
Sexual Dimorphism in the Chinese Endemic Species Hynobius maoershanensis (Urodela: Hynobiidae)

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Data Collection
2.3. Statistical Analysis
3. Results
3.1. The Relationship between Morphological Eigenvalues and SVL
3.2. Comparison of the Morphometric Variables of Males and Females
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shine, R. Ecological causes for the evolution of sexual dimorphism: A review of the evidence. Q. Rev. Biol. 1989, 64, 419–461. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, C.M.; Mann, E.H.J.; Stock, M.K.; Villamil, C.I.; Montague, M.J.; Dickinson, E.; Surratt, S.B.; Martinez, M.; Williams, S.A.; Anton, S.C.; et al. The ontogeny of sexual dimorphism in free-ranging rhesus macaques. Am. J. Biol. Anthropol. 2022, 177, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.; Gigena, D.J.; Benitez-Vieyra, S.M.; Valdez, D.J. Subtle sexual plumage color dimorphism and size dimorphism in a South American colonial breeder, the Monk Parakeet (Myiopsitta monachus). Avian Res. 2020, 11, 157–165. [Google Scholar] [CrossRef]
- Gazda, M.A.; Araujo, P.M.; Lopes, R.J.; Toomey, M.B.; Andrade, P.; Afonso, S.; Marques, C.; Nunes, L.; Pereira, P.; Trigo, S.; et al. A genetic mechanism for sexual dichromatism in birds. Science 2020, 368, 1270–1274. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Zhang, B.; Liu, Q.; Pan, T.; Gou, J. Sexual dimorphism in the Chinese endemic species Pachyhynobius shangchengensis Fei, Qu and Wu, 1983 (Urodela: Hynobiidae). PeerJ 2019, 7, e6408. [Google Scholar] [CrossRef]
- McClelland, B.E.; Ryan, M.J.; Wilczynski, W. Does sexual dimorphism vary by population? Laryngeal and ear anatomy in cricket frogs. Curr. Zool. 2019, 65, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Portik, D.M.; Blackburn, D.C.; McGuire, J.A. Macroevolutionary Patterns of Sexual Size Dimorphism Among African Tree Frogs (Family: Hyperoliidae). J. Hered. 2020, 111, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Hudson, C.M.; Fu, J. Male-biased sexual size dimorphism, resource defense polygyny, and multiple paternity in the Emei moustache toad (Leptobrachium boringii). PLoS ONE 2013, 8, e67502. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Seglie, D.; Roy, D.; Giacoma, C. Sexual dimorphism and age structure in a population of Tylototriton verrucosus (Amphibia: Salamandridae) from the Himalayan Region. Copeia 2010, 2010, 600–608. [Google Scholar] [CrossRef]
- Labus, N.; Cvijanović, M.; Vukov, T. Sexual size and shape dimorphism in Salamandra salamandra (Amphibia, Caudata, Salamandridae) from the central Balkans. Arch. Biol. Sci. 2013, 65, 969–975. [Google Scholar] [CrossRef]
- Reinhard, S.; Renner, S.; Kupfer, A. Sexual dimorphism and age of Mediterranean salamanders. Zoology 2015, 118, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Shine, R. Sexual selection and sexual dimorphism in the Amphibia. Copeia 1979, 2, 297–306. [Google Scholar] [CrossRef]
- Peter, P.; Alexander, K. Flesh and bone: An integrative approach towards sexual size dimorphism of a terrestrial salamander (genus Salamandrina). J. Morphol. 2018, 279, 1468–1479. [Google Scholar]
- Carvajal-Castro, J.D.; Vargas-Salinas, F.; Casas-Cardona, S.; Rojas, B.; Santos, J.C. Aposematism facilitates the diversification of parental care strategies in poison frogs. Sci. Rep. 2021, 11, 19047. [Google Scholar] [CrossRef] [PubMed]
- Shine, R.; Goiran, C. Sexual dimorphism in size and shape of the head in the sea snake Emydocephalus annulatus (Hydrophiinae, Elapidae). Sci. Rep. 2021, 11, 20026. [Google Scholar] [CrossRef] [PubMed]
- Blanckenhorn, W.U. Behavioral causes and consequences of sexual size dimorphism. Ethology 2005, 111, 977–1016. [Google Scholar] [CrossRef]
- Dursun, C.; Gul, S.; Ozdemir, N. Sexual size and shape dimorphism in Turkish common toads (Bufo bufo Linnaeus 1758). Anat. Rec. 2022, 305, 1548–1558. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Fu, J. Does life history shape sexual size dimorphism in anurans? A comparative analysis. BMC Evol. Biol. 2013, 13, 27. [Google Scholar] [CrossRef]
- Ringler, E.; Pašukonis, A.; Hödl, W.; Ringler, M. Tadpole transport logistics in a Neotropical poison frog: Indications for strategic planning and adaptive plasticity in anuran parental care. Front. Zool. 2013, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Requena, G.S.; Machado, G. Effects of egg attendance on male mating success in a harvestman with exclusive paternal care. Behav. Ecol. 2015, 26, 926–935. [Google Scholar] [CrossRef]
- Ah-King, M.; Kvarnemo, C.; Tullberg, B.S. The influence of territoriality and mating system on the evolution of male care: A phylogenetic study on fish. J. Evol. Biol. 2005, 18, 371–382. [Google Scholar] [CrossRef]
- Mank, J.E.; Promislow, D.E.; Avise, J.C. Phylogenetic perspectives in the evolution of parental care in ray-finned fishes. Evolution 2005, 59, 1570–1578. [Google Scholar] [CrossRef] [PubMed]
- Laporte, M.; Berrebi, P.; Claude, J.; Vinyoles, D.; Pou-Rovira, Q.; Raymond, J.; Magnan, P. The ecology of sexual dimorphism in size and shape of the freshwater blenny Salaria fluviatilis. Curr. Zool. 2018, 64, 183–191. [Google Scholar] [CrossRef]
- Székely, T.; Reynolds, J.D.; Figuerola, J. Sexual size dimorphism in shorebirds, gulls, and alcids: The influence of sexual and natural selection. Evolution 2000, 54, 1404–1413. [Google Scholar] [CrossRef]
- Li, X.; Yang, Z.; Yang, X.; Li, Q. Sexual Age and Size Dimorphism of Nidirana pleuraden. Sichuan J. Zool. 2022, 41, 284–289. [Google Scholar]
- Cruz-Elizalde, R.; Ramirez-Bautista, A.; Pacheco, L.F.R.; Lozano, A.; Rodriguez-Romero, F.D. Sexual dimorphism in size and shape among populations of the lizard Sceloporus variabilis (Squamata: Phrynosomatidae). Zoology 2020, 140, 125781. [Google Scholar] [CrossRef] [PubMed]
- Davies, N.; Halliday, T. Optimal mate selection in the toad Bufo bufo. Nature 1977, 269, 56–58. [Google Scholar] [CrossRef]
- Liao, H.; Xu, F.; Yang, W. Sexual size dimorphism of amphibians and reptiles: A review. Chin. J. Ecol. 2013, 32, 3082–3093. [Google Scholar]
- Shine, R. The evolution of large body size in females: A critique of Darwin’s “fecundity advantage” model. Am. Nat. 1988, 131, 124–131. [Google Scholar] [CrossRef]
- Yao, C.; Wang, Z.; Wang, F.; Jiang, G.; Lv, T.; Tang, Y.; Chen, M. Sexual Dimorphism and Female Reproductive Ability of Microhyla fissipes. Chin. J. Zool. 2020, 55, 317–322. [Google Scholar]
- Yao, C.; He, R.; Huang, Y.; Lv, T.; Wang, Z.; Liang, L.; Xiao, J.; Chen, M. Sexual Dimorphism of Tylototriton shanjing Yulong Snow Mountain Population. Sichuan J. Zool. 2019, 38, 666–671. [Google Scholar]
- Shetty, S.; Shine, R. Sexual divergence in diets and morphology in Fijian sea snakes Laticauda colubrina (Laticaudinae). Austral Ecol. 2002, 27, 77–84. [Google Scholar] [CrossRef]
- Cox, R.M.; Skelly, S.L.; John-Alder, H.B. A comparative test of adaptive hypotheses for sexual size dimorphism in lizards. Evolution 2003, 57, 1653–1669. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.; Lin, Y.; Lin, Y. Sexual size and shape dimorphism in an agamid lizard, Japalura swinhonis (Squamata: Lacertilia: Agamidae). Zool. Stud. 2009, 48, 351–361. [Google Scholar]
- Chen, W. Hynobius maoershanensis. Guangxi For. 2017, 10, 19. [Google Scholar]
- Zhou, F.; Jiang, A.; Jiang, D. A New Species of the Genus Hynobius from Guangxi Zhuang Autonomous Region China (Caudata Hynobiidae). Acta Zootaxonomica Sin. 2006, 31, 670–674. [Google Scholar]
- Available online: http://www.yyj.moa.gov.cn/gzdt/202102/t20210205_6361296.htm (accessed on 13 May 2022).
- Huang, H.; Bu, R.; Xie, H.; Hou, S.; Wu, Z. Habitat selection by Hynobius maoershanensis during its breeding period. Acta Ecol. Sin. 2019, 39, 6443–6451. [Google Scholar]
- Fei, L.; Ye, C.; Jiang, J.; Xie, F. An Illustrated Key to Chinese Amphibians; Sichuan Publishing House of Science and Technology: Chengdu, China, 2004; pp. 4–5. [Google Scholar]
- Gibbon, J.W.; Lovich, J.E. On the slider turtle (Trachemys scripta). Herpetol. Monogr. 1990, 4, 1–29. [Google Scholar] [CrossRef]
- Cogalniceanu, D.; Rosioru, D.; Szekely, P.; Szekely, D.; Buhaciuc, E.; Stanescu, F.; Miaud, C. Age and Body Size in Populations of Two Syntopic Spadefoot Toads (Genus pelobates) at the Limit of Their Ranges. J. Herpetol. 2014, 48, 537–545. [Google Scholar] [CrossRef]
- Ma, J.; Yuan, Z.; Hu, Q.; Liu, Q. Statistical analysis of quantitive character of two categories of paramesotriton in Guangxi. J. Guangxi Med. Univ. 1999, 16, 131–133. [Google Scholar]
- Xie, C.; Wang, X.; Chen, L.; Li, X.; Tang, C.; Liu, F.; Yang, J. Sexual Size Dimorphism and Character Scaling of Cynops orientalis. Sichuan J. Zool. 2021, 30, 772–776. [Google Scholar]
- Hernández-Salinas, U.; Ramírez-Bautista, A.; Stephenson, B.P.; Cruz-Elizalde, R.; Berriozabal-Islas, C.; Balderas-Valdivia, C.J. Amphibian life history in a temperate environment of the Mexican Plateau: Dimorphism, phenology and trophic ecology of a hylid frog, Hyla eximia (=Dryophytes eximius). PeerJ 2018, 6, e5897. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Arcos, V.H.; Calzada-Arciniega, R.A.; Alfaro-Juantorena, L.A.; Vazquez-Reyes, L.D.; Blair, C.; Parra-Olea, G. A new species of Charadrahyla (Anura: Hylidae) from the cloud forest of western Oaxaca, Mexico. Zootaxa 2019, 4554, 371–385. [Google Scholar] [CrossRef]
- Caldart, V.M.; Loebens, L.; Brum, A.J.C.; Bataioli, L.; Cechin, S.Z. Reproductive Cycle, Size and Age at Sexual Maturity, and Sexual Dimorphism in the Stream-Breeding Frog Crossodactylus schmidti (Hylodidae). South Am. J. Herpetol. 2019, 14, 1–11. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, J.; Feng, L.; Zheng, W. Sexual Dimorphism of Beilun Pygmy Frog (Microhyla beilunensis) from Jiulongshan Mountain, Zhejiang. Chin. J. Zool. 2020, 55, 178–188. [Google Scholar] [CrossRef]
- Long, J.; Hou, J.; Hu, Y.; Xiang, J.; Pan, W. Sexual Dimorphism and Fecundity of Quasipaa spinosa. Chin. J. Wildl. 2021, 42, 817–824. [Google Scholar]
- Zhao, H.; Wang, Q.; Jiang, W.; Deng, J.; Kong, F.; Zhang, H. Measurement and analysis of morphological features of the Batrachuperus taibaiensis, Jiyinzuxue Yu Yingyong Shengwuxue. Genom. Appl. Biol. 2017, 36, 622–629. [Google Scholar]
- Tao, X.; Jiang, Z.; Ji, S.; Chu, H.; Li, N.; Yang, D.; Li, C. The relationship between body condition index and ecological factors in two Phrynocephalus species in the piedmont desert of Altai Mountains. Biodivers. Sci. 2020, 28, 412–421. [Google Scholar] [CrossRef]
- Xiong, J.; Liu, X.; Zhang, X.; Li, M.; Min, Y. Sexual Dimorphism of the Jilin Clawed Salamander, Onychodactylus zhangyapingi,(Urodela: Hynobiidae: Onychodactylinae) from Jilin Province, China. Asian Herpetol. Res. 2016, 7, 220–226. [Google Scholar]
- Ethier, J.P.; Fayard, A.; Soroye, P.; Choi, D.; Mazerolle, M.J.; Trudeau, V.L. Life history traits and reproductive ecology of North American chorus frogs of the genus Pseudacris (Hylidae). Front. Zool. 2021, 18, 40. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.; Hong, Y.; Wu, A.; Zhou, G.; Tang, C. Effect of different reproductive modes on the reproductive effect of Andrias davidianus. Hebei Fish. 2019, 10, 26–28, 33. [Google Scholar]
- Gong, D.; Qin, W.; Guo, X.; Sun, C.; Li, X. Observation and anaylsis of reproductive behavior of Batrachuperus tibetanus in Jishishan county. J. Northwest Norm. Univ. (Nat. Sci.) 2013, 49, 83–88. [Google Scholar]
- Souza, J.R.D.; Kaefer, I.L.; Lima, A.P. The peculiar breeding biology of the Amazonian frog Allobates subfolionidificans (Aromobatidae). An. Da Acad. Bras. De Cienc. 2017, 89, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.K.; Okada, S.; Fukuda, Y. From embryos to larvae: Seven-month-long paternal care by male Japanese giant salamander. J. Zool. 2017, 302, 24–31. [Google Scholar] [CrossRef]
- Zhang, X.; Xiong, J.L.; Lv, Y.Y.; Zhang, L.; Sun, Y.Y. Sexual size and shape dimorphism in the Wushan salamander, Liua shihi (Liu, 1950) (Urodela: Hynobiidae). Ital. J. Zool. 2014, 81, 368–373. [Google Scholar] [CrossRef]
- Marvin, G.A. Sexual and Seasonal Dimorphism in the Cumberland Plateau Woodland Salamander, Plethodon kentucki (Caudata: Plethodontidae). Copeia 2009, 2009, 227–232. [Google Scholar] [CrossRef]
- Schoener, T.W.; Slade, J.B.; Stinson, C.H. Diet and sexual dimorphism in the very catholic lizard genus, Leiocephalus of the Bahamas. Oecologia 1982, 53, 160–169. [Google Scholar] [CrossRef]
- Bakkegard, K.A.; Rhea, R.A. Tail Length and Sexual Size Dimorphism (SSD) in Desmognathan Salamanders. J. Herpetol. 2012, 46, 304–311. [Google Scholar] [CrossRef]
- Kakegawa, M.; Kishi, F.; Saikawa, Y.; Hasumi, M. Seasonal changes in body shape and mass in a lotic-breeding and externally fertilizing salamander Hynobius kimurae. Zool. Anzeiger. 2017, 268, 55–63. [Google Scholar] [CrossRef]
- Malmgren, J.C.; Thollesson, M. Sexual size and shape dimorphism in two species of newts, Triturus cristatus and T. vulgaris (Caudata: Salamandridae). J. Zool. 1999, 249, 127–136. [Google Scholar] [CrossRef]
- Bovero, S.; Sotgiu, G.; Castellano, S.; Giacoma, C. Age and sexual dimorphism in a population of Euproctus platycephalus (Caudata: Salamandridae) from Sardinia. Copeia 2003, 2003, 149–154. [Google Scholar] [CrossRef]
- Bu, R. Research of Reproductive Behavior and the Habitat Selection of Breeding Period Hynobius maoershanensis. Master Thesis, Guangxi Normal University, Guilin, China, 2017. [Google Scholar]
- Howard, R.D.; Kluge, A.G. Proximate mechanisms of sexual selection in wood frogs. Evolution 1985, 39, 260–277. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
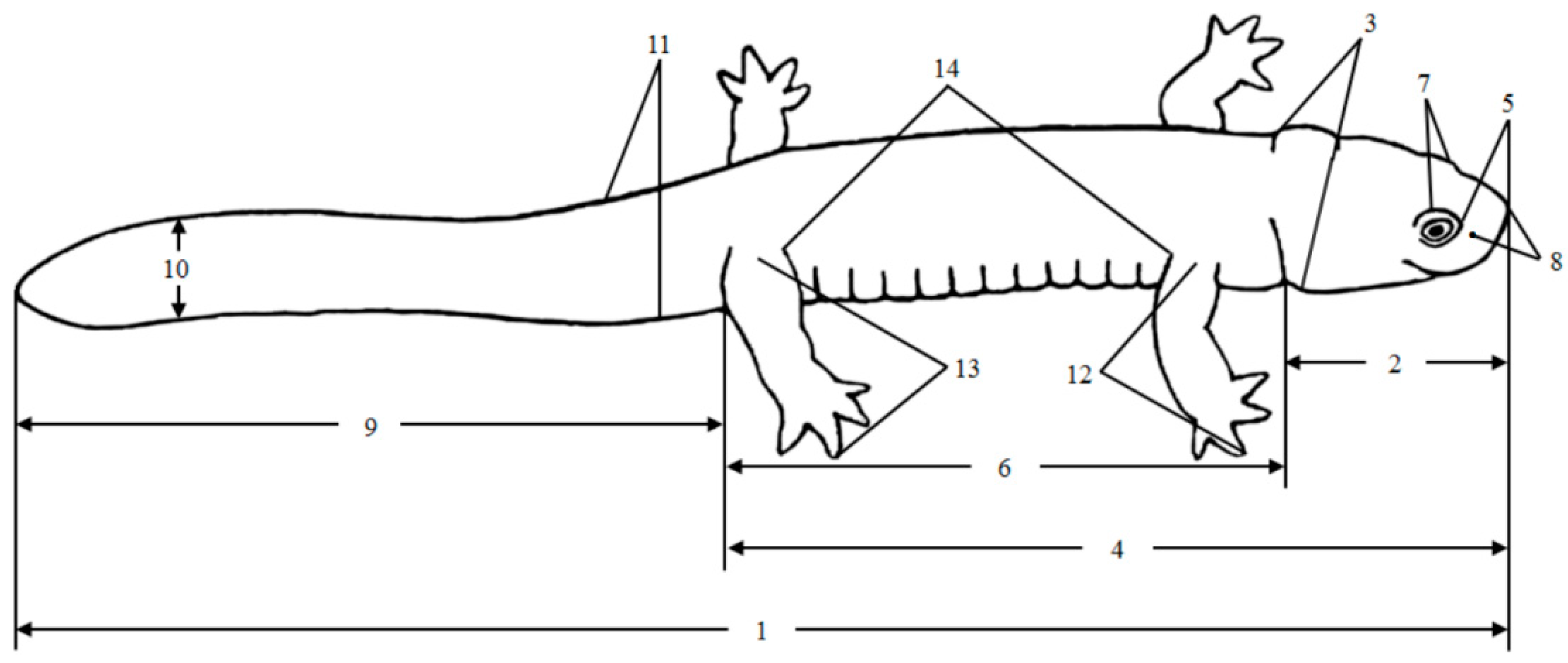
| Morphological Variables | Definition |
|---|---|
| 1. Total length (TOL) | From the snout to the tail end |
| 2. Head length (HL) | From the rostral end to the posterior edge of the jaw joint |
| 3. Head width (HW) | Width of the head at its widest point |
| 4. Snout–vent length (SVL) | From the tip of the snout to the posterior margin of the cloaca |
| 5. Snout length (SL) | From the anterior border of the eye to the tip of the snout |
| 6. Trunk length (TRL) | Length from the neck drape to the posterior edge of the anal hole |
| 7. Interorbital space (IOS) | The narrowest distance between the inner margins of the left and right upper eyelids |
| 8. Internasal space (INS) | The distance between the two nostrils |
| 9. Tail length (TL) | From the posterior margin of the cloaca to the tip of the tail |
| 10. Tail height (TH) | The height of the tail at its highest point |
| 11. Tail width (TW) | The width of the tail at its widest point |
| 12. Forelimb length (FLL) | Length from the base of the forelimb to the longest finger end |
| 13. Hindlimb length (HLL) | Length from the base of the hindlimb to the end of the longest toe |
| 14. Space between axilla and groin (AGS) | Distance from the posterior edge of the forelimb to the anterior edge of the hindlimb |
| Variables | Hynobius maoershanensis | ||
|---|---|---|---|
| Female (n = 9) | Male (n = 52) | p-Value | |
| Mean ± SE | mean ± SE | ||
| SVL | 88.61 ± 2.28 | 89.13 ± 0.71 | - |
| TOL | 160.18 ± 5.63 | 167.68 ± 1.83 | <0.001 ** |
| HL | 18.90 ± 1.34 | 19.82 ± 0.60 | 0.003 ** |
| HW | 15.46 ± 0.69 | 17.89 ± 0.36 | <0.001 ** |
| SL | 4.44 ± 0.21 | 4.77 ± 0.10 | 0.061 |
| TRL | 71.47 ± 2.58 | 70.52 ± 0.81 | <0.001 ** |
| IOS | 5.22 ± 0.23 | 5.74 ± 0.13 | 0.246 |
| INS | 5.24 ± 0.27 | 5.42 ± 0.07 | <0.001 ** |
| TL | 70.12 ± 3.69 | 78.32 ± 1.28 | <0.001 ** |
| TH | 9.81 ± 0.45 | 11.74 ± 0.22 | <0.001 ** |
| TW | 6.61 ± 0.34 | 7.55 ± 0.26 | <0.001 ** |
| FLL | 22.24 ± 0.51 | 25.62 ± 0.29 | 0.019 * |
| HLL | 25.74 ± 0.36 | 28.08 ± 0.31 | 0.007 ** |
| AGS | 46.17 ± 1.84 | 43.02 ± 0.63 | <0.001 ** |
| W | 20.64 ± 2.36 | 22.11 ± 0.65 | <0.001 ** |
| Variables | F-Value | p-Value | Sex Bias |
|---|---|---|---|
| TOL | 4.743 | 0.033 | M |
| HW | 8.812 | 0.004 | M |
| TRL | 0.557 | 0.459 | n.s |
| INS | 0.693 | 0.409 | n.s |
| TL | 7.250 | 0.009 | M |
| TH | 15.497 | 0.0001 | M |
| TW | 2.533 | 0.117 | n.s |
| FLL | 22.297 | 0.00001 | M |
| HLL | 9.616 | 0.003 | M |
| AGS | 6.796 | 0.012 | F |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Bu, R.; Ning, M.; Yang, B.; Wu, Z.; Huang, H. Sexual Dimorphism in the Chinese Endemic Species Hynobius maoershanensis (Urodela: Hynobiidae). Animals 2022, 12, 1712. https://doi.org/10.3390/ani12131712
Chen H, Bu R, Ning M, Yang B, Wu Z, Huang H. Sexual Dimorphism in the Chinese Endemic Species Hynobius maoershanensis (Urodela: Hynobiidae). Animals. 2022; 12(13):1712. https://doi.org/10.3390/ani12131712
Chicago/Turabian StyleChen, Huiqun, Rongping Bu, Meihong Ning, Bo Yang, Zhengjun Wu, and Huayuan Huang. 2022. "Sexual Dimorphism in the Chinese Endemic Species Hynobius maoershanensis (Urodela: Hynobiidae)" Animals 12, no. 13: 1712. https://doi.org/10.3390/ani12131712
APA StyleChen, H., Bu, R., Ning, M., Yang, B., Wu, Z., & Huang, H. (2022). Sexual Dimorphism in the Chinese Endemic Species Hynobius maoershanensis (Urodela: Hynobiidae). Animals, 12(13), 1712. https://doi.org/10.3390/ani12131712