Comparison of Pure and Crossbred Japanese Black Steers in Growth Performance and Metabolic Features from Birth to Slaughter at a Spanish Fattening Farm

 , , , ,
, , , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals and Farm
2.3. Evaluation of Weight, Growth Pattern, and Fatness
2.4. Evaluation of Metabolic Status
2.5. Statistical Analyses
3. Results
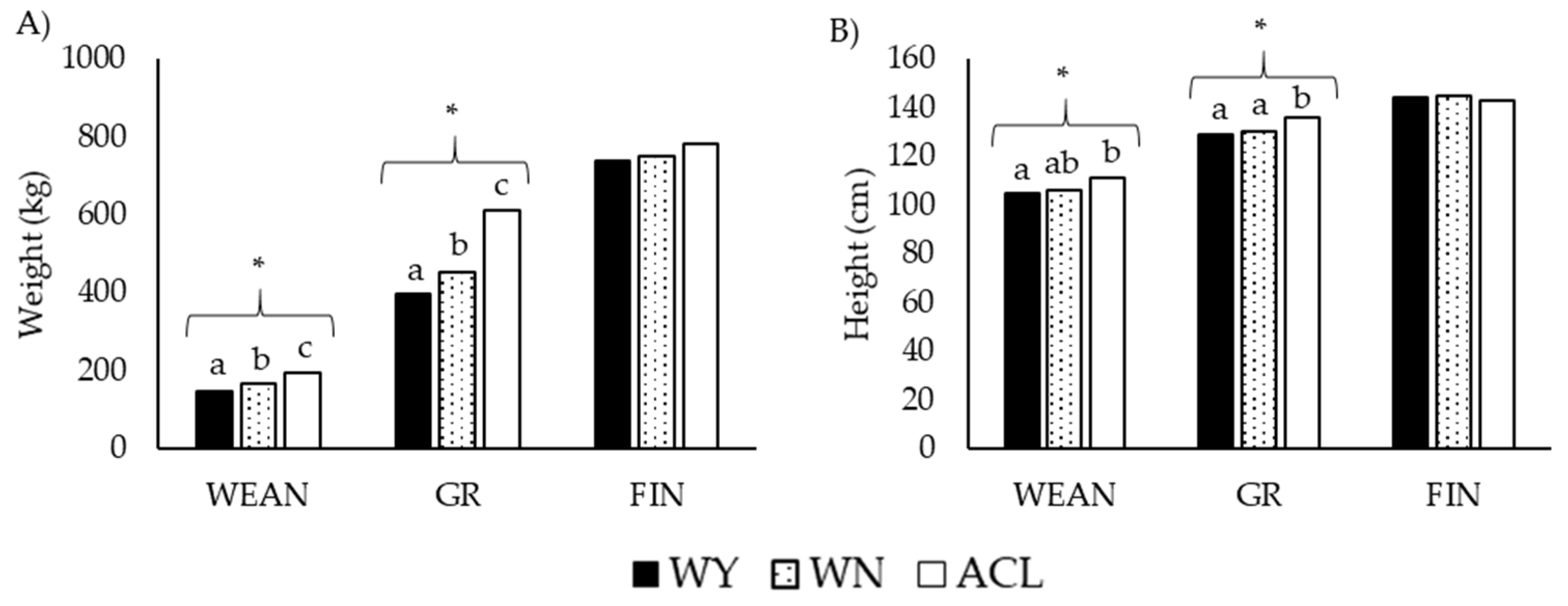
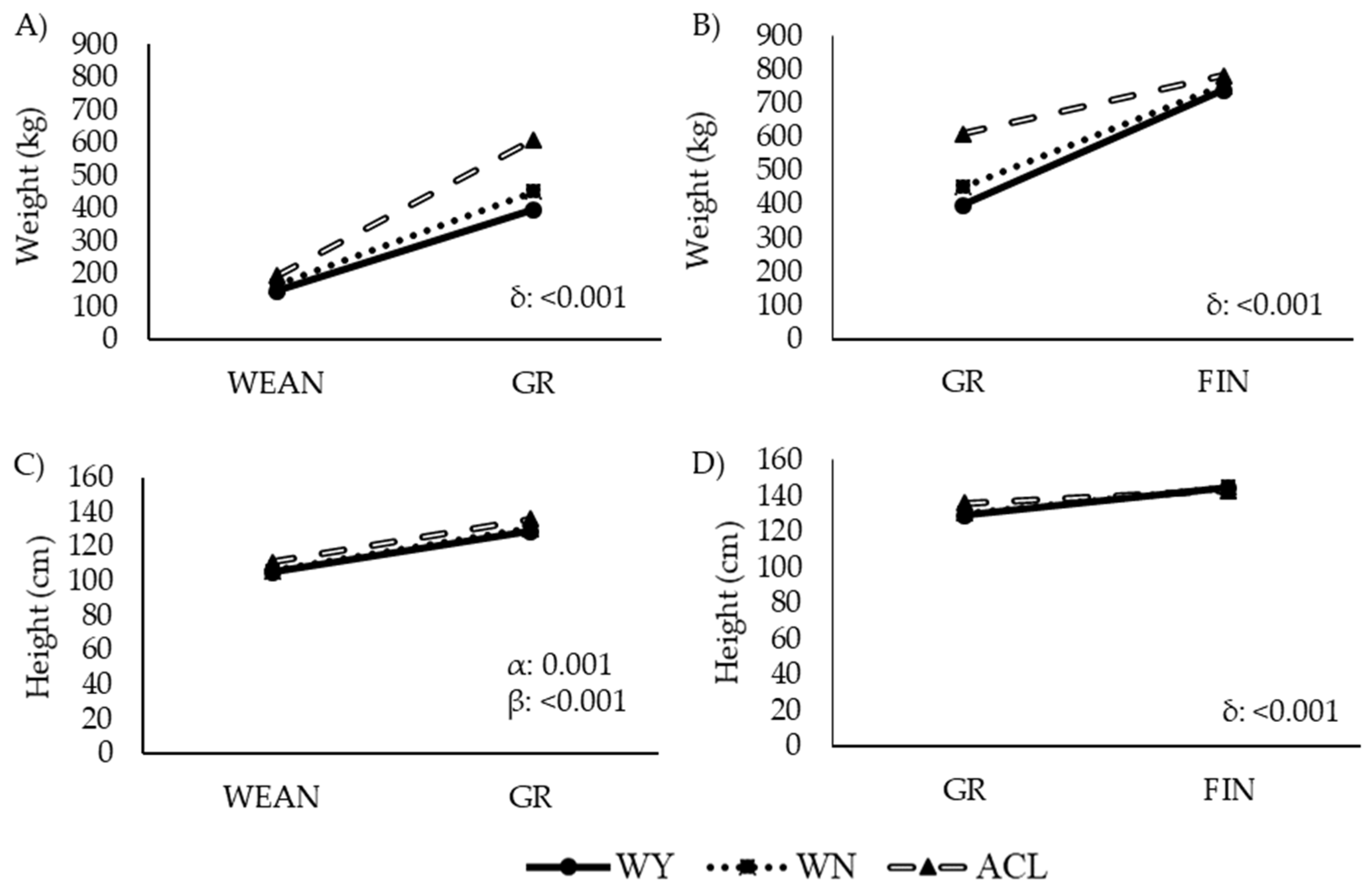
3.1. Growth Pattern (Weight, Height, and Average Daily Gain)
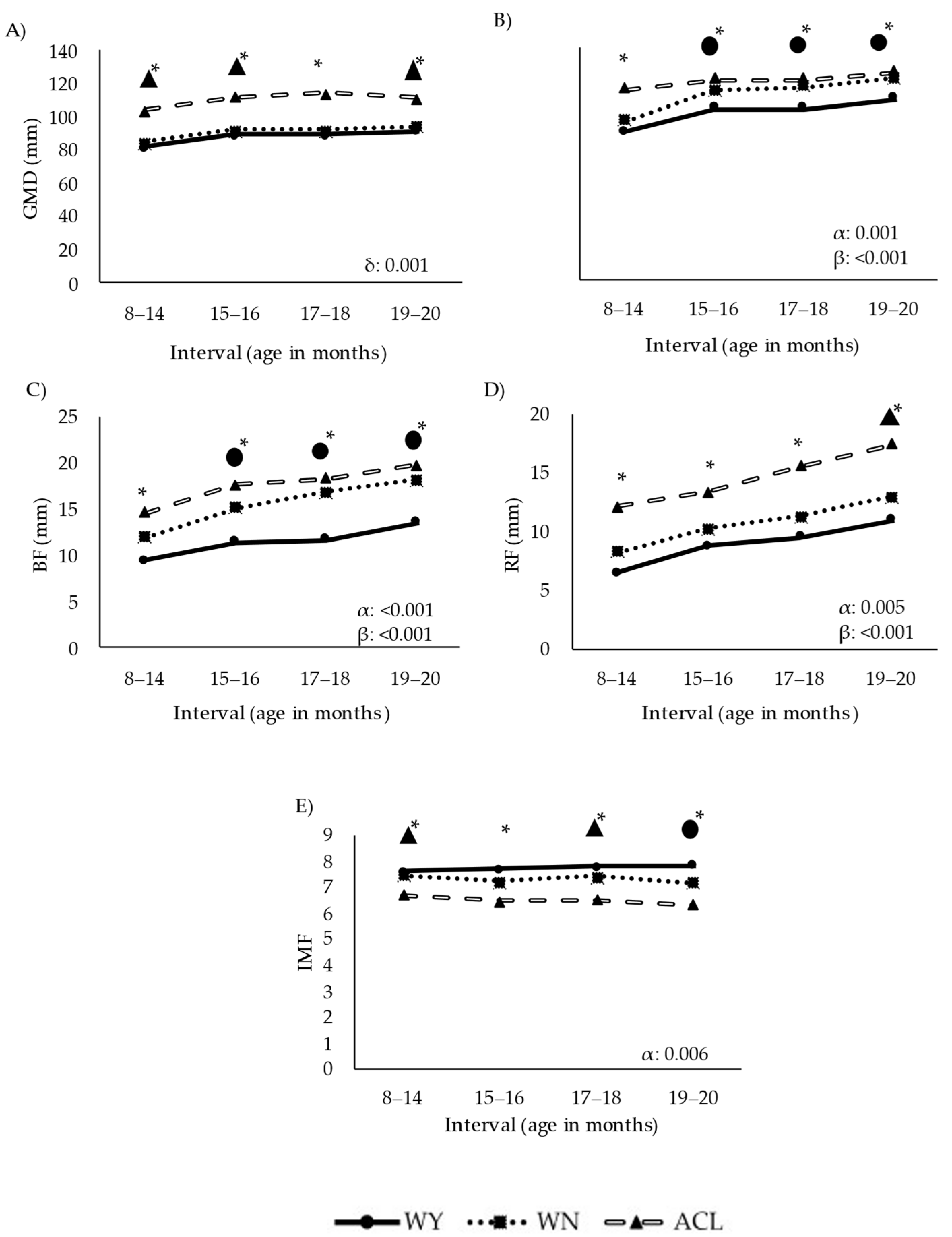
3.2. Ultrasound Evaluation of Fat and Muscle Deposition
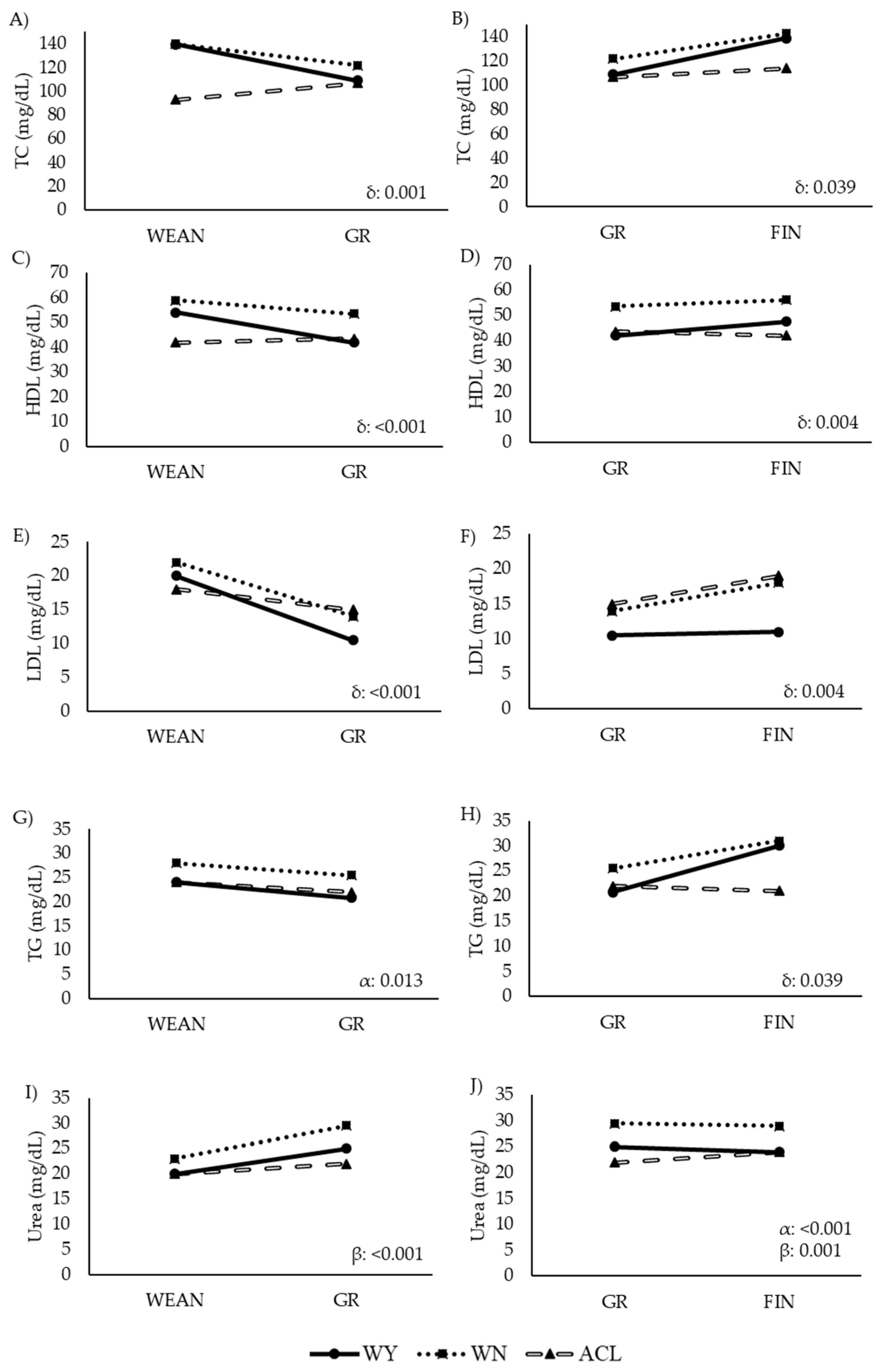
3.3. Metabolic Status
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Albertí, P.; Panea, B.; Sañudo, C.; Olleta, J.; Ripoll, G.; Ertbjerg, P.; Christensen, M.; Gigli, S.; Failla, S.; Concetti, S.; et al. Live weight, body size and carcass characteristics of young steers of fifteen European breeds. Livest. Sci. 2008, 114, 19–30. [Google Scholar] [CrossRef]
- Wheeler, T.L.; Cundiff, L.V.; Koch, M. Effect of marbling degree on beef palatability in Bos taurus and Bos indicus cattle. J. Anim. Sci. Anim. Sci. 2020, 51, 369–378. [Google Scholar] [CrossRef]
- Nishimura, T.; Hattori, A.; Takahashi, K. Structural changes in intramuscular connective tissue during the fattening of Japanese Black cattle: Effect of marbling on beef tenderization. J. Anim. Sci. 1999, 77, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Muir, P.D.; Wallace, G.J.; Dobbie, P.M.; Bown, M.D. A comparison of animal performance and carcass and meat quality characteristics in Hereford, Hereford × Friesian, and Friesian steers grazed together at pasture. N. Z. J. Agric. Res. 2000, 43, 193–205. [Google Scholar] [CrossRef][Green Version]
- Radunz, A.E.; Loerch, S.C.; Lowe, G.D.; Fluharty, F.L.; Zerby, H.N. Effect of Wagyu-versus Angus-sired calves on feedlot performance, carcass characteristics, and tenderness. J. Anim. Sci. 2009, 87, 2971–2976. [Google Scholar] [CrossRef]
- Gotoh, T.; Nishimura, T.; Kuchida, K.; Mannen, H. The Japanese Wagyu beef industry: Current situation and future prospects—A review. Asian-Australas. J. Anim. Sci. 2018, 31, 933–950. [Google Scholar] [CrossRef]
- Park, S.J.; Beak, S.-H.; Jung, D.J.S.; Kim, S.Y.; Jeong, I.H.; Piao, M.Y.; Kang, H.J.; Fassah, D.M.; Na, S.W.; Yoo, S.P.; et al. Genetic, management, and nutritional factors affecting intramuscular fat deposition in beef cattle—A review. Asian-Australas. J. Anim. Sci. 2018, 31, 1043–1061. [Google Scholar] [CrossRef]
- Lunt, D.K.; Riley, R.R.; Smith, S.B. Growth and carcass characteristics of Angus and American Wagyu steers. Meat Sci. 1993, 34, 327–334. [Google Scholar] [CrossRef]
- Mir, P.S.; Bailey, D.R.C.; Mir, Z.; Entz, T.; Jones, S.D.M.; Tson, W.M.R.; Weselake, R.J.; Lozeman, F.J. Growth, carcass and meat quality characteristics of beef cattle with 0, 50 and 75 percent Wagyu genetic influence. Can. J. Anim. Sci. 1999, 79, 129–137. [Google Scholar] [CrossRef]
- Coleman, L.W.; Hickson, R.E.; Schreurs, N.M.; Martin, N.P.; Kenyon, P.R.; Lopez-Villalobos, N.; Morris, S.T. Carcass characteristics and meat quality of Hereford sired steers born to beef-cross-dairy and Angus breeding cows. Meat Sci. 2016, 121, 403–408. [Google Scholar] [CrossRef]
- Nogalski, Z.; Pogorzelska-Przybyłek, P.; Sobczuk-Szul, M.; Nogalska, A.; Modzelewska-Kapituła, M.; Purwin, C. Carcass characteristics and meat quality of steers and steers slaughtered at two different ages. Ital. J. Anim. Sci. 2017, 17, 279–288. [Google Scholar] [CrossRef]
- Nabenishi, H.; Yamazaki, A. Effects of temperature–humidity index on health and growth performance in Japanese black calves. Trop. Anim. Health Prod. 2017, 49, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, J.A.; He, Y.; Li, Y.; Liu, J.; Erdman, R.A.; Sonstegard, T.S.; Song, J. Integrated metabolomic and transcriptome analyses reveal finishing forage affects metabolic pathways related to beef quality and animal welfare. Sci. Rep. 2016, 6, 1–16. [Google Scholar] [CrossRef]
- Takeda, K.I.; Sato, S.; Sugawara, K. Familiarity and group size affect emotional stress in Japanese Black heifers. Appl. Anim. Behav. Sci. 2003, 82, 1–11. [Google Scholar] [CrossRef]
- Chen, S.; Ogura, S. Effects of environmental enrichment on welfare of cattle. J. Integr. Field Sci. 2017, 14, 55–59. [Google Scholar]
- Ninomiya, S. Grooming device effects on behaviour and welfare of Japanese Black fattening cattle. Animals 2019, 9, 186. [Google Scholar] [CrossRef]
- Nakajima, N.; Mitsuishi, H.; Yayota, M. Temperament trait changes in Japanese Black cows under grazing and confined conditions. Front. Vet. Sci. 2021, 8, 1–9. [Google Scholar] [CrossRef]
- May, S.G.; Sturdivant, C.A.; Lunt, D.K.; Miller, R.K.; Smith, S.B. Comparison of sensory characteristics and fatty acid composition between Wagyu crossbred and Angus steers. Meat Sci. 1993, 35, 289–298. [Google Scholar] [CrossRef]
- Greenwood, P.L.; Café, L.M.; Hearnshaw, H.; Hennessy, D.W.; Thompson, J.M.; Morris, S.G. Long-term consequences of birth weight and growth to weaning on carcass, yield and beef quality characteristics of Piedmontese- and Wagyu-sired cattle. Aust. J. Exp. Agric. 2006, 46, 257–269. [Google Scholar] [CrossRef]
- Café, L.M.; Hennessy, D.W.; Hearnshaw, H.; Morris, S.G.; Greenwood, P.L. Influences of nutrition during pregnancy and lactation on birth weights and growth to weaning of calves sired by Piedmontese or Wagyu steers. Aust. J. Exp. Agric. 2006, 46, 245–255. [Google Scholar] [CrossRef]
- Café, L.M.; Hennessy, D.W.; Hearnshaw, H.; Morris, S.G.; Greenwood, P.L. Consequences of prenatal and preweaning growth for feedlot growth, intake and efficiency of Piedmontese- and Wagyu-sired cattle. Anim. Prod. Sci. 2009, 49, 461–467. [Google Scholar] [CrossRef]
- Greenwood, P.L.; Café, L.M.; Hearnshaw, H.; Hennessy, D.W.; Morris, S.G. Consequences of prenatal and preweaning growth for yield of beef primal cuts from 30-month-old Piedmontese-and Wagyu-sired cattle. Anim. Prod. Sci. 2009, 49, 468–478. [Google Scholar] [CrossRef]
- Vann, R.C.; Randel, R.D.; Welsh, T.H.; Willard, S.T.; Carroll, J.A. Evaluation of temperament and transportation stress on body composition traits and meat quality in beef cattle. In Proceedings of the 61st American Meat Science Association Reciprocal Meat Conterence, Gainesville, FL, USA, 22–25 June 2008; pp. 1–5. [Google Scholar]
- Davies, B.L.; Alford, A.R.; Griffith, G.R. Economic effects of alternate growth path, time of calving and breed type combinations across southern Australian beef cattle environments: Industry-wide effects. Anim. Prod. Sci. 2009, 49, 542–547. [Google Scholar] [CrossRef]
- Khounsaknalath, S.; Etoh, K.; Sakuma, K.; Saito, K.; Saito, A.; Abe, T.; Ebara, F.; Sugiyama, T.; Kobayashi, E.; Gotoh, T. Effects of early high nutrition related to metabolic imprinting events on growth, carcass characteristics, and meat quality of grass-fed Wagyu (Japanese Black cattle). J. Anim. Sci. 2021, 99, 1–9. [Google Scholar] [CrossRef]
- Prado, R.; Prado, I.; Marques, J.; Rotta, P.; Visentainer, J.; Silva, R.; Souza, N. Meat quality of the Longissimus muscle of steers and steers (½ Nellore vs. ½ Simmental) finished in feedlot. J. Anim. Feed Sci. 2009, 18, 221–230. [Google Scholar] [CrossRef]
- Toušová, R.; Ducháček, J.; Stádník, L.; Ptáček, M.; Beran, J. The effect of selected factors on the growth ability of Charolais cattle. Acta Univ. Agric. Silvic. Mendel. Brun. 2014, 62, 255–260. [Google Scholar] [CrossRef]
- Owens, F.N.; Gill, D.R.; Secrist, D.S.; Coleman, S.W. Review of some aspects of growth and development of feedlot cattle. J. Anim. Sci. 1995, 73, 3152–3172. [Google Scholar] [CrossRef]
- Schreurs, N.M.; Garcia, F.; Jurie, C.; Agabriel, J.; Micol, D.; Bauchart, D.; Listrat, A.; Picard, B. Meta-analysis of the effect of animal maturity on muscle characteristics in different muscles, breeds, and sexes of cattle. J. Anim. Sci. 2008, 86, 2872–2887. [Google Scholar] [CrossRef]
- Shibata, M.; Hikino, Y.; Imanari, M.; Matsumoto, K.; Yamamoto, N. Influence of rice whole-crop silage diet on growth performance, carcass and meat characteristics and muscle-related gene expression in Japanese Black steers. Anim. Sci. J. 2016, 87, 929–937. [Google Scholar] [CrossRef]
- Shibata, M.; Hikino, Y.; Imanari, M.; Matsumoto, K. Comprehensive evaluation of growth performance and meat characteristics of a fattening system combining grazing with feeding rice whole-crop silage in Japanese Black steers. Anim. Sci. J. 2019, 90, 504–512. [Google Scholar] [CrossRef]
- Ndlovu, T.; Chimonyo, M.; Okoh, A.I.; Muchenje, V.; Dzama, K.; Raats, J.G. Assessing the nutritional status of beef cattle: Current practices and future prospects. Afr. J. Biotechnol. 2007, 6, 2727–2734. [Google Scholar] [CrossRef]
- Shingu, H.; Hayashi, H.; Touno, E.; Oshibe, A.; Kushibiki, S.; Oda, S.; Katoh, K.; Obara, Y. Characteristics of developmental changes in the kinetics of glucose and urea in Japanese Black calves: Comparison with Holstein calves. J. Anim. Sci. 2007, 85, 2910–2915. [Google Scholar] [CrossRef] [PubMed]
- Ishida, S.; Yonezawa, T.; Eirai, S.; Kizaki, K.; Hashizume, K.; Tomioka, M.; Kurose, Y.; Hirano, T.; Watanabe, D. Hormonal differences in peripheral blood and gene profiling in the liver and lymphocytes in Japanese Black cattle with growth retardation. J. Vet. Med. Sci. 2013, 75, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, U.; Okamoto, K.; Miyamoto, A.; Otoi, T.; Yamato, O.; Tshering, C.; Takagi, M. A Japanese Black breeding herd exhibiting low blood urea nitrogen: A metabolic profile study examining the effect on reproductive performance. Anim. Sci. J. 2013, 84, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Kessler, E.C.; Gross, J.J.; Bruckmaier, R.M.; Albrecht, C. Cholesterol metabolism, transport, and hepatic regulation in dairy cows during transition and early lactation. J. Dairy Sci. 2014, 97, 5481–5490. [Google Scholar] [CrossRef]
- Otomaru, K.; Wataya, K.; Uto, T.; Kasai, K. Blood biochemical values in Japanese Black calves in Kagoshima Prefecture, Japan. J. Vet. Med. Sci. 2016, 78, 301–303. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Watanabe, A.; Takenouchi, N.; Suzuki, H.; Matsuzaki, M. Comparison on the growth characteristics between calves of two different japanese beef breeds suckled by Japanese shorthorn dams. Jpn. Agric. Res. Q. 2020, 54, 93–100. [Google Scholar] [CrossRef]
- 53/2013 R. Real Decreto 53/2013, de 1 de Febrero, Por el Que se Establecen las Normas Básicas Aplicables para la Protección de Los Animales Utilizados en Experimentación y Otros Fines Científicos, Incluyendo la Docencia. 2013. Available online: https://www.boe.es/eli/es/rd/2013/02/01/53 (accessed on 14 March 2022).
- NRC. National Research Council. Nutrient Requirements of Beef Cattle, 7th Revised ed.; Update 2000; The National Academies Press: Washington, DC, USA, 2000; pp. 102–112. [CrossRef]
- Chambaz, A.; Dufey, P.A.; Kreuzer, M.; Gresham, J. Sources of variation influencing the use of real-time ultrasound to predict intramuscular fat in live beef cattle. Can. J. Anim. Sci. 2002, 82, 133–139. [Google Scholar] [CrossRef][Green Version]
- Weik, F.; Hickson, R.E.; Morris, S.T.; Garrick, D.J.; Archer, J.A. Genetic parameters for growth, ultrasound and carcass traits in New Zealand beef cattle and their correlations with maternal performance. Animals 2022, 12, 25. [Google Scholar] [CrossRef]
- Szabó, F.S.S. Performance recording of animals: State of the art, 2004. In Proceedings of the 34th Biennial Session of ICAR, Sousse, Tumisia, 28 May–3 June 2005; Guellouz, M., Dimitriadou, A., Mosconi, C., Eds.; EAAP Scientific Series. Wageningen Academic Publishers: Wageningen, The Netherlands, 2005; Volume 113, pp. 141–145. [Google Scholar] [CrossRef]
- Casas, E.; Lunstra, D.D.; Cundiff, L.V.; Ford, J.J. Growth and pubertal development of F1 steers from Hereford, Angus, Norwegian Red, Swedish Red and White, Friesian, and Wagyu sires. J. Anim. Sci. 2007, 85, 2904–2909. [Google Scholar] [CrossRef]
- Motoyama, M.; Sasaki, K.; Watanabe, A. Wagyu and the factors contributing to its beef quality: A Japanese industry overview. Meat Sci. 2016, 120, 10–18. [Google Scholar] [CrossRef]
- Otomaru, K.; Shiga, H.; Kanome, J.; Yanagita, K. Blood biochemical values in Japanese black breeding cows in Kagoshima Prefecture, Japan. J. Vet. Med. Sci. 2015, 77, 1021–1023. [Google Scholar] [CrossRef]
- Watanabe, U.; Takagi, M.; Yamato, O.; Otoi, T.; Okamoto, K. Retrospective surveillance of metabolic parameters affecting reproductive performance of Japanese Black breeding cows. J. Vet. Sci. 2014, 15, 283–288. [Google Scholar] [CrossRef]
- Brethour, J.R. Using serial ultrasound measures to generate models of marbling and backfat thickness changes in feedlot cattle. J. Anim. Sci. 2000, 78, 2055–2061. [Google Scholar] [CrossRef]
- Tait, R.G. Ultrasound use for body composition and carcass quality assessment in cattle and lambs. Vet. Clin. N. Am. Food Anim. Pract. 2016, 32, 207–218. [Google Scholar] [CrossRef]
- Duff, C.J.; Van Der Werf, J.H.J.; Parnell, P.F.; Clark, S.A. Comparison of two live-animal ultrasound systems for genetic evaluation of carcass traits in Angus cattle. Transl. Anim. Sci. 2021, 5, 1–11. [Google Scholar] [CrossRef]
- Peña, F.; Molina, A.; Juárez, M.; Requena, F.; Avilés, C.; Santos, R.; Domenech, V.; Horcada, A. Use of serial ultrasound measures in the study of growth- and breed-related changes of ultrasonic measurements and relationship with carcass measurements in lean cattle breeds. Meat Sci. 2014, 96, 247–255. [Google Scholar] [CrossRef]
- Ribeiro, F.R.B.; Tedeschi, L.O.; Stouffer, J.R.; Carstens, G.E. A novel technique to assess internal body fat of cattle by using real-time ultrasound. J. Anim. Sci. 2008, 86, 763–767. [Google Scholar] [CrossRef]
- McGilchrist, P.; Polkinghorne, R.J.; Ball, A.J.; Thompson, J.M. The meat standards Australia index indicates beef carcass quality. Animal 2019, 13, 1750–1757. [Google Scholar] [CrossRef]
- Burnett, D.D.; Legako, J.F.; Phelps, K.J.; Gonzalez, J.M. Biology, strategies, and fresh meat consequences of manipulating the fatty acid composition of meat. J. Anim. Sci. 2020, 98, 1–12. [Google Scholar] [CrossRef]
- Pethick, D.W.; Hocquette, J.F.; Scollan, N.D.; Dunshea, F.R. Review: Improving the nutritional, sensory and market value of meat products from sheep and cattle. Animal 2021, 15, 100356. [Google Scholar] [CrossRef] [PubMed]
- Fiore, E.; Fabbri, G.; Gallo, L.; Morgante, M.; Muraro, M.; Boso, M.; Gianesella, M. Application of texture analysis of b-mode ultrasound images for the quantification and prediction of intramuscular fat in living beef cattle: A methodological study. Res. Vet. Sci. 2020, 131, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, B.J.; Wolcott, M.L.; McPhee, M.J. Modeling the relationship between scanned rump and 12th-rib fat in young temperate and tropical bovines: Model development and evaluation. J. Anim. Sci. 2010, 88, 1848–1859. [Google Scholar] [CrossRef] [PubMed]
- Tokunaga, T.; Jomane, F.N.; Mandai, S.; Ishida, T.H.H. Estimation of the marbling development pattern in Japanese Black cattle by using serial ultrasound measurement data. Anim. Sci. J. 2021, 92, 135333. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.Z.; Lee, Y.; Son, H.; Hanna, L.H.; Riley, D.G.; Mannen, H.; Sasazaki, S.; Park, S.P.; Kim, J.J. Genetic characteristics of Korean Jeju Black cattle with high density single nucleotide polymorphisms. Anim. Breed Genet. 2021, 34, 789–800. [Google Scholar] [CrossRef]
- Ryu, J.L.C. Genomic heritability of bovine growth using a mixed model. Asian-Australas. J. Anim. Sci. 2014, 27, 1521–1527. [Google Scholar] [CrossRef]
- Davis, M.E.; Simmen, R.C.M. Genetic parameter estimates for serum insulin-Like growth factor I concentration and performance traits in Angus beef cattle. J. Anim. Sci. 1997, 75, 317–324. [Google Scholar] [CrossRef]
- Lee, C.; Van Tassell, C.P.; Pollak, E.J. Estimation of genetic variance and covariance components for weaning weight in Simmental cattle. J. Anim. Sci. 1997, 75, 325–330. [Google Scholar] [CrossRef]
- Rolfe, K.M.; Snelling, W.M.; Nielsen, M.K.; Freetly, H.C.; Ferrell, C.L.; Jenkins, T.G. Genetic and phenotypic parameter estimates for feed intake and other traits in growing beef cattle, and opportunities for selection. J. Anim. Sci. 2011, 89, 3452–3459. [Google Scholar] [CrossRef]
- Polkinghorne, R.J.; Thompson, J.M. Meat standards and grading. A world view. Meat Sci. 2010, 86, 227–235. [Google Scholar] [CrossRef]
- Matsuzaki, M.; Takizawa, S.; Ogawa, M. Plasma insulin, metabolite concentrations, and carcass characteristics of Japanese Black, Japanese Brown, and Holstein steers. J. Anim. Sci. 1997, 75, 3287–3293. [Google Scholar] [CrossRef] [PubMed]
- Trenkle, A.; Topel, D.G. Relationship of some endocrine measurements to growth and carcass composition of cattle. J. Anim. Sci. 1978, 46, 1604–1609. [Google Scholar] [CrossRef]
- Warner, R.D.; Ferguson, D.M.; Cottrell, J.J.; Knee, B.W. Acute stress induced by the preslaughter use of electric prodders causes tougher beef meat. Aust. J. Exp. Agric. 2007, 47, 782–788. [Google Scholar] [CrossRef]
- Aierqing, S.; Nakagawa, A.; Bungo, T. Association between temperament and polymorphisms of CRH and leptin in Japanese Black Cattle. J. Adv. Vet. Anim. Res. 2020, 7, 1–5. [Google Scholar] [CrossRef]
- Kawaguchi, H.; Miyoshi, N.; Miura, N.; Fujiki, M.; Horiuchi, M.; Izumi, Y.; Miyajima, H.; Nagata, R.; Misumi, K.; Takeuchi, T.; et al. Microminipig, a non-rodent experimental animal optimized for life science research: Novel atherosclerosis model induced by high fat and cholesterol diet. J. Pharmacol. Sci. 2011, 115, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Ference, B.A.; Ginsberg, H.N.; Graham, I.; Ray, K.K.; Packard, C.J.; Bruckert, E.; Hegele, R.A.; Krauss, R.M.; Raal, F.J.; Schunkert, H.; et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease. Evidence from genetic, epidemiologic, and clinical studies. A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2017, 38, 2459–2472. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, A.; Guo, J.-R.; Weng, X.-S.; Ahmed, B.; Hayen, M.; Dahl, G.; Bernard, J.; Tao, S. Effect of maternal heat stress during the dry period on growth and metabolism of calves. J. Dairy Sci. 2016, 99, 3896–3907. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | Starter Feed |
|---|---|
| Crude fiber (%) | 7.1 |
| Ash (%) | 4.9 |
| Crude protein (%) | 17.2 |
| Crude fat (%) | 4 |
| Ca (%) | 0.71 |
| P (%) | 0.34 |
| Na (%) | 0.14 |
| Vitamin E (UI/kg) | 26 |
| Vitamin A (KUI/kg) | 16 |
| Vitamin D (KUI/kg) | 2.2 |
| Co (mg/kg) | 0.2 |
| Mn (mg/kg) | 100 |
| Zn (mg/kg) | 90 |
| Se (mg/kg) | 0.46 |
| I (mg/kg) | 1 |
| Cu (mg/kg) | 26 |
| Nutrient | Growth Dry-TMR | Fattening Wet-TMR | Finishing Dry-TMR |
|---|---|---|---|
| Humidity (%) | 10.90 | 67.79 | 11.35 |
| Dry matter (%) | 89.09 | 32.21 | 88.65 |
| Crude fiber (%) | 17.49 | 22.89 | 11.15 |
| Ash (%) | 6.64 | 6.95 | 5.58 |
| NDF (%) | 35.62 | 51.09 | 26.43 |
| Crude protein (%) | 16.34 | 13.52 | 13.19 |
| Crude fat (%) | 3.50 | 3.94 | 6.98 |
| ADF (%) | 22.73 | 30.12 | 14.16 |
| Ca (%) | 0.852 | 0.717 | 0.628 |
| P (%) | 0.286 | 0.286 | 0.292 |
| Na (%) | 0.175 | 0.304 | 0.219 |
| Cl (%) | 0.323 | 0.479 | 0.419 |
| Mg (%) | 0.34 | 0.331 | 0.219 |
| K (%) | 1.045 | 1.14 | 0.778 |
| S (%) | 0.183 | 0.25 | 0.172 |
| Vitamin E (mg/kg) | 32.32 | 14.693 | 41.62 |
| Vitamin A (KUI/kg) | 2.508 | 1.189 | 4.23 |
| Vitamin D (KUI/kg) | 0.702 | 0.333 | 1.18 |
| NFC | 37.898 | 24.50 | 47.83 |
| Starch (%) | 27.83 | 7.51 | 40.46 |
| Fodder (%) | 17.78 | ||
| ME (kcal/kg) | 2785.93 | 3651.41 | 3026.4 |
| TDN (%) | 73.58 | 62.66 | 80.93 |
| Wagyu | Wangus | ACL | ||||
|---|---|---|---|---|---|---|
| Time Point | n | Age (Months) | n | Age (Months) | n | Age (Months) |
| WEAN | 192 | 4.2 ± 0.95 | 56 | 4.5 ± 0.83 | 19 | 4.4 ± 0.92 |
| GR | 178 | 12.8 ± 1.75 | 85 | 14.1 ± 2.52 | 38 | 14.3 ± 1.59 |
| FIN | 60 | 28.6 ± 3.65 | 55 | 27.9 ± 3.72 | 32 | 20.1 ± 2.53 |
| SL | 11 | 34.9 ± 4.46 | 28 | 32.8 ± 6.46 | ||
| Wagyu | Wangus | ACL | |||||
|---|---|---|---|---|---|---|---|
| Interval | Time Point | n | Age (Months) | n | Age (Months) | n | Age (Months) |
| WEAN to GR | WEAN | 139 | 4.6 ± 0.77 | 50 | 4.7 ± 0.75 | 17 | 4.6 ± 0.93 |
| GR | 12.5 ± 1.74 | 12.7 ± 1.49 | 13.7 ± 0.99 | ||||
| GR to FIN | GR | 57 | 13.8 ± 0.84 | 49 | 15.3 ± 2.36 | 31 | 14.3 ± 1.21 |
| FIN | 28.4 ± 3.39 | 27.6 ± 3.42 | 20 ± 2.51 | ||||
| Interval | Wagyu | Wangus | ACL | p-Value |
|---|---|---|---|---|
| WEAN to GR | 0.916 (0.806–1.025) a | 1.046 (0.915–1.196) b | 1.293 (1.107–1.406) c | <0.001 |
| GR to FIN | 0.628 (0.583–0.753) a | 0.64 (0.587–0.732) a | 0.802 (0.704–1.048) b | 0.001 |
| WEAN to FIN | 0.78 (0.73–0.85) a | 0.86 (0.76–0.89) a | 1.12 (0.92–1.20) b | <0.001 |
| WEAN | GR | FIN | SLAUGHTER | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Median (IQR) | p-Value | Median (IQR) | p-Value | Median (IQR) | p-Value | Median (IQR) | p-Value | ||
| WY | 0.3 (0.2–0.4) a | 0.001 | 0.5 (0.4–0.5) | 0.13 | 0.4 (0.4–0.5) a | 0.001 | 0.4 (0.4–0.6) | 0.54 | |
| BHB (mmol/L) | WN | 0.3 (0.2–0.4) a | 0.5 (0.4–0.5) | 0.5 (0.4–0.5) a | 0.4 (0.3–0.5) | ||||
| ACL | 0.5 (0.4–0.5) b | 0.4 (0.4–0.5) | 0.4 (0.3–0.4) b | ||||||
| WY | 0.2 (0.1–0.3) | 0.2 | 0.2 (0.1–0.3) | 0.066 | 0.3 (0.2–0.3) a | 0.019 | 0.2 (0.1–0.3) | 0.32 | |
| NEFA (mmol/L) | WN | 0.2 (0.1–0.3) | 0.2 (0.2–0.4) | 0.20 (0.1–0.3) b | 0.2 (0.2–0.4) | ||||
| ACL | 0.1 (0.1–0.4) | 0.2 (0.1–0.3) | 0.3 (0.2–0.3) a | ||||||
| WY | 110 (101–120) a | 0.002 | 95 (89.0–100) | 0.84 | 84.5 (80.5–92.0) a | 0.023 | 90.5 (83.0–111) | 0.1 | |
| GLU (mg/dL) | WN | 110 (99–116) a | 95 (90.0–100) | 87.0 (78.3–93.0) a | 83.0 (75.7–97.0) | ||||
| ACL | 118 (115–127) b | 94.1 (90.0–99.0) | 93.0 (84.0–98.0) b | ||||||
| WY | 342 (303–389) | 0.1 | 297 (273–318) | 0.14 | 327 (293–362) a | 0.005 | 302 (282–364) | 0.4 | |
| FRU (mg/dL) | WN | 333 (297–368) | 299 (274–327) | 299 (269–324) b | 292 (272–315) | ||||
| ACL | 317 (280–345) | 307 (286–333) | 299 (278–326) b | ||||||
| WY | 24.0 (18.0–33.0) a | 0.001 | 23.0 (16.5–32.0) ab | 0.012 | 19.5 (13.0–30.5) | 0.109 | 27.5 (13.0–41.0) | 0.69 | |
| LAC (mg/dL) | WN | 22.0 (17.0–37.0) a | 21.71 (16.0–29.5) a | 24.0 (17.0–37.0) | 19.0 (12.5–32.3) | ||||
| ACL | 58.0 (24.8–67.0) b | 30.0 (20.0–39.0) b | 21.0 (13.0–33.0) | ||||||
| WY | 139 (88.0–177) a | 0.027 | 109 (95.0–125) a | 0.001 | 139 (117–162) a | 0.003 | 148 (111–181) | 0.98 | |
| TC (mg/dL) | WN | 140 (113–155) a | 122 (106–140) b | 143 (118–175) a | 137 (124–165) | ||||
| ACL | 93.0 (80.5–117) b | 107 (89.0–143) ab | 114 (97.0–138) b | ||||||
| WY | 54.0 (39.0–64.0) ab | 0.033 | 42.0 (36.5–49.0) a | 0.003 | 47.5 (41.5–60.5) a | 0.016 | 60.0 (48.5–65.0) | 0.97 | |
| HDL (mg/dL) | WN | 59.0 (47.0–66.0) a | 53.47 (45.0–62.0) b | 56.0 (45.0–73.2) b | 54.0 (46.5–87.0) | ||||
| ACL | 42.0 (38.5–49.0) b | 43.5 (33.0–58.0) a | 42.0 (38.0–51.0) a | ||||||
| WY | 20.0 (11.0–28.0) | 0.17 | 10.5 (7.0–14.5) a | <0.001 | 11.0 (6.0–17.5) a | 0.001 | 8.5 (6.0–18.0) | 0.04 | |
| LDL (mg/dL) | WN | 22.0 (15.0–27.0) | 14.0 (9.7–18.0) b | 18.0 (12.0–22.5) b | 19.7 (14.2–22.2) | ||||
| ACL | 18.0 (10.0–22.0) | 15.0 (12.0–18.0) b | 19.0 (13.0–25.0) b | ||||||
| WY | 24.0 (17.0–35.0) | 0.14 | 20.7 (16.0–29.0) a | 0.001 | 30.0 (21.8–45.0) ab | 0.023 | 29.0 (23.0–38.0) | 0.23 | |
| TG (mg/dL) | WN | 28.0 (20.0–39.7) | 25.5 (20.0–34.5) b | 31.0 (25.0–45.0) a | 30.0 (23.5–38.5) | ||||
| ACL | 24.0 (17.0–27.0) | 22.0 (15.0–28.0) a | 21.0 (16.0–43.0) b | ||||||
| WY | 20.0 (16.0–24.0) a | 0.037 | 25.0 (21.0–31.0) a | 0.003 | 24.0 (20.5–28.5) a | 0.001 | 23.0 (19.0–28.0) | 0.05 | |
| Urea (mg/dL) | WN | 23.0 (17.5–27.0) b | 29.5 (24.0–35.3) b | 29.0 (25.0–35.0) b | 28.7 (23.0–39.0) | ||||
| ACL | 20.0 (14.0–23.0) ab | 22.0 (20.0–25.3) c | 24.0 (20.0–28.0) a | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez-Mosquera, J.M.; de Mercado, E.; Fernández-Novo, A.; Gardón, J.C.; Pesántez-Pacheco, J.L.; Pérez-Solana, M.L.; Revilla-Ruiz, Á.; Martínez, D.; Villagrá, A.; Sebastián, F.; et al. Comparison of Pure and Crossbred Japanese Black Steers in Growth Performance and Metabolic Features from Birth to Slaughter at a Spanish Fattening Farm. Animals 2022, 12, 1671. https://doi.org/10.3390/ani12131671
Vázquez-Mosquera JM, de Mercado E, Fernández-Novo A, Gardón JC, Pesántez-Pacheco JL, Pérez-Solana ML, Revilla-Ruiz Á, Martínez D, Villagrá A, Sebastián F, et al. Comparison of Pure and Crossbred Japanese Black Steers in Growth Performance and Metabolic Features from Birth to Slaughter at a Spanish Fattening Farm. Animals. 2022; 12(13):1671. https://doi.org/10.3390/ani12131671
Chicago/Turabian StyleVázquez-Mosquera, Juan M., Eduardo de Mercado, Aitor Fernández-Novo, Juan C. Gardón, José L. Pesántez-Pacheco, María Luz Pérez-Solana, Ángel Revilla-Ruiz, Daniel Martínez, Arantxa Villagrá, Francisco Sebastián, and et al. 2022. "Comparison of Pure and Crossbred Japanese Black Steers in Growth Performance and Metabolic Features from Birth to Slaughter at a Spanish Fattening Farm" Animals 12, no. 13: 1671. https://doi.org/10.3390/ani12131671
APA StyleVázquez-Mosquera, J. M., de Mercado, E., Fernández-Novo, A., Gardón, J. C., Pesántez-Pacheco, J. L., Pérez-Solana, M. L., Revilla-Ruiz, Á., Martínez, D., Villagrá, A., Sebastián, F., Pérez-Garnelo, S. S., & Astiz, S. (2022). Comparison of Pure and Crossbred Japanese Black Steers in Growth Performance and Metabolic Features from Birth to Slaughter at a Spanish Fattening Farm. Animals, 12(13), 1671. https://doi.org/10.3390/ani12131671