
3.2. Financial Effects of an RT on a Herd

{kind=link}
| Conception Rate | Successful Live Births per Year a | Cows not Impregnated and Thus Not Lactating per Year a | Days per Year without Milk b | Opportunity Cost p.a. (Expressed as Value of Milk Lost GBP) c | Opportunity Cost p.a. (Expressed as Value of Calf That Could Be Sold GBP) d | Total Opportunity Cost per Year from Days without Milk (GBP) e | |
|---|---|---|---|---|---|---|---|
| Without RT | 55% | 12,650 | 10,350 | 569,250 | 3,073,950.00 | 358,627.50 | 3,432,577.50 |
| With an RT | 50% | 11,500 | 11,500 | 632,500 | 3,415,500.00 | 389,475.00 | 3,813,975.00 |
| 45% | 10,350 | 12,650 | 695,250 | 4,195,372.00 | 438,322.50 | 4,195,372.50 | |
| Difference (i.e., RT cost) | 5% f | 1150 | 1150 | 63,250 | 339,550.00 | 41,847.50 | 381,397.50 |
| 10% g | 2300 | 2300 | 126,000 | 1,121,422.00 | 79,695.00 | 1,201,117.00 |
3.3. Relative Incidence of RobTs vs. RTs
- The reported incidence that RECTs are 4Õ more common than non-1:29 RobTs but bearing in mind that it is estimated that around 86% have remained undetected [11].
- The fact that no screening programme would have actively removed RECTs from a breeding herd in the way that they have been near eliminated for RobT 1:29 in Holstein cattle [12].
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ritchie, H.; Roser, M. Meat and Dairy Production. Our World Data 2017. Available online: https://ourworldindata.org/meat-production (accessed on 10 January 2020).
- Simm, G.; Pollott, G.; Mrode, R.A.; Houston, R.; Marshall, K. Genetic Improvement of Farmed Animals; CABI: Wallingford, UK; Boston, MA, USA, 2020. [Google Scholar]
- Jennings, R.L.; Griffin, D.K.; O’Connor, R.E. A new Approach for Accurate Detection of Chromosome Rearrangements that Affect Fertility in Cattle. Animals 2020, 10, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, E.M.; Kelly, A.K.; O’Meara, C.; Eivers, B.; Lonergan, P.; Fair, S. Influence of bull age, ejaculate number, and season of collection on semen production and sperm motility parameters in Holstein Friesian bulls in a commercial artificial insemination centre. J. Anim. Sci. 2018, 96, 2408–2418. [Google Scholar] [CrossRef] [PubMed]
- Balsom, A. 4 Reasons to Use Artificial Insemination in Beef Herds; Farmers Weekly; MA Agriculture Ltd.: London, UK, 2020; Available online: https://www.fwi.co.uk/livestock/livestock-breeding/4-reasons-to-use-artificial-insemination-in-beef-herds (accessed on 1 April 2022).
- Short, W. In Pictures: Behind the Scenes at a Dairy Bull Stud; Farmers Weekly; MA Agriculture Ltd.: London, UK, 2015; Available online: https://www.fwi.co.uk/livestock/livestock-breeding/pictures-behind-scenes-dairy-bull-stud (accessed on 25 April 2022).
- O’Connor, R.E.; Kiazim, L.G.; Rathje, C.C.; Jennings, R.L.; Griffin, D.K. Rapid Multi-Hybridisation FISH Screening for Balanced Porcine Reciprocal Translocations Suggests a Much Higher Abnormality Rate Than Previously Appreciated. Cells 2021, 10, 250. [Google Scholar] [CrossRef] [PubMed]
- Lewis, N.M.; Rathje, C.C.; Canedo-Ribeiro, C.; Bosman, L.M.; Kiazim, L.G.; Jennings, R.L.; O’Connor, R.E.; Silvestri, G.; Griffin, D.K. Incidence, Reproductive Outcome, and Economic Impact of Reciprocal Translocations in the Domestic Pig. DNA 2021, 1, 68–76. [Google Scholar] [CrossRef]
- Garrick, D.J.; Garrick, D.J.; Ruvinsky, A.; CABI. The Genetics of Cattle, 2nd ed.; CABI: Wallingford, UK, 2014. [Google Scholar]
- Holečková, B.; Schwarzbacherová, V.; Galdíková, M.; Koleničová, S.; Halušková, J.; Staničová, J.; Verebová, V.; Jutková, A. Chromosomal Aberrations in Cattle. Genes 2021, 12, 1330. [Google Scholar] [CrossRef] [PubMed]
- Ogur, C.; Griffin, D.K. Preimplantation Genetic Testing for Structural Rearrangements. In Preimplantation Genetic Testing; Taylor and Francis: Abingdon-on-Thames, UK, 2020; pp. 49–76. [Google Scholar]
- De Lorenzi, L.; Morando, P.; Planas, J.; Zannotti, M.; Molteni, L.; Parma, P. Reciprocal translocations in cattle: Frequency estimation. J. Anim. Breed. Genet. 2012, 129, 409–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udroiu, I.; Sgura, A. Cytogenetic tests for animal production: State of the art and perspectives. Anim. Genet. 2017, 48, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.P.; Evans, R.D. The response to genetic merit for milk production in dairy cows differs by cow body weight. JDS Commun. 2021, 3, 32–37. [Google Scholar] [CrossRef]
- Bonnet-Garnier, A.; Lacaze, S.; Beckers, J.F.; Berland, H.M.; Pinton, A.; Yerle, M.; Ducos, A. Meiotic segregation analysis in cows carrying the t(1;29) Robertsonian translocation. Cytogenet. Genome Res. 2008, 120, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Ducos, A.; Revay, T.; Kovacs, A.; Hidas, A.; Pinton, A.; Bonnet-Garnier, A.; Molteni, L.; Slota, E.; Switonski, M.; Arruga, M.V.; et al. Cytogenetic screening of livestock populations in Europe: An overview. Cytogenet. Genome Res. 2008, 120, 26–41. [Google Scholar] [CrossRef] [PubMed]

| Karyotype | Total Number of Cases |
|---|---|
| Heterozygous RobT (1:29) | 5 |
| Homozygous RobT (1:29) | 2 |
| RECT (1:25) | 1 |
| RECT (12:23) | 2 |
| Complex Translocation (26) | 1 |
| XX/XY chimeric * | 1 |
| Normal | 47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lewis, N.M.; Canedo-Ribeiro, C.; Rathje, C.C.; Jennings, R.L.; Danihel, M.; Bosman, L.M.; Silvestri, G.; Griffin, D.K. The Economic Burden of Chromosome Translocations and the Benefits of Enhanced Screening for Cattle Breeding. Animals 2022, 12, 1982. https://doi.org/10.3390/ani12151982
Lewis NM, Canedo-Ribeiro C, Rathje CC, Jennings RL, Danihel M, Bosman LM, Silvestri G, Griffin DK. The Economic Burden of Chromosome Translocations and the Benefits of Enhanced Screening for Cattle Breeding. Animals. 2022; 12(15):1982. https://doi.org/10.3390/ani12151982
Chicago/Turabian StyleLewis, Nicole M., Carla Canedo-Ribeiro, Claudia C. Rathje, Rebecca L. Jennings, Maxim Danihel, Lisa M. Bosman, Giuseppe Silvestri, and Darren K. Griffin. 2022. "The Economic Burden of Chromosome Translocations and the Benefits of Enhanced Screening for Cattle Breeding" Animals 12, no. 15: 1982. https://doi.org/10.3390/ani12151982
APA StyleLewis, N. M., Canedo-Ribeiro, C., Rathje, C. C., Jennings, R. L., Danihel, M., Bosman, L. M., Silvestri, G., & Griffin, D. K. (2022). The Economic Burden of Chromosome Translocations and the Benefits of Enhanced Screening for Cattle Breeding. Animals, 12(15), 1982. https://doi.org/10.3390/ani12151982