
Effects of Raw and Pasteurized Camel Milk on Metabolic Responses in Pigs Fed a High-Fat Diet

 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
- Low fat diet (control)
- High fat diet (HF) (the main source of fat was tallow)
- High fat diet + 500 mL/per day Raw Camel Milk (CM)
- High fat diet + 500 mL/per day 63 °C Pasteurized Camel Milk (CM)
2.1. Measurements
2.1.1. Macrocompositional Analysis of CM
2.1.2. Liveweight, Backfat, and Feed Intake
2.1.3. Blood Sampling
2.1.4. In Vitro Glucose Tolerance Test (IVGTT)
2.2. Laboratory Analysis
2.3. Metabolic Challenges Data Calculations
- CR = [(ln [ta] − ln [tb])/(tb-ta)] × 100, where [ta] is the concentration of the metabolite at time a (ta) and [tb] is the concentration of metabolite at time b (tb).
- Tbasal glucose = [(ln [2 min] − ln [240 min])/CR2-30 glucose] × 100
- Tbasal insulin = [(ln [2 min] − ln [240 min])/CR20-120 insulin] × 100
- Increment = peak concentration − basal concentration
- Decrement = nadir concentration − basal concentration.
- Change = [(peak (or nadir) concentration − basal concentration)/basal concentration] × 100
2.4. MINMOD Parameters
- GB = basal glucose concentration pre-infusion (mM)
- IB = basal insulin concentration pre-infusion (mU/L)
- SG = glucose effectiveness (min−1), which refers to the capacity of glucose to mediate its own uptake
- SI = insulin sensitivity ((mU/L)−1.min−1), which refers to the capacity of insulin to promote glucose uptake
- AIRg = acute insulin response to glucose (mIU/L−1.min−1), which addresses the adequacy of insulin secretion in response to a glucose bolus
- DI = disposition index (AIR × SI), which represents the ability of the islet cells to secrete insulin
- G0 = distributed glucose concentration at time 0
- GEZI = glucose effectiveness at zero insulin
- Other indices that are included in the MINMOD Millennium output are derived from the Homeostatic assessment model (HOMA), as follows:
- IR = insulin resistance (mM/mU/L−2), calculated by the equation: (IB × GB)/22.5
- Β-cell function (BCF) = pancreatic β-cell function (mU/mM), calculated by the equation: (20 × IB)/(GB-3.5)
2.5. Statistical Analyses
3. Results
3.1. Growth, Feed Intake, and Plasma Metabolite Responses
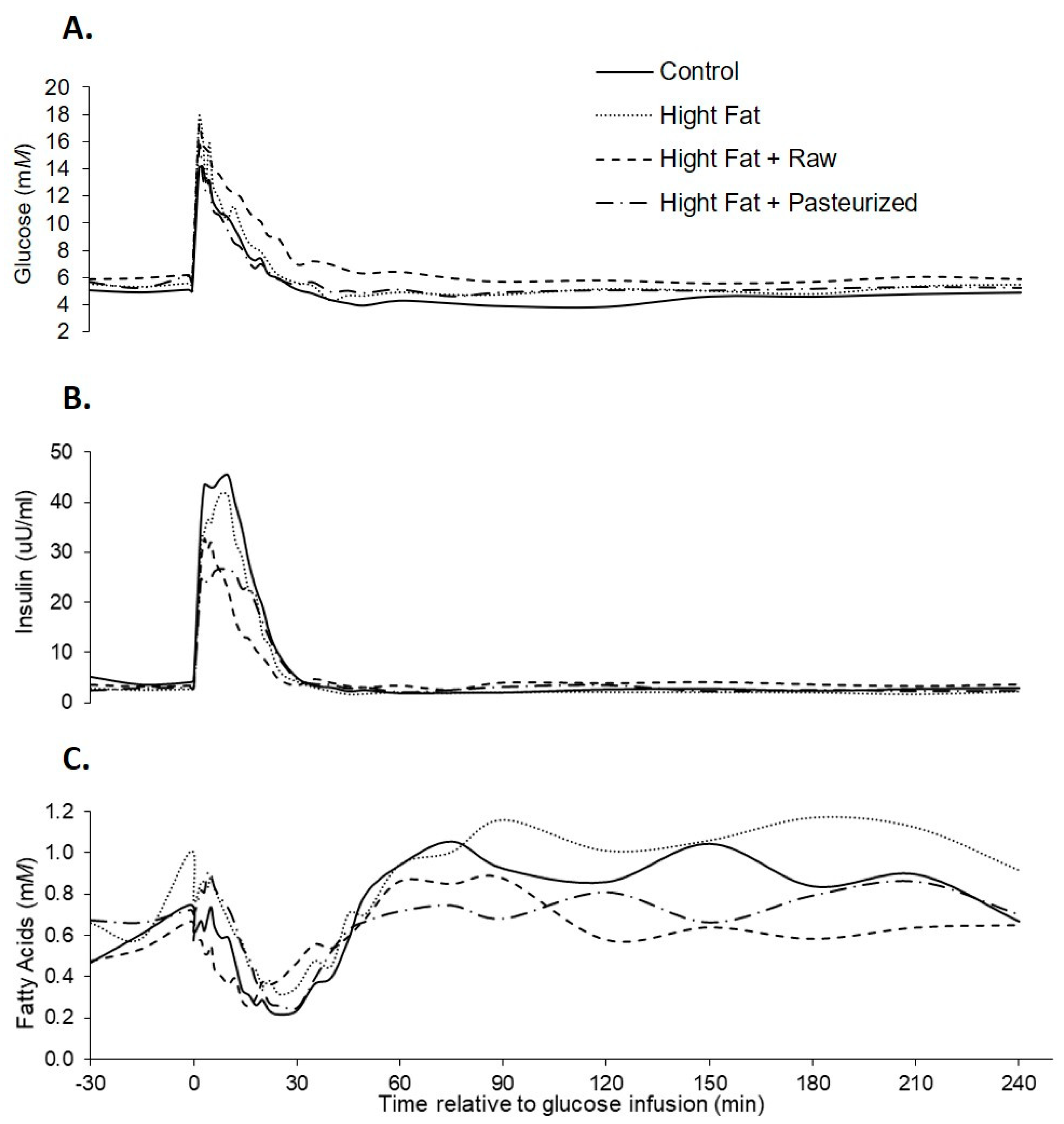
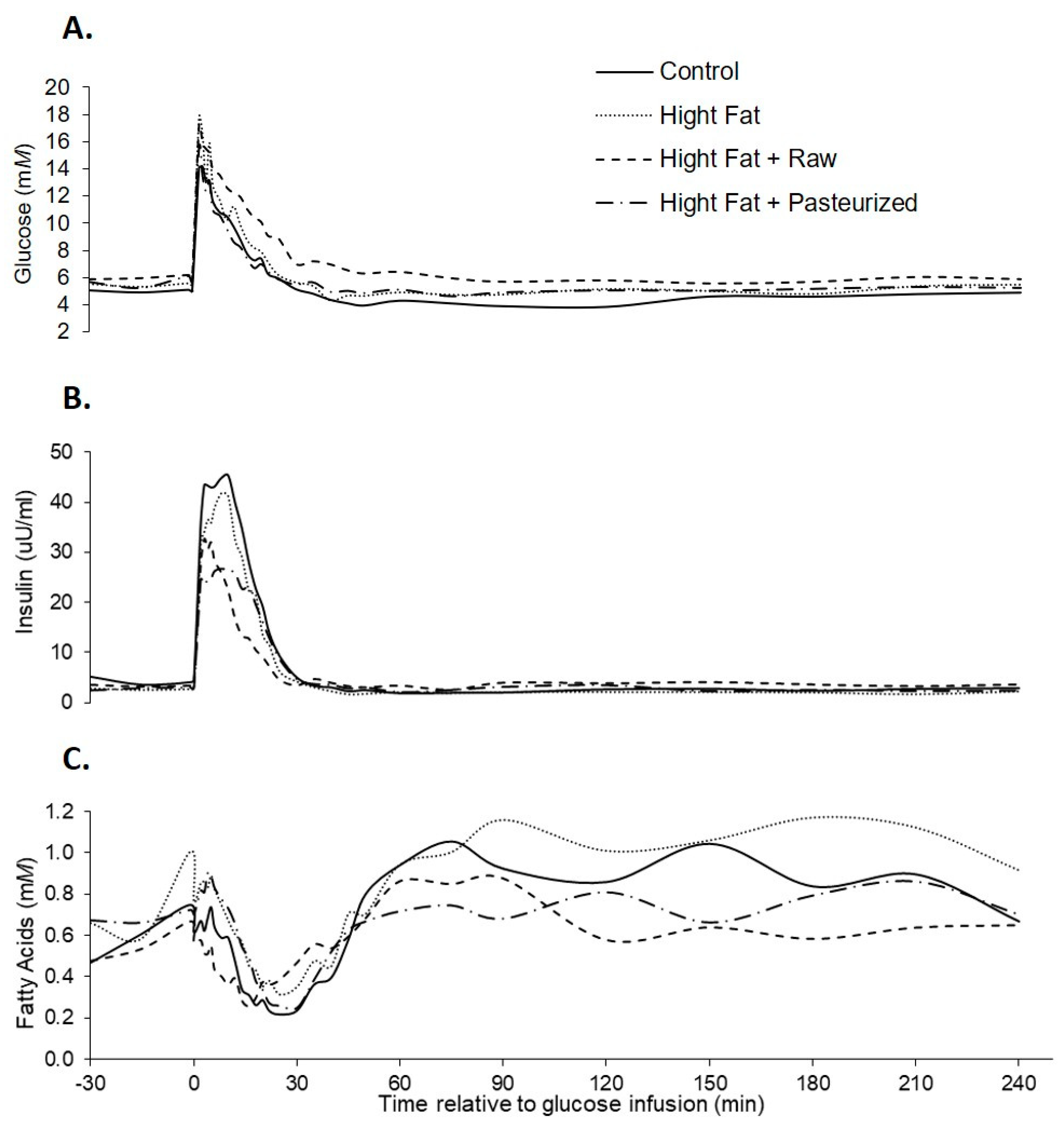
3.2. IVGTT Responses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Izadi, A.; Khedmat, L.; Mojtahedi, S.Y. Nutritional and therapeutic perspectives of camel milk and its protein hydrolysates: A review on versatile biofunctional properties. J. Funct. Foods 2019, 60, 103441. [Google Scholar] [CrossRef]
- Hamad, E.M. Beneficial effect of camel milk on liver and kidneys function in diabetic Sprague-Dawley rats. Int. J. Dairy. Sci. 2011, 6, 190–197. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, R.P.; Jain, S.; Shah, S.; Chopra, A.; Agarwal, V. Effect of camel milk on glycemic control and insulin requirement in patients with type 1 diabetes: 2-years randomized controlled trial. Eur. J. Clin. Nutr. 2011, 65, 1048–1052. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, R.P.; Kochar, D.K.; Sahani, M.S.; Tuteja, F.C.; Ghorui, S.K. Hypoglycemic activity of camel milk in streptozotocin induced diabetic rats. Int. J. Diabetes Dev. Ctries. 2004, 24, 47–49. [Google Scholar]
- Mirmiran, P.; Ejtahed, H.-S.; Angoorani, P.; Eslami, F.; Azizi, F. Camel milk has beneficial effects on diabetes mellitus: A systematic review. Int. J. Endocrinol. Metab. 2017, 15, e42150. [Google Scholar] [CrossRef] [Green Version]
- Hussain, H.; Wattoo, F.H.; Wattoo, M.H.S.; Gulfraz, M.; Masud, T.; Shah, I.; Ali, S.; Alavi, S.E. Camel milk as an alternative treatment regimen for diabetes therapy. Food Sci. Nutr. 2021, 9, 1347–1356. [Google Scholar] [CrossRef]
- Korish, A.A.; Abdel Gader, A.G.M.; Alhaider, A.A. Comparison of the hypoglycemic and antithrombotic (anticoagulant) actions of whole bovine and camel milk in streptozotocin-induced diabetes mellitus in rats. J. Dairy Sci. 2020, 103, 30–41. [Google Scholar] [CrossRef] [Green Version]
- Sangild, P.T. Gut responses to enteral nutrition in preterm infants and animals. Exp. Biol. Med. 2006, 231, 1695–1711. [Google Scholar] [CrossRef]
- Spurlock, M.E.; Gabler, N.K. The development of porcine models of obesity and the metabolic syndrome. J. Nutr. 2008, 138, 397–402. [Google Scholar] [CrossRef]
- Ou, J.Z.; Cottrell, J.J.; Ha, N.; Pillai, N.; Yao, C.K.; Berean, K.J.; Ward, S.A.; Grando, D.; Muir, J.G.; Harrison, C.J.; et al. Potential of in vivo real-time gastric gas profiling: A pilot evaluation of heat-stress and modulating dietary cinnamon effect in an animal model. Sci. Rep. 2016, 6, 33387. [Google Scholar] [CrossRef]
- National Research Council, NRC. Nutrient Requirements of Swine, 11th Revised ed.; The National Academies Press: Washington, DC, USA, 2012; p. 420. [Google Scholar]
- Liu, F.; Cottrell, J.J.; Wijesiriwardana, U.; Kelly, F.W.; Chauhan, S.S.; Pustovit, R.V.; Gonzales-Rivas, P.A.; DiGiacomo, K.; Leury, B.J.; Celi, P.; et al. Effects of chromium supplementation on physiology, feed intake, and insulin related metabolism in growing pigs subjected to heat stress. Trans. Anim. Sci. 2017, 1, 116–125. [Google Scholar]
- Johnson, M.M.; Peters, J.P. Technical note: An improved method to quantify nonesterified fatty acids in bovine plasma. J. Anim. Sci. 1993, 71, 753–756. [Google Scholar] [CrossRef]
- Pires, J.A.A.; Souza, A.H.; Grummer, R.R. Induction of Hyperlipidemia by Intravenous Infusion of Tallow Emulsion Causes Insulin Resistance in Holstein Cows. J. Dairy Sci. 2007, 90, 2735–2744. [Google Scholar] [CrossRef] [Green Version]
- Boston, R.C.; Stefanovski, D.; Moate, P.J.; Sumner, A.E.; Watanabe, R.M.; Bergman, R.N. MINMOD Millennium: A computer program to calculate glucose effectiveness and insulin sensitivity from the frequently sampled intravenous glucose tolerance test. Diabetes Technol. Ther. 2003, 5, 1003–1015. [Google Scholar] [CrossRef] [Green Version]
- Payne, R.W.; Harding, S.A.; Murray, D.A.; Soutar, D.M.; Baird, D.B.; Welham, S.J.; Kane, A.F.; Gilmour, R.; Thompson, R.; Webster, R.; et al. GenStat Release 11 Reference Manual; VSN International: Hertfordshire, UK, 2008. [Google Scholar]
- Cottrell, J.J.; Furness, J.B.; Wijesiriwardana, U.A.; Ringuet, M.; Liu, F.; DiGiacomo, K.; Leury, B.J.; Clarke, I.; Dunshea, F.R. The effect of heat stress on respiratory alkalosis and insulin sensitivity in cinnamon supplemented pigs. Animals 2020, 10, 690. [Google Scholar] [CrossRef]
- Dunshea, F.R.; Cox, M.L. Effect of dietary protein on body composition and insulin resistance using a pig model of the child and adolescent. Nutr. Diet. 2008, 65, S60–S65. [Google Scholar] [CrossRef]
- Ashraf, A.; Mudgil, P.; Palakkott, A.; Iratni, R.; Gan, C.-Y.; Maqsood, S.; Ayoub, M.A. Molecular basis of the anti-diabetic properties of camel milk through profiling of its bioactive peptides on dipeptidyl peptidase IV (DPP-IV) and insulin receptor activity. J. Dairy Sci. 2021, 104, 61–77. [Google Scholar] [CrossRef]
- Korish, A.A. The antidiabetic action of camel milk in experimental Type 2 diabetes mellitus: An overview on the changes in incretin hormones, insulin resistance, and inflammatory cytokines. Horm. Metab. Res. 2014, 46, 404–411. [Google Scholar] [CrossRef]
- Zahra, F.; Awat, F.; Mahin, H.; Roya, K. Effect of fermented camel milk on glucose metabolism, insulin resistance, and inflammatory biomarkers of adolescents with metabolic syndrome: A double-blind, randomized, crossover trial. J. Res. Med. Sci. 2018, 23, 32. [Google Scholar]
- Agrawal, R.P.; Saran, S.; Sharma, P.; Gupta, R.P.; Kochar, D.K.; Sahani, M.S. Effect of camel milk on residual β-cell function in recent onset type 1 diabetes. Diabetes Res. Clin. Pract. 2007, 77, 494–495. [Google Scholar] [CrossRef]
- Agrawal, R.P.; Swami, S.C.; Beniwal, R.; Kochar, D.K.; Sahani, M.S.; Tuteja, F.C.; Ghouri, S.P. Effect of camel milk on glycemic control, risk factors and diabetes quality of life in type 1 diabetes: A randomised prospective controlled study. J. Camel Pract. Res. 2003, 10, 45–50. [Google Scholar]
- Alavi, F.; Salami, M.; Emam-Djomeh, Z.; Mohammadian, M. Nutraceutical properties of camel Milk. In Nutrients in Dairy and their Implications on Health and Disease; Watson, R.R., Collier, R.J., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 451–468. [Google Scholar]
- De Meyts, P. The Insulin Receptor and Its Signal Transduction Network. In Endotext [Internet]; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., Dungan, K., Grossman, A., Hershman, J.M., Kaltsas, G., Koch, C., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Abdulrahman, A.; Ismel, M.A.; Al-Hosaini, K.; Rame, C.; Al-Senaidy, A.M.; Dupont, J.; Ayoub, M.A. Differential effects of camel milk on insulin receptor signaling—Toward understanding the insulin-like properties of camel milk. Front. Endocrinol. 2016, 7, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darwish, H.A.; Abd Raboh, N.R.; Mahdy, A. Camel’s milk alleviates alcohol-induced liver injury in rats. Food Chem. Toxicol. 2012, 50, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.I.; Bayoumi, M.M. Efficiency of camel milk and honey bee in alleviation of diabetes in rats. Nat. Sci. 2010, 8, 333–341. [Google Scholar]
- Tomas, F.M.; Walton, P.E.; Dunshea, F.R.; Ballard, F.J. IGF-I variants which bind poorly to IGF-binding proteins show more potent and prolonged hypoglycaemic action than native IGF-I in pigs and marmoset monkeys. J. Endocrinol. 1997, 155, 377–386. [Google Scholar] [CrossRef]
- Devedjian, J.C.; George, M.; Casellas, A.; Pujol, A.; Visa, J.; Pelegrín, M.; Gros, L.; Bosch, F. Transgenic mice overexpressing insulin-like growth factor-II in beta cells develop type 2 diabetes. J. Clin. Investig. 2000, 105, 731–740. [Google Scholar] [CrossRef] [Green Version]
- Casellas, A.; Mallol, C.; Salavert, A.; Jimenez, V.; Garcia, M.; Agudo, J.; Obach, M.; Haurigot, V.; Vilà, L.; Molas, M.; et al. Insulin-like growth factor 2 overexpression induces β-cell dysfunction and increases β-cell susceptibility to damage. J. Biol. Chem. 2015, 290, 16772–16785. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
| Control | High Fat | |
|---|---|---|
| DM (%) | 90.7 | 91.6 |
| DE (MJ/kg) | 14.2 | 17.4 |
| CP (%) | 18.4 | 18.4 |
| Fat (%) | 2.7 | 16.2 |
| Starch (%) | 56.3 | 44.5 |
| Fiber (%) | 2.4 | 2.0 |
| Control | High Fat | Camel Milk | SED | p-Value | |
|---|---|---|---|---|---|
| Glucose (mM) | 0.816 | 0.842 | 0.863 | 0.0242 | 0.15 |
| (6.55) | (6.95) | (7.30) | - | ||
| Insulin (uU/mL) | 0.680 | 0.697 | 0.681 | 0.1173 | 0.99 |
| (4.79) | (4.98) | (4.80) | - | ||
| Fatty acids (mM) | 2.068 a | 2.29 b | 2.33 c | 0.05958 | <0.001 |
| (117) | (195) | (212) | - | ||
| Urea (mM) | 0.912 | 0.790 | 0.878 | 0.1324 | 0.68 |
| (8.17) | (6.17) | (7.56) | - | ||
| Cholesterol Mm | 0.0547 a | 0.161 b | 0.195 c | 0.03682 | 0.003 |
| (1.13) | (1.45) | (1.57) | - | ||
| IL-10 pg/mL | 2.09 | 1.92 | 2.00 | 0.1686 | 0.68 |
| (122) | (83.9) | (99.5) | - | ||
| IGF1 ng/mL | 1.41 | 1.40 | 1.43 | 0.02807 | 0.49 |
| (25.8) | (25.0) | (26.9) | - | ||
| IGF-2 ng/mL | 1.61 | 1.61 | 1.58 | 0.02878 | 0.44 |
| (41.1) | (40.8) | (38.4) | - | ||
| IL-6 pg/mL | 2.81 | 3.34 | 2.56 | 0.4377 | 0.19 |
| (640) | (2208) | (362) | - | ||
| Glucagon pg/mL | 2.39 | 2.21 | 2.52 | 0.1897 | 0.23 |
| (248) | (162) | (334) | - | ||
| GLP1 ng/mL | 1.27 | 1.25 | 1.14 | 0.07736 | 0.17 |
| (18.5) | (17.8) | (13.6) | - | ||
| TNFa pg/mL | 1.88 | 2.97 | 1.94 | 0.2822 | 0.93 |
| (75.9) | (923) | (87.7) | - | ||
| C-Pep. ng/mL | 0.441 | 0.616 | 0.593 | 0.09457 | 0.18 |
| (2.76) | (4.13) | (3.92) |
| Control | High Fat | Camel Milk | SED | p-Value | ||
|---|---|---|---|---|---|---|
| Base (mM) | Glucose | 5.03 | 5.51 | 5.81 | 0.603 | 0.28 |
| Insulin | 4.94 | 3.09 | 3.46 | 1.199 | 0.22 | |
| NEFA | 0.649 | 0.723 | 0.663 | 0.1371 | 0.83 | |
| Peak (mM) | Glucose | 14.7 | 17.7 | 16.6 | 2.579 | 0.46 |
| Insulin | 57.9 | 44.6 | 33.4 | 12.04 | 0.058 | |
| NEFA | 1.32 | 1.39 | 1.13 | 0.1744 | 0.18 | |
| Nadir (mM) | Glucose | 3.03 a | 4.22 b | 4.73 c | 0.606 | 0.009 |
| Insulin | 1.69 | 1.22 | 2.28 | 1.137 | 0.38 | |
| NEFA | 0.177 | 0.256 | 0.248 | 0.0520 | 0.23 | |
| AUC 0–30 (mM.min) | Glucose | 94 | 88 | 119 | 45.0 | 0.63 |
| Insulin | 772 | 508 | 425 | 192.1 | 0.097 | |
| NEFA | −7.3 | −6.9 | 4.3 | 3.74 | 0.52 | |
| CR 2–30 (%/min) | Glucose | 3.90 | 3.24 | 3.65 | 0.6860 | 0.61 |
| Insulin | 7.25 | 7.05 | 6.67 | 1.806 | 0.91 | |
| NEFA | 3.75 | 3.33 | 2.48 | 1.859 | 0.65 | |
| Increment from basal (mM) | Glucose | 9.66 | 12.2 | 10.8 | 2.315 | 0.54 |
| Insulin | 52.9 | 41.5 | 30.0 | 11.23 | 0.055 | |
| NEFA | 0.668 | 0.671 | 0.470 | 0.1382 | 0.12 | |
| Decrement from basal (mM) | Glucose | 2.00 | 1.30 | 1.09 | 0.5680 | 0.15 |
| Insulin | 3.25 a | 1.87 b | 0.980 b | 0.9700 | 0.028 | |
| NEFA | 0.472 | 0.466 | 0.425 | 0.1310 | 0.88 |
| Control | High Fat | Camel Milk | SED | p-Value | |
|---|---|---|---|---|---|
| Insulin baseline (mU/L) | 4.89 | 3.05 | 3.50 | 1.0050 | 0.22 |
| Glucose baseline (mg/dL) | 86.5 | 94.6 | 101 | 8.762 | 0.21 |
| AIRg (mU.L−1.min) | 372 | 274 | 223 | 62.80 | 0.059 |
| β-cell function (mU/mM) | 84.9 a | 35.7 b | 33.8 b | 15.48 | 0.006 |
| DI | 9312 | 5346 | 13,078 | 9915.0 | 0.72 |
| GEZI (min−1) | 0.023 | 0.027 | 0.041 | 0.0171 | 0.44 |
| G0 (mg/dL) | 252 | 271 | 295 | 29.38 | 0.29 |
| Insulin Resistance (mM.mU/L−2) | 1.03 | 0.745 | 0.916 | 0.2963 | 0.68 |
| SI ((mU/L)−1.min−1) | 30.7 | 18.4 | 65.9 | 53.21 | 0.60 |
| SG (min−1) | 0.0345 | 0.0329 | 0.0637 | 0.02415 | 0.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
DiGiacomo, K.; Zamuner, F.; Sun, Y.; Dunshea, F.R.; Raynes, J.K.; Leury, B.J. Effects of Raw and Pasteurized Camel Milk on Metabolic Responses in Pigs Fed a High-Fat Diet. Animals 2022, 12, 1701. https://doi.org/10.3390/ani12131701
DiGiacomo K, Zamuner F, Sun Y, Dunshea FR, Raynes JK, Leury BJ. Effects of Raw and Pasteurized Camel Milk on Metabolic Responses in Pigs Fed a High-Fat Diet. Animals. 2022; 12(13):1701. https://doi.org/10.3390/ani12131701
Chicago/Turabian StyleDiGiacomo, Kristy, Fernanda Zamuner, Yushu Sun, Frank R. Dunshea, Jared K. Raynes, and Brian J. Leury. 2022. "Effects of Raw and Pasteurized Camel Milk on Metabolic Responses in Pigs Fed a High-Fat Diet" Animals 12, no. 13: 1701. https://doi.org/10.3390/ani12131701
APA StyleDiGiacomo, K., Zamuner, F., Sun, Y., Dunshea, F. R., Raynes, J. K., & Leury, B. J. (2022). Effects of Raw and Pasteurized Camel Milk on Metabolic Responses in Pigs Fed a High-Fat Diet. Animals, 12(13), 1701. https://doi.org/10.3390/ani12131701