
Exploring the Potential of Myrothamnus flabellifolius Welw. (Resurrection Tree) as a Phytogenic Feed Additive in Animal Nutrition


Abstract
Simple Summary
Abstract
1. Introduction
2. Nutritional Composition of Myrothamnus flabellifolius (Welw.)
3. Phytochemical Composition of Myrothamnus flabellifolius (Welw.)
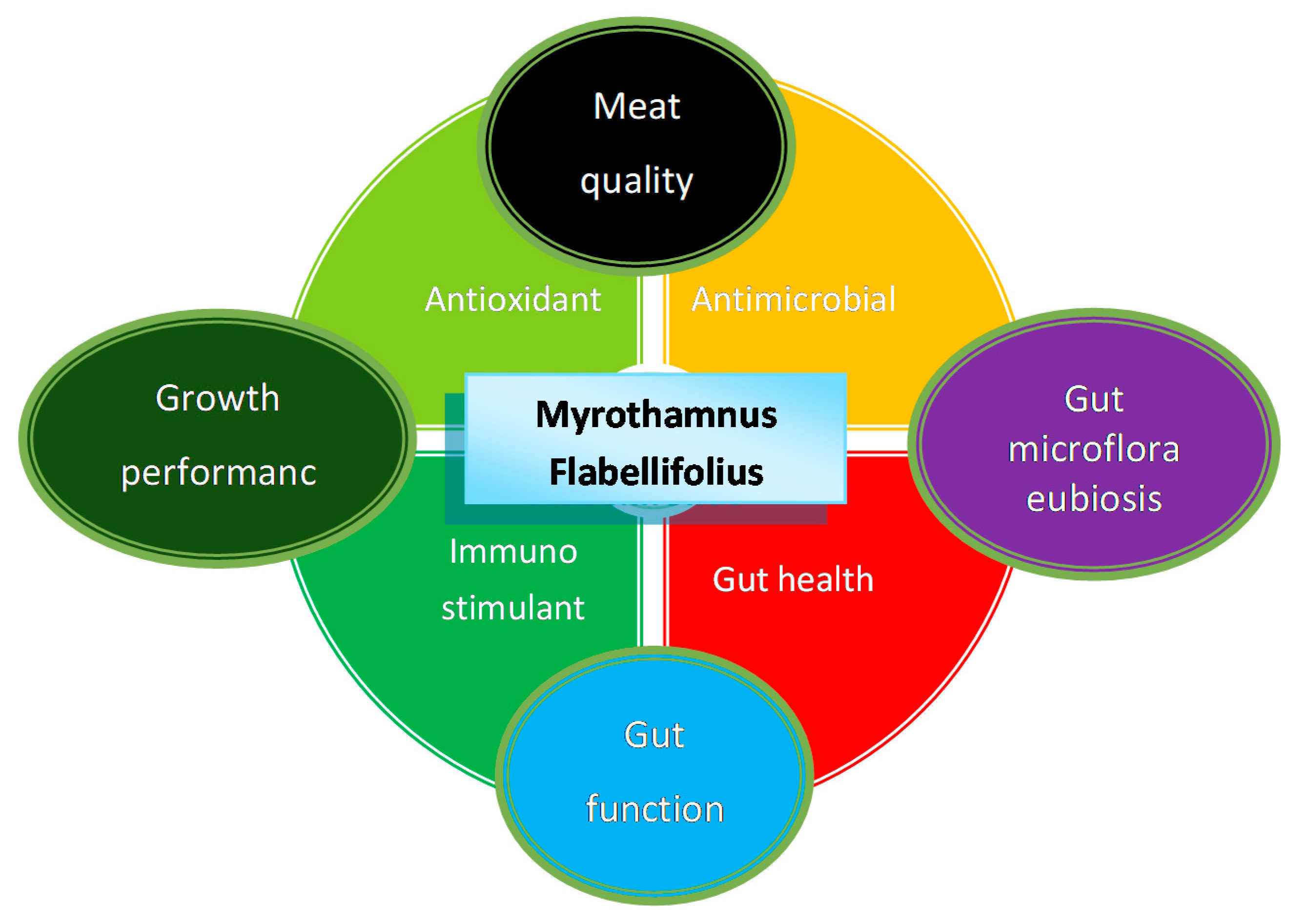
4. Potential Animal Production and Health Benefits
4.1. Antioxidant Effects
4.2. Antimicrobial Potential
4.3. Growth Performance and Health Effects
5. Limitations to the Use of M. flabellifolius (Welw.) in Animal Nutrition
5.1. Low Biomass and Seasonal Availability
5.2. Bioaccessibility, Bioavailability, and Transfer Efficiencies of Phytogenic Nutrients
5.3. Antinutritional Issues and Potential Adverse Effects and Toxicity
6. Improving Bioavailability and Bioaccessibility of Nutrients in Myrothamnus flabellifolius (Welw.)
6.1. Extraction of Essential Oil and Oleoresins
6.2. Microbial Fermentation
6.3. Microencapsulation
6.4. Blending of M. flabellifolius with Complimentary Compounds
7. Future Perspective and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moore, J.P.; Westall, K.L.; Ravenscroft, N.; Farrant, J.M.; Lindsey, G.G.; Brandt, W.F. The predominant polyphenol in the leaves of the resurrection plant Myrothamnus flabellifolius, 3,4,5 tri-O-galloylquinic acid, protects membranes against desiccation and free radical-induced oxidation. Biochem. J. 2004, 385, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.P.; Farrant, J.M.; Lindsey, G.G.; Brandt, W.F. The South African and Namibian Populations of the Resurrection Plant Myrothamnus flabellifolius are Genetically Distinct and Display Variation in Their Galloylquinic Acid Composition. J. Chem. Ecol. 2005, 31, 2823–2834. [Google Scholar] [CrossRef] [PubMed]
- Bentley, J.; Moore, J.P.; Farrant, J.M. Metabolomic profiling of the desiccation-tolerant medicinal shrub Myrothamnus flabellifolia indicates phenolic variability across its natural habitat: Implications for tea and cosmetics production. Molecules 2019, 24, 1240. [Google Scholar] [CrossRef] [PubMed]
- Bentley, J.; Moore, J.P.; Farrant, J.M. Metabolomics as a complement to phylogenetics for assessing intraspecific boundaries in the desiccation-tolerant medicinal shrub Myrothamnus flabellifolia (Myrothamnaceae). Phytochemistry 2019, 159, 127–136. Available online: https://www.sciencedirect.com/science/article/pii/S003194221830517X (accessed on 15 June 2021). [CrossRef] [PubMed]
- Koonjul, P.K.; Brandt, W.F.; Lindsey, G.G.; Farrant, J.M. Isolation and Characterisation of Chloroplasts from Myrothamnus flabellifolius Welw. J. Plant Physiol. 2000, 156, 584–594. Available online: https://www.sciencedirect.com/science/article/pii/S0176161700802175 (accessed on 15 June 2021). [CrossRef]
- Nicoletti, M.; Maggi, F.; Papa, F.; Vittori, S.; Quassinti, L.; Bramucci, M.; Lupidi, G.; Petrelli, D.; Vitali, L.A.; Ralaibia, E.; et al. In vitro biological activities of the essential oil from the ‘resurrection plant’ Myrothamnus moschatus (Baillon) Niedenzu endemic to Madagascar. Nat. Prod. Res. 2012, 26, 2291–2300. [Google Scholar] [CrossRef] [PubMed]
- Erhabor, J.O.; Komakech, R.; Kang, Y.; Tang, M.; Matsabisa, M.G. Ethnopharmacological importance and medical applications of Myrothamnus flabellifolius Welw. (Myrothamnaceae)—A review. J. Ethnopharmacol. 2020, 252, 112576. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, A.M.; Klepser, M.E.; Ernst, E.J.; Keele, D.; Roling, E.; Van Vuuren, S.; Demirci, B.; Baser, K.F.C.; van Wyk, B.-E.; Jäger, A.K. The composition and antimicrobial activity of the essential oil of the resurrection plant Myrothamnus flabellifolius. S. Afr. J. Bot. 2002, 68, 100–105. [Google Scholar] [CrossRef]
- Motlhanka, D.M.T.; Mathapa, G. Antioxidant activities of crude extracts from medicinal plants used by diabetic patients in Eastern Botswana. J. Med. Plants Res. 2012, 6, 5460–5463. [Google Scholar]
- Maroyi, A. Traditional use of medicinal plants in south-central Zimbabwe: Review and perspectives. J. Ethnobiol. Ethnomed. 2013, 9, 31. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, P.; Kwape, T.E.; Majinda, R.R.T. Antioxidant and antidiabetic potential of Myrothamnus flabellifolius found in Botswana. Cogent. Biol. 2016, 2, 1275403. [Google Scholar] [CrossRef]
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of plant polyphenols to combat oxidative stress and inflammatory processes in farm animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628. [Google Scholar] [CrossRef]
- Syed, B.; Kesselring, J.; Sánchez, J.; Gracia, M. Growth Performance and Nutrient Digestibility in Broiler Chickens Fed with an Encapsulated Blend of a Phytogenic Feed Additive. J. World’s Poult. Res. 2021, 11, 278–285. [Google Scholar] [CrossRef]
- Elbein, A.D.; Pan, Y.T.; Pastuszak, I.; Carroll, D. New insights on trehalose: A multifunctional molecule. Glycobiology 2003, 13, 17R–27R. [Google Scholar] [CrossRef] [PubMed]
- Brar, J.; Fultang, N.; Askey, K.; Tettamanzi, M.C.; Peethambaran, B. A novel anti-triple negative breast cancer compound isolated from medicinal herb Myrothamnus flabellifolius. J. Med. Plants Res. 2018, 12, 7–14. [Google Scholar]
- Chukwuma, C.I.; Matsabisa, M.G.; Rautenbach, F.; Rademan, S.; Oyedemi, S.O.; Chaudhary, S.K.; Javu, M. Evaluation of the nutritional composition of Myrothamnus flabellifolius (Welw.) herbal tea and its protective effect against oxidative hepatic cell injury. J. Food Biochem. 2019, 43, e13026. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, C.; Petereit, F.; Lechtenberg, M.; Liefländer-Wulf, U.; Hensel, A. Qualitative and quantitative phytochemical characterization of Myrothamnus flabellifolia Welw. Fitoterapia 2016, 114, 69–80. [Google Scholar] [CrossRef]
- Goddijn, O.; Smeekens, S. Sensing trehalose biosynthesis in plants. Plant J. 1998, 14, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Gechev, T.S.; Hille, J.; Woerdenbag, H.J.; Benina, M.; Mehterov, N.; Toneva, V.; Fernie, A.R.; Mueller-Roeber, B. Natural products from resurrection plants: Potential for medical applications. Biotechnol. Adv. 2014, 32, 1091–1101. [Google Scholar] [CrossRef] [PubMed]
- Singer, M.A.; Lindquist, S. Multiple effects of trehalose on protein folding in vitro and in vivo. Mol. Cell. 1998, 1, 639–648. [Google Scholar] [CrossRef]
- Feofilova, E.P.; Usov, A.I.; Mysyakina, I.S.; Kochkina, G.A. Trehalose: Chemical Structure, Biological Functions, and practical application. Microbiology 2014, 83, 184–194. [Google Scholar] [CrossRef]
- Colaco, C.; Roser, B. Trehalose-a multifunctional additive for food preservation. In Food Packaging and Preservation; Springer: Berlin/Heidelberg, Germany, 1994; pp. 123–140. [Google Scholar]
- Jun, H.; Hong-biao, D.; Guo-qi, Z. Effect of Trehalose on the Growth and Carcass Performance of AA Broilers. J. Agric. Sci. Technol. 2009, 11, 58. [Google Scholar]
- Ruangpanit, Y.; Matsushita, K.; Mukai, K.; Kikusato, M. Effect of trehalose supplementation on growth performance and intestinal morphology in broiler chickens. Vet. Anim. Sci. 2020, 10, 100142. [Google Scholar] [CrossRef] [PubMed]
- Drennan, P.M.; Smith, M.T.; Goldsworthy, D.; Van Staden, J. The occurrence of trehalose in the leaves of the desiccation-tolerant angiosperm Myrothamnus flabellifolius Welw. J. Plant Physiol. 1993, 142, 493–496. [Google Scholar] [CrossRef]
- Aoki, N.; Furukawa, S.; Sato, K.; Kurokawa, Y.; Kanda, S.; Takahashi, Y.; Mitsuzumi, H.; Itabashi, H. Supplementation of the diet of dairy cows with trehalose results in milk with low lipid peroxide and high antioxidant content. J. Dairy Sci. 2010, 93, 4189–4195. [Google Scholar] [CrossRef] [PubMed]
- Sato, K. Molecular nutrition: Interaction of nutrients, gene regulations and performances. Anim. Sci. J. 2016, 87, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Miura, H.; Mukai, K.; Sudo, K.; Haga, S.; Suzuki, Y.; Kobayashi, Y.; Koike, S. Effect of trehalose supplementation in milk replacer on the incidence of diarrhea and fecal microbiota in preweaned calves. J. Anim. Sci. 2021, 99, skab012. [Google Scholar] [CrossRef]
- Chen, Y.; Srionnual, S.; Onda, T.; Yanagida, F. Effects of prebiotic oligosaccharides and trehalose on growth and production of bacteriocins by lactic acid bacteria. Lett. Appl. Microbiol. 2007, 45, 190–193. [Google Scholar] [CrossRef]
- Bhuiyan, M.M.; Gao, F.; Chee, S.H.; Iji, P.A. Minimising weight loss in new broiler hatchlings through early feeding of simple sugars. Anim. Prod. Sci. 2011, 51, 1002–1007. [Google Scholar] [CrossRef]
- Iji, P.A.; Saki, A.A.; Tivey, D.R. Intestinal structure and function of broiler chickens on diets supplemented with a mannan oligosaccharide. J. Sci. Food Agric. 2001, 81, 1186–1192. [Google Scholar] [CrossRef]
- Kikusato, M.; Nanto, F.; Mukai, K.; Toyomizu, M. Effects of trehalose supplementation on the growth performance and intestinal innate immunity of juvenile chicks. Br. Poult. Sci. 2016, 57, 375–380. [Google Scholar] [CrossRef]
- Lee, H.-J.; Yoon, Y.-S.; Lee, S.-J. Mechanism of neuroprotection by trehalose: Controversy surrounding autophagy induction. Cell Death Dis. 2018, 9, 712. [Google Scholar] [CrossRef]
- Bianchi, G.; Gamba, A.; Limiroli, R.; Pozzi, N.; Elster, R.; Salamini, F.; Bartels, D. The unusual sugar composition in leaves of the resurrection plant Myrothamnus flabellifolia. Physiol. Plant. 1993, 87, 223–226. [Google Scholar] [CrossRef]
- Sawangwan, T. Glucosylglycerol on performance of prebiotic potential. Funct. Foods Health Dis. 2015, 5, 427–436. [Google Scholar] [CrossRef][Green Version]
- Bolivar, J.M.; Luley-Goedl, C.; Leitner, E.; Sawangwan, T.; Nidetzky, B. Production of glucosyl glycerol by immobilized sucrose phosphorylase: Options for enzyme fixation on a solid support and application in microscale flow format. J. Biotechnol. 2017, 257, 131–138. [Google Scholar] [CrossRef]
- Jeong, J.-W.; Seo, D.-H.; Jung, J.-H.; Park, J.-H.; Baek, N.-I.; Kim, M.-J.; Park, C.-S. Biosynthesis of glucosyl glycerol, a compatible solute, using intermolecular transglycosylation activity of amylosucrase from Methylobacillus flagellatus KT. Appl. Biochem. Biotechnol. 2014, 173, 904–917. [Google Scholar] [CrossRef]
- Wolf, S.; Becker, J.; Tsuge, Y.; Kawaguchi, H.; Kondo, A.; Marienhagen, J.; Bott, M.; Wendisch, V.F.; Wittmann, C. Advances in metabolic engineering of Corynebacterium glutamicum to produce high-value active ingredients for food, feed, human health, and well-being. Essays Biochem. 2021, 65, 197–212. [Google Scholar]
- Mikkat, S.; Galinski, E.A.; Berg, G.; Minkwitz, A.; Schoor, A. Salt adaptation in pseudomonads: Characterization of glucosylglycerol-synthesizing isolates from brackish coastal waters and the rhizosphere. Syst. Appl. Microbiol. 2000, 23, 31–40. [Google Scholar] [CrossRef]
- Kausar, S.; Humayun, A.; Ahmed, Z.; Abbas, M.A.; Tahir, A. Effect of apple cider vinegar on glycemic control, hyperlipidemia and control on body weight in type 2 diabetes patients. Int. J. Med. Res. Health Sci. 2019, 8, 59–74. [Google Scholar]
- Rastegari, H.; Jazini, H.; Ghaziaskar, H.S.; Yalpani, M. Applications of biodiesel by-products. In Biodiesel; Springer: Berlin/Heidelberg, Germany, 2019; pp. 101–125. [Google Scholar]
- Amer, S.A.; Naser, M.A.F.; Abdel-Wareth, A.A.A.; Saleh, A.A.; Elsayed, S.A.M.; Metwally, A.E. Effect of dietary supplementation of alpha-galactosidase on the growth performance, ileal digestibility, intestinal morphology, and biochemical parameters in broiler chickens. BMC Vet. Res. 2020, 16, 144. [Google Scholar] [CrossRef]
- Qiang, X.; YongLie, C.; QianBing, W. Health benefit application of functional oligosaccharides. Carbohydr. Polym. 2009, 77, 435–441. [Google Scholar] [CrossRef]
- Patel, S.; Goyal, A. Functional oligosaccharides: Production, properties and applications. World J. Microbiol. Biotechnol. 2011, 27, 1119–1128. [Google Scholar] [CrossRef]
- Bednarczyk, M.; Stadnicka, K.; Kozłowska, I.; Abiuso, C.; Tavaniello, S.; Dankowiakowska, A.; Sławińska, A.; Maiorano, G. Influence of different prebiotics and mode of their administration on broiler chicken performance. Animal 2016, 10, 1271–1279. [Google Scholar] [CrossRef]
- Bogucka, J.; Ribeiro, D.M.; Bogusławska-Tryk, M.; Dankowiakowska, A.; da Costa, R.P.R.; Bednarczyk, M. Microstructure of the small intestine in broiler chickens fed a diet with probiotic or synbiotic supplementation. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1785–1791. [Google Scholar] [CrossRef]
- Xu, G.; Xing, W.; Li, T.; Ma, Z.; Liu, C.; Jiang, N.; Luo, L. Effects of dietary raffinose on growth, non-specific immunity, intestinal morphology and microbiome of juvenile hybrid sturgeon (Acipenser baeri Brandt♀× A. schrenckii Brandt♂). Fish Shellfish Immunol. 2018, 72, 237–246. [Google Scholar] [CrossRef]
- Jiang, H.Q.; Gong, L.M.; Ma, Y.X.; He, Y.H.; Li, D.F.; Zhai, H.X. Effect of stachyose supplementation on growth performance, nutrient digestibility and caecal fermentation characteristics in broilers. Br. Poult. Sci. 2006, 47, 516–522. [Google Scholar] [CrossRef]
- Migas, P.; Krauze-Baranowska, M. The significance of arbutin and its derivatives in therapy and cosmetics. Phytochem. Lett. 2015, 13, 35–40. [Google Scholar] [CrossRef]
- Masumoto, S.; Nakamura, Y.; Haginaka, J. Molecularly imprinted polymers for arbutin and rutin by modified precipitation polymerization and their application for selective extraction of rutin in nutritional supplements. J. Pharm. Biomed. Anal. 2021, 205, 114294. [Google Scholar] [CrossRef]
- Bonifacio, M.A.; Cerqueni, G.; Cometa, S.; Licini, C.; Sabbatini, L.; Mattioli-Belmonte, M.; De Giglio, E. Insights into arbutin effects on bone cells: Towards the development of antioxidant titanium implants. Antioxidants 2020, 9, 579. [Google Scholar] [CrossRef]
- Murray, A.H.; Iason, G.R.; Stewart, C. Effect of simple phenolic compounds of heather (Calluna vulgaris) on rumen microbial activity in vitro. J. Chem. Ecol. 1996, 22, 1493–1504. [Google Scholar] [CrossRef]
- Noikotra, K.; Chaveeracha, A.; Sudmoona, R.; Taneea, T.; Patarapadungkita, N. Phytochemicals, cytotoxicity, and genotoxicity of three Artocarpus species reveal arbutin in A. lacucha. Sci. Asia 2018, 44, 170–178. [Google Scholar] [CrossRef]
- Cheikhyoussef, A.; Summers, R.W.; Kahaka, G. Qualitative and quantitative analysis of phytochemical compounds in Namibian Myrothamnus flabellifolius Phytochemical Compounds in M. flabellifolius. Int. Sci. Technol. J. Namibia 2015, 5, 71–83. [Google Scholar]
- Abdelli, N.; Sol, D.; Francisco, P. Phytogenic Feed Additives in Poultry: Achievements, Prospective and Challenges. Animals 2021, 11, 3471. [Google Scholar] [CrossRef]
- Ajao, A.A.; Ashafa, A.O.T. Kinetics of inhibitory action of Myrothamnus flabellifolius Welw. on the activities of α-amylase and α-glucosidase. Comp. Clin. Pathol. 2017, 26, 911–920. [Google Scholar] [CrossRef]
- Kilic, I.; Yeşiloğlu, Y.; Bayrak, Y. Spectroscopic studies on the antioxidant activity of ellagic acid. Spectrochim. Acta Part A Mol. Spectrosc. 2014, 130, 447–452. [Google Scholar] [CrossRef]
- Cermak, R.; Landgraf, S.; Wolffram, S. The bioavailability of quercetin in pigs depends on the glycoside moiety and on dietary factors. J. Nutr. 2003, 133, 2802–2807. [Google Scholar] [CrossRef]
- Surai, P.F. Polyphenol compounds in the chicken/animal diet: From the past to the future. J. Anim. Physiol. Anim. Nutr. 2014, 98, 19–31. [Google Scholar] [CrossRef]
- Kamng’ona, A.; Moore, J.P.; Lindsey, G.; Brandt, W. Inhibition of HIV-1 and M-MLV reverse transcriptases by a major polyphenol (3,4,5 tri-O-galloylquinic acid) present in the leaves of the South African resurrection plant, Myrothamnus flabellifolia. J. Enzym. Inhib. Med. Chem. 2011, 26, 843–853. [Google Scholar] [CrossRef]
- Li, Y.; Ding, Y.; Xiao, W.; Zhu, J.-B. Investigation on the active ingredient and mechanism of Cannabis sativa L. for treating epilepsy based on network pharmacology. Biotechnol. Biotechnol. Equip. 2021, 35, 994–1009. [Google Scholar] [CrossRef]
- Motta, E.V.S.; Lemos, M.; Costa, J.C.; Banderó-Filho, V.C.; Sasse, A.; Sheridan, H.; Bastos, J.K. Galloylquinic acid derivatives from Copaifera langsdorffii leaves display gastroprotective activity. Chem. Biol. Interact. 2017, 261, 145–155. [Google Scholar] [CrossRef]
- Lemos, M.; Santin, J.R.; Mizuno, C.S.; Boeing, T.; de Sousa, J.P.B.; Nanayakkara, D.; Bastos, J.K.; de Andrade, S.F. Copaifera langsdorffii: Evaluation of potential gastroprotective of extract and isolated compounds obtained from leaves. Rev. Bras. Farmacogn. 2015, 25, 238–245. [Google Scholar] [CrossRef]
- Bardy, G.; Virsolvy, A.; Quignard, J.F.; Ravier, M.A.; Bertrand, G.; Dalle, S.; Cros, G.; Magous, R.; Richard, S.; Oiry, C. Quercetin induces insulin secretion by direct activation of L-type calcium channels in pancreatic beta cells. Br. J. Pharmacol. 2013, 169, 1102–1113. [Google Scholar] [CrossRef]
- Salehi, B.; Machin, L.; Monzote, L.; Sharifi-Rad, J.; Ezzat, S.M.; Salem, M.A.; Merghany, R.M.; el Mahdy, N.M.; Kılıç, C.S.; Sytar, O.; et al. Therapeutic potential of quercetin: New insights and perspectives for human health. ACS Omega 2020, 5, 11849–11872. [Google Scholar] [CrossRef]
- Chagas, V.T.; França, L.M.; Malik, S.; Paes, A.M.D.A. Syzygium cumini (L.) skeels: A prominent source of bioactive molecules against cardiometabolic diseases. Front. Pharmacol. 2015, 6, 259. [Google Scholar] [CrossRef]
- Ahmadipour, B.; Hassanpour, H.; Khajali, F. Evaluation of hepatic lipogenesis and antioxidant status of broiler chickens fed mountain celery. BMC Vet. Res. 2018, 14, 234. [Google Scholar] [CrossRef]
- Ying, L.; Chaudhry, M.T.; Xiao, F.; Mao, Y.; Wang, M.; Wang, B.; Wang, S.; Li, Y. The effects and mechanism of quercetin dietary supplementation in streptozotocin-induced hyperglycemic arbor acre broilers. Oxidative Med. Cell. Longev. 2020, 10, 9585047. [Google Scholar] [CrossRef]
- Sierzant, K.; Orda, J.; Korzeniowska, M.; Malicki, A. Effect of dietary supplementation with extracts of rosemary, olive leaves, pine bark and quercetin on selected performance indices of broiler chickens and microbiological status of their ileum. Med. Weter 2019, 75, 247–252. [Google Scholar] [CrossRef]
- Rasoanaivo, P.; Ralaibia, E.; Maggi, F.; Papa, F.; Vittori, S.; Nicoletti, M. Phytochemical investigation of the essential oil from the ‘resurrection plant’ Myrothamnus moschatus (Baillon) Niedenzu endemic to Madagascar. J. Essent. Oil Res. 2012, 24, 299–304. [Google Scholar] [CrossRef]
- Zorzetto, C.; Sánchez-Mateo, C.C.; Rabanal, R.M.; Lupidi, G.; Bramucci, M.; Quassinti, L.; Iannarelli, R.; Papa, F.; Maggi, F. Antioxidant activity and cytotoxicity on tumour cells of the essential oil from Cedronella canariensis var. canariensis (L.) Webb & Berthel.(Lamiaceae). Nat. Prod. Res. 2015, 29, 1641–1649. [Google Scholar]
- Fernández-Cervantes, M.; Pérez-Alonso, M.J.; Blanco-Salas, J.; Soria, A.C.; Ruiz-Téllez, T. Analysis of the essential oils of Chamaemelum fuscatum (Brot.) Vasc. from Spain as a contribution to reinforce its ethnobotanical use. Forests 2019, 10, 539. [Google Scholar] [CrossRef]
- Fotina, A.A.; Fisinin, V.I.; Surai, P.F. Recent developments in usage of natural antioxidants to improve chicken meat production and quality. Bulg. J. Agric. Sci. 2013, 19, 889–896. [Google Scholar]
- Oladeji, I.S.; Adegbenro, M.; Osho, I.B.; Olarotimi, O.J. The Efficacy of Phytogenic Feed Additives in Poultry Production: A Review. Turk. J. Agric.-Food Sci. Technol. 2019, 7, 2038–2041. [Google Scholar] [CrossRef]
- Bhebhe, M.; Chipurura, B.; Muchuweti, M. Determination and comparison of phenolic compound content and antioxidant activity of selected local Zimbabwean herbal teas with exotic Aspalathus linearis. S. Afr. J. Bot. 2015, 100, 213–218. [Google Scholar] [CrossRef]
- Van Vuuren, S.F.; Viljoen, A.M. A comparative investigation of the antimicrobial properties of indigenous South African aromatic plants with popular commercially available essential oils. J. Essent. Oil Res. 2006, 18 (Suppl. 1), 66–71. [Google Scholar] [CrossRef]
- Van Vuuren, S.F.; Du Toit, L.C.; Parry, A.; Pillay, V.; Choonara, Y.E. Encapsulation of Essential Oils within a Polymeric Liposomal Formulation for Enhancement of Antimicrobial Efficacy. Nat. Prod. Commun. 2010, 5, 1401–1408. [Google Scholar] [CrossRef] [PubMed]
- Favaretto, J.A.; Alba, D.F.; Marchiori, M.S.; Marcon, H.J.; Souza, C.F.; Baldissera, M.D.; Bianchi, A.E.; Zanluchi, M.; Klein, B.; Wagner, R.; et al. Supplementation with a blend based on micro-encapsulated carvacrol, thymol, and cinnamaldehyde in lambs feed inhibits immune cells and improves growth performance. Livest. Sci. 2020, 240, 104144. [Google Scholar] [CrossRef]
- Yitbarek, M.B. Phytogenics as feed additives in poultry production: A review. Int. J. Extensive Res. 2015, 3, 49–60. [Google Scholar]
- Alloui, M.N.; Agabou, A.; Alloui, N. Application of herbs and phytogenic feed additives in poultry production—A Review. Glob. J. Anim. Sci. Res. 2014, 2, 234–243. [Google Scholar]
- Yang, C.; Chowdhury, M.A.K.; Hou, Y.; Gong, J. Phytogenic Compounds as Alternatives to In-Feed Antibiotics: Potentials and Challenges in Application. Pathogens 2015, 4, 137–156. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.P.; Waldron, M.; Lindsey, G.G.; Farrant, J.M.; Brandt, W.F. An ultrastructural investigation of the surface microbiota present on the leaves and reproductive structures of the resurrection plant Myrothamnus flabellifolia. S. Afr. J. Bot. 2011, 77, 485–491. [Google Scholar] [CrossRef]
- Iriyanti, N.; Hartoyo, B. Encapsulated Fermeherbafit Bioavailability and the Application to Broilers. J. Agric. Sci. Technol. A 2019, 9, 157–165. [Google Scholar]
- Bacova, K.; Eglseer, K.Z.; Räuber, G.K.; Chrastinova, L.; Laukova, A.; Takacsova, M.; Simonova, M.P.; Placha, L. Effect of Sustained Administration of Thymol on Its Bioaccessibility and Bioavailability in Rabbits. Animals 2021, 11, 2595. [Google Scholar] [CrossRef] [PubMed]
- Alkhalidy, H.; Wang, Y.; Liu, D. Dietary flavonoids in the prevention of T2D: An overview. Nutrients 2018, 10, 438. [Google Scholar] [CrossRef] [PubMed]
- Angelino, D.; Cossu, M.; Marti, A.; Zanoletti, M.; Chiavaroli, L.; Brighenti, F.; Rio, D.D.; Martini, D. Bioaccessibility and bioavailability of phenolic compounds in bread: A review. Food Funct. 2017, 8, 2368–2393. [Google Scholar] [CrossRef] [PubMed]
- Kramer, N.I.; Krismartina, M.; Blaauboer, B.J.; Hermens, J.L.M. Measuring and modeling bioavailability in in vitro systems. Chem. Biol. Interact. 2007, 2, 137. [Google Scholar] [CrossRef]
- Weichselbaum, E.; Buttriss, J.L. Polyphenols in the Diet; Wiley Online Library: Hoboken, NJ, USA, 2010. [Google Scholar]
- Scalbert, A.; Williamson, G. Chocolate: Modern science investigates an ancient medicine. J. Med. Food 2000, 3, 121–125. [Google Scholar]
- Rivaroli, D.C.; Guerrero, A.; Valero, M.V.; Zawadzki, F.; Eiras, C.E.; del Mar Campo, M.; Sañudo, C.; Jorge, A.M.; do Prado, I.N. Effect of essential oils on meat and fat qualities of crossbred young bulls finished in feedlots. Meat Sci. 2016, 121, 278–284. [Google Scholar] [CrossRef] [PubMed]
- da Costa, O.B.; Del Menezzi, C.H.S.; Benedito, L.E.C.; Resck, I.S.; Vieira, R.F.; Ribeiro Bizzo, H. Essential oil constituents and yields from leaves of Blepharocalyx salicifolius (Kunt) O. Berg and Myracrodruon urundeuva (Allemão) collected during daytime. Int. J. For. Res. 2014, 2014, 982576. [Google Scholar]
- Liu, M.; Luo, F.; Qing, Z.; Yang, H.; Liu, X.; Yang, Z.; Zeng, J. Chemical composition and bioactivity of essential oil of ten Labiatae species. Molecules 2020, 25, 4862. [Google Scholar] [CrossRef] [PubMed]
- Mandeya, K. Extraction of Essential oils from Myrothamnus Flabellifolious and Their Application as Beef Oxidative Stabilizers. Ph.D. Thesis, Bindura University of Science Education, Bindura, Zimbabwe, 2018. [Google Scholar]
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef]
- Sugiharto, S.; Widiastuti, E.; Pratama, A.R.; Wahyuni, H.I.; Yudiarti, T.; Sartono, T.A. Hematological and Intestinal Responses of Broilers to Dietary Supplementations of Lactic Fermented Turmeric, Black Pepper or a Mixture of Both. Acta Univ. Agric. Silvic. Mendel. Brun. 2021, 69, 10. [Google Scholar] [CrossRef]
- Qiao, H.; Song, Y.; Shi, H.; Bian, C. Fermented Astragalus in diet altered the composition of fecal microbiota in broiler chickens. AMB Express 2018, 8, 151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Sun, Z.Y.; Cao, F.L.; Ahmad, H.; Yang, X.H.; Zhao, L.G.; Wang, T. Effects of dietary supplementation with fermented ginkgo leaves on antioxidant capacity, intestinal morphology and microbial ecology in broiler chicks. Br. Poult. Sci. 2015, 56, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Cao, F.; Sun, Z.; Yu, W.; Zhao, L.; Wang, G.; Wang, T. Effect of feeding Aspergillus niger-fermented Ginkgo biloba-leaves on growth, small intestinal structure and function of broiler chicks. Livest. Sci. 2012, 147, 170–180. [Google Scholar] [CrossRef]
- Hlahla, L.N.; Mudau, F.N.; Mariga, I.K. Effect of fermentation temperature and time on the chemical composition of bush tea (Athrixia phylicoides DC.). J. Med. Plants Res. 2010, 4, 824–829. [Google Scholar]
- Gong, J.; Yin, F.; Hou, Y.; Yin, Y. Review: Chinese herbs as alternatives to antibiotics in feed for swine and poultry production: Potential and challenges in application. Can. J. Anim. Sci. 2014, 94, 223–241. [Google Scholar] [CrossRef]
- Mandey, J.S.; Sompie, F.N. Phytogenic Feed Additives as an Alternative to Antibiotic Growth Promoters in Poultry Nutrition. Adv. Stud. 21st Century Anim. Nutr. 2021, 8, 19. [Google Scholar]
- Nouri, A. Chitosan nano-encapsulation improves the effects of mint, thyme, and cinnamon essential oils in broiler chickens. Br. Poult. Sci. 2019, 60, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Kailasapathy, K. Encapsulation technologies for functional foods and nutraceutical product development. CAB Rev. 2009, 4, 1–19. [Google Scholar] [CrossRef]
- Gökmen, S.; Üniversitesi, K.M.; Palamutoglu, R. Applications of Encapsulation in Food Industry Determination of the Effects of Various Levels of Lemon and Pomegranate Peels on the Microbiological Properties of Cig Köfte View Project Author View Project. Available online: www.teknolojikarastirmalar.com (accessed on 7 February 2022).
- Öztürk, E.; Temiz, U. Encapsulation Methods and Use in Animal Nutrition. Selcuk J. Agric. Food Sci. 2018, 32, 624–631. [Google Scholar]
- Huesca-Urióstegui, K.; García-Valderrama, E.J.; Gutierrez-Uribe, J.A.; Antunes-Ricardo, M.; Guajardo-Flores, D. Nanofiber Systems as Herbal Bioactive Compounds Carriers: Current Applications in Healthcare. Pharmaceutics 2022, 14, 191. [Google Scholar] [CrossRef] [PubMed]
- Gopi, S.; Amalraj, A.; Varma, K.; Jude, S.; Reddy, P.B.; Divya, C.; Haponiuk, J.T.; Thomas, S. Turmeric nanofiber-encapsulated natural product formulation act as a phytogenic feed additive—A study in broilers on growth performance, biochemical indices of blood, and E. coli in cecum. Int. J. Polym. Mater. Polym. Biomater. 2018, 67, 581–588. [Google Scholar] [CrossRef]
- Tolve, R.; Tchuenbou-magaia, F.; Di, M.; Carmela, M.; Scarpa, T.; Galgano, F. Encapsulation of bioactive compounds for the formulation of functional animal feeds: The biofortification of derivate foods. Anim. Feed Sci. Technol. 2021, 279, 115036. [Google Scholar] [CrossRef]
- Hafeez, A.; Männer, K.; Schieder, C.; Zentek, J. Effect of supplementation of phytogenic feed additives (powdered vs. encapsulated) on performance and nutrient digestibility in broiler chickens. Poult. Sci. 2016, 95, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.; Almeida, D.; Bouça, G.; Silva, M.; Maria, D.; Freire, G.; Moura-Nunesd, N.; Monteiro, M.; Perrone, D. Bioaccessibility and gut metabolism of phenolic compounds of breads added with green co ff ee infusion and enzymatically bioprocessed. Food Chem. 2020, 333, 127473. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Le, T.V.A.; Dang, N.N.; Nguyen, D.C.; Nguyen, P.T.N.; Tran, T.T.; Nguyen, Q.V.; Bach, L.G.; Nguyen Pham, D.T. Microencapsulation of Essential Oils by Spray-Drying and Influencing Factors. J. Food Qual. 2021, 2021, 5525879. [Google Scholar] [CrossRef]
- Bag, A.; Chattopadhyay, R.R. Evaluation of synergistic antibacterial and antioxidant efficacy of essential oils of spices and herbs in combination. PLoS ONE 2015, 10, e0131321. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.H.; Seok, W.J.; Kim, I.H. Organic acids mixture as a dietary additive for pigs—A review. Animals 2020, 10, 952. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.B.; Luciano, F.B.; Miyada, V.S.; Gois, F.D. Herbal extracts and organic acids as natural feed additives in pig diets. S. Afr. J. Anim. Sci. 2013, 43, 181–193. [Google Scholar]
- Sawicki, R.; Golus, J.; Przekora, A.; Ludwiczuk, A.; Sieniawska, E.; Ginalska, G. Antimycobacterial activity of cinnamaldehyde in a Mycobacterium tuberculosis (H37Ra) model. Molecules 2018, 23, 2381. [Google Scholar] [CrossRef] [PubMed]
- Bhavaniramya, S.; Vishnupriya, S.; Al-Aboody, M.S.; Vijayakumar, R.; Baskaran, D. Role of essential oils in food safety: Antimicrobial and antioxidant applications. Grain Oil Sci. Technol. 2019, 2, 49–55. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, X.; Xin, H.; Chen, S.; Yang, C.; Duan, Y.; Yang, X. Effects of a protected inclusion of organic acids and essential oils as antibiotic growth promoter alternative on growth performance, intestinal morphology and gut microflora in broilers. Anim. Sci. J. 2017, 88, 1414–1424. [Google Scholar] [CrossRef] [PubMed]
- El-Azrak, K.E.-D.M.; Morsy, A.S.; Soltan, Y.A.; Hashem, N.M.; Sallam, S.M.A. Impact of specific essential oils blend on milk production, serum biochemical parameters and kid performance of goats. Anim. Biotechnol. 2021, 1–9. [Google Scholar] [CrossRef]
- Fascina, V.B.; Pasquali, G.A.M.; Carvalho, F.B.; Muro, E.M.; Vercese, F.; Aoyagi, M.M.; Pezzato, A.C.; Gonzales, E.; Sartori, J.R. Effects of phytogenic additives and organic acids, alone or in combination, on the performance, intestinal quality and immune responses of broiler chickens. Braz. J. Poult. Sci. 2017, 19, 497–508. [Google Scholar] [CrossRef]
- Dey, A.; Misra, S.S.; Dahiya, S.S.; Balhara, A.K.; Kumar, K.; Das, A.K.; Singh, K.; Attri, K.; Imaz, J.A.; Sarkar, N.C. Essential oils as phytogenic feed additive: Potential benefits on environment, livestock health and production. Int. J. Econ. Plants 2017, 4, 177–181. [Google Scholar]
- Hassan, H.M.A.; Youssef, A.W.; Ali, H.M.; Mohamed, M.A. Adding phytogenic material and/or organic acids to broiler diets: Effect on performance, nutrient digestibility and net profit. Asian J. Poult. Sci. 2015, 9, 97–105. [Google Scholar] [CrossRef]


{kind=link}
| Quantity | Potential Mode of Action | Reference | |
|---|---|---|---|
| Nutrient | |||
| Trehalose | 34.7 ± 7.2 mgg−1 | proteins and lipid stability | [25] |
| Raffinose | 2.49 g/100 g | osmotic stress protection | [16] |
| Gallic acid | 26.62% | Anti-obesity | [16] |
| Ferulic acid | 15.23% | Antiinflammation | [16] |
| Stachyose | 2.18 g/100 g | osmotic stress protection | [16] |
| Sucrose | 56.5 ± 6.6 mgg−1 | [25] | |
| Phytochemical | |||
| Carvacrol | - | Antibacterial, antioxidant | [17,57] |
| 3,4,5 tri-O-galloylquicic acid | 0.73, 0.32, and 0.0029 g/100 g | Membrane protectant, Anti-viral reverse transcriptase activity, | [3,4] |
| Galloylquinic acid | - | Antioxidant | [58] |
| Arbutin | - | Melanogenesis inhibitor | [23] |
| Ellagic acid | - | Antioxidant | [59] |
| Gallocatechin | 1.43 ± 0.03 mg/g | Chemoprotective, antioxidant, antibacterial | [4,56] |
| Trans-pinocarveol | 19.57% | Antimicrobial | [8] |
| Quercetin glucoside | 3 mg AWEw | Antioxidant | [60] |
| Pinocarvone | 11.13% | Antimicrobial | [8] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nantapo, C.W.T.; Marume, U. Exploring the Potential of Myrothamnus flabellifolius Welw. (Resurrection Tree) as a Phytogenic Feed Additive in Animal Nutrition. Animals 2022, 12, 1973. https://doi.org/10.3390/ani12151973
Nantapo CWT, Marume U. Exploring the Potential of Myrothamnus flabellifolius Welw. (Resurrection Tree) as a Phytogenic Feed Additive in Animal Nutrition. Animals. 2022; 12(15):1973. https://doi.org/10.3390/ani12151973
Chicago/Turabian StyleNantapo, Carlos Wyson Tawanda, and Upenyu Marume. 2022. "Exploring the Potential of Myrothamnus flabellifolius Welw. (Resurrection Tree) as a Phytogenic Feed Additive in Animal Nutrition" Animals 12, no. 15: 1973. https://doi.org/10.3390/ani12151973
APA StyleNantapo, C. W. T., & Marume, U. (2022). Exploring the Potential of Myrothamnus flabellifolius Welw. (Resurrection Tree) as a Phytogenic Feed Additive in Animal Nutrition. Animals, 12(15), 1973. https://doi.org/10.3390/ani12151973