
Temporal and Spatial Activity Patterns of Sympatric Wild Ungulates in Qinling Mountains, China


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Data Analysis
3. Results
3.1. General Summary
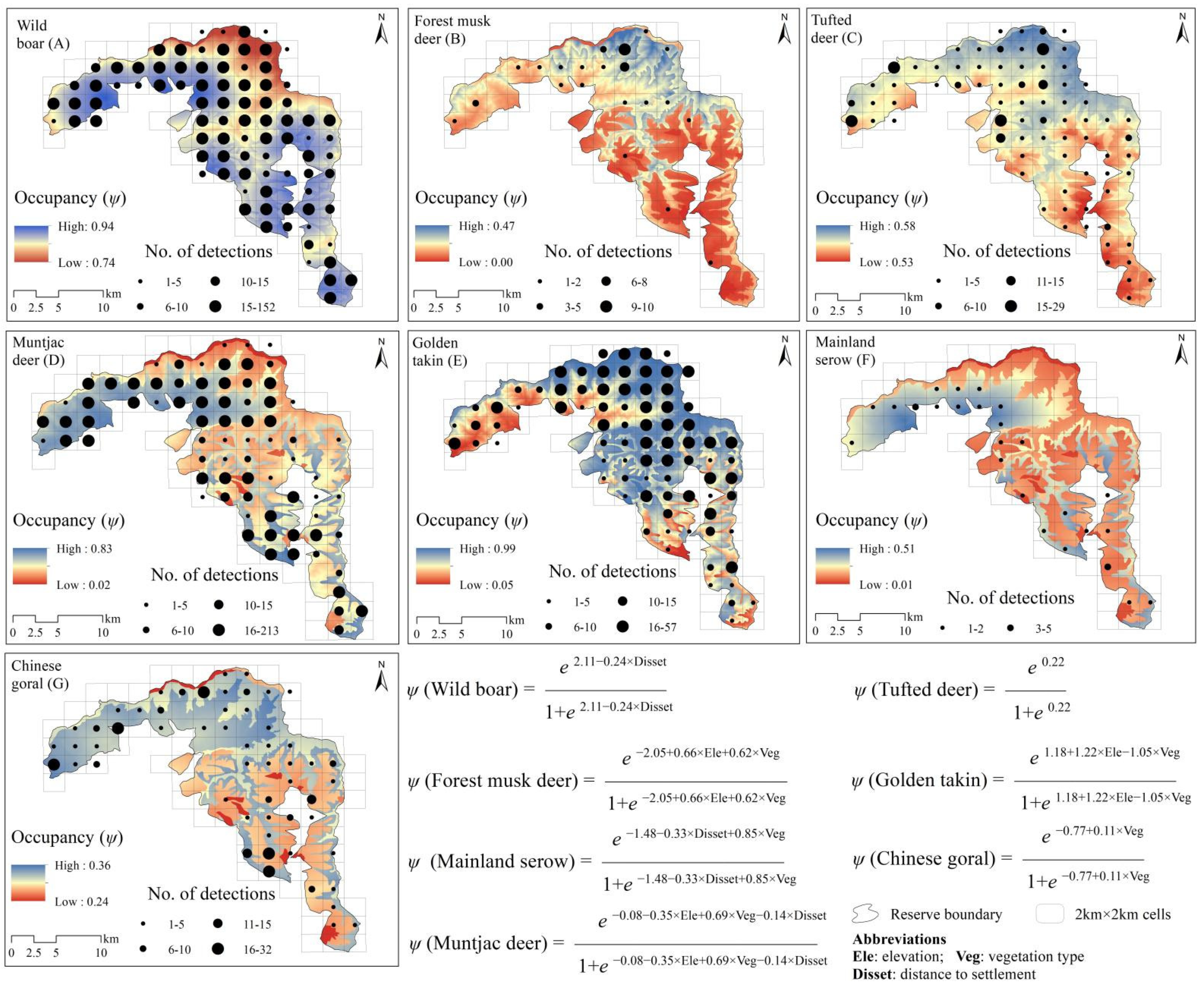
3.2. Spatial Utilization Distribution
3.3. Spatial Overlaps
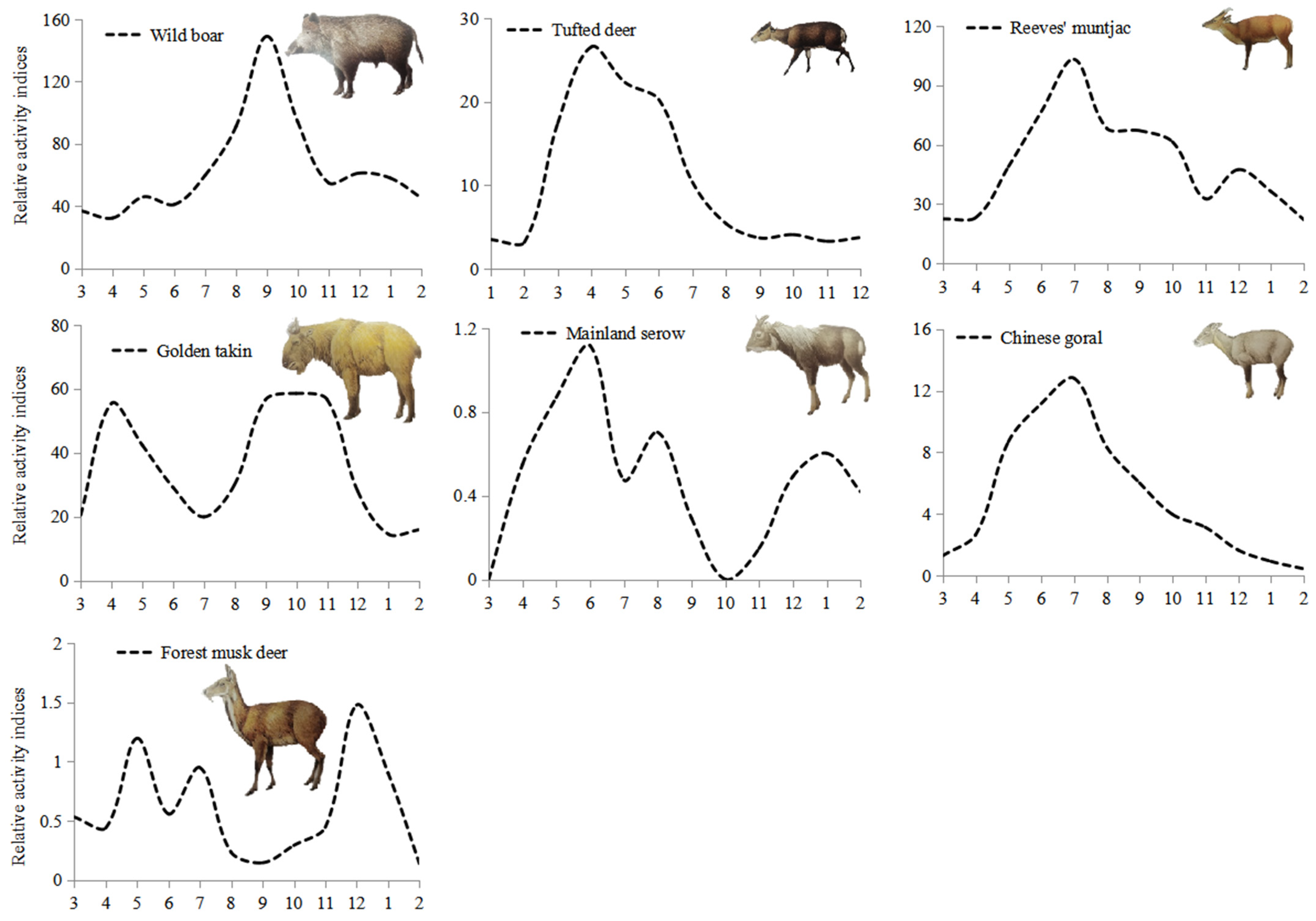
3.4. Annual Activity Patterns
3.5. Daily Temporal Patterns
3.6. Temporal Overlaps
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hillerislambers, J.; Adler, P.B.; Harpole, W.S.; Levine, J.M.; Mayfield, M.M. Rethinking community assembly through the lens of coexistence theory. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 227–248. [Google Scholar] [CrossRef]
- Andersen, G.E.; Johnson, C.N.; Jones, M.E. Space use and temporal partitioning of sympatric Tasmanian devils and spotted-tailed quolls. Austral. Ecol. 2020, 45, 355–365. [Google Scholar] [CrossRef]
- Finke, D.L.; Snyder, W.E. Niche partitioning increases resource exploitation by diverse communities. Science 2008, 321, 1488–1490. [Google Scholar] [CrossRef]
- Kamler, J.F.; Stenkewitz, U.; Klare, U.; Jacobsen, N.F.; Macdonald, D.W. Resource partitioning among cape foxes, bat-eared foxes, and black-backed jackals in South Africa. J. Wildl. Manag. 2012, 76, 1241–1253. [Google Scholar] [CrossRef]
- Letten, A.D.; Ke, P.J.; Fukami, T. Linking modern coexistence theory and contemporary niche theory. Ecol. Monogra 2017, 87, 161–177. [Google Scholar] [CrossRef]
- Carvalho, C.J.; Cardoso, P. Decomposing the causes for niche differentiation between species using hypervolumes. Front. Ecol. Evol. 2020, 8, 243. [Google Scholar] [CrossRef]
- Croose, E.; Bled, F.; Fowler, N.L.; Beyer, D.E., Jr.; Belant, J.L. American marten and fisher do not segregate in space and time during winter in a mixed-forest system. Ecol. Evol. 2019, 9, 4906–4913. [Google Scholar] [CrossRef]
- Farris, Z.J.; Gerber, B.D.; Karpanty, S.; Murphy, A.; Wampole, E.; Ratelolahy, F.; Kelly, M.J. Exploring and interpreting spatiotemporal interactions between native and invasive carnivores across a gradient of rainforest degradation. Biol. Invasions 2020, 22, 2033–2047. [Google Scholar]
- Gordon, I.J.; Prins, H.H.T. The Ecology of Browsing and Grazing II. Ecological Studies (Analysis and Synthesis); Springer: Gewerbestrasse, Switzerland, 2019; p. 239. [Google Scholar]
- Lillian, S.; Redak, R.A.; Daugherty, M.P. Assessing the role of differential herbivore performance among plant species in associational effects involving the invasive stink bug Bagrada hilaris (Hemiptera: Pentatomidae). Community Ecosyst. Ecol. 2019, 48, 114–121. [Google Scholar]
- Rooney, T.P. High white-tailed deer densities benefit graminoids and contribute to biotic homogenization of forest ground-layer vegetation. Plant. Ecol. 2009, 202, 103–111. [Google Scholar] [CrossRef]
- Schulze, E.D.; Bouriaud, O.; Wäldchen, N.; Eisenhauer, H.; Walentowski, C.; Seele, E.; Helnze, U.; Pruschitzke, G.; Dănilă, G.; Marln, D.; et al. Ungulate browsing causes species loss in deciduous forests independent of community dynamics and silvicultural management in central and southeastern europe. Ann. For. Res. 2014, 57, 267–288. [Google Scholar] [CrossRef]
- Kasahara, M.; Fujii, S.; Tanikawa, T.; Mori, A.S. Ungulates decelerate litter decomposition by altering litter quality above and below ground. Eur. J. Forest. Res. 2016, 135, 849–856. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.K.; Liu, W.H. Autumn bed site selection by sika deer (Cervus nippon) in the Taohongling national nature reserve, China. Russ. J. Ecol. 2017, 48, 384–391. [Google Scholar] [CrossRef]
- Li, J.; Xue, Y.D.; Zhang, Y.; Dong, W.; Shan, G.Y.; Sun, R.Q.; Hacker, C.; Wu, B.; Li, D.Q. Spatial and temporal activity patterns of Golden takin (Budorcas taxicolor bedfordi) recorded by camera trapping. PeerJ 2020, 8, e10353. [Google Scholar] [CrossRef]
- Ripple, W.J.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; Nelson, M.P.; et al. Status and ecological effects of the world’s largest carnivores. Science 2014, 343, 1241484. [Google Scholar] [CrossRef]
- Valente, A.M.; Acevedo, P.; Figueiredo, A.M.; Fonseca, C.; Torres, R.T. Overabundant wild ungulate populations in Europe: Management with consideration of socio-ecological consequences. Mammal. Rev. 2020, 50, 353–366. [Google Scholar] [CrossRef]
- Iida, T.; Soga, M.; Koike, S. Large herbivores affect forest ecosystem functions by altering the structure of dung beetle communities. Acta Oecol. 2018, 88, 65–70. [Google Scholar] [CrossRef]
- Pascual-Rico, R.; Pérez-García, J.M.; Sebastián-González, E. Is diversionary feeding a useful tool to avoid human-ungulate conflicts? A case study with the aoudad. Eur. J. Wildl. Res. 2018, 64, 67. [Google Scholar] [CrossRef]
- Khanyari, M.; Uulu, K.Z.; Luecke, S.; Mishra, C.; Suryawanshi, K.R. Understanding population baselines: Status of mountain ungulate populations in the Central Tien Shan Mountains, Kyrgyzstan. Mammalian 2020, 85, 16–23. [Google Scholar] [CrossRef]
- Joshua, Q.I.; Hofmeyr, M.D.; Henen, B.T. Seasonal and site variation in angulate tortoise diet and activity. J. Herpetol. 2010, 44, 124–134. [Google Scholar] [CrossRef]
- Ping, X.G.; Li, C.W.; Jiang, Z.G.; Liu, W.H.; Zhu, H.B. Sexual difference in seasonal patterns of salt lick use by south China sika deer Cervus nippon. Mamm. Biol. 2011, 72, 196–200. [Google Scholar] [CrossRef]
- Guan, T.P.; Ge, B.M.; Mcshea, W.J.; Li, S.; Song, Y.L.; Stewart, C.M. Seasonal migration by a large forest ungulate: A study on takin (Budorcas taxicolor) in Sichuan Province, China. Eur. J. Wildl. Res. 2013, 59, 81–91. [Google Scholar] [CrossRef]
- Kukielka, E.; Barasona, J.A.; Cowie, C.E.; Drewe, J.A.; Gortazar, C.; Cotarelo, I.; Vicente, J. Spatial and temporal interactions between livestock and wildlife in South Central Spain assessed by camera traps. Prev. Vet. Med. 2013, 3, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Guan, T.P.; Gong, M.H.; Hu, J.; Qiu, J.; Chen, L.M.; Cheng, W.L.; Zheng, W.C.; Song, Y.L. The rhythm of Chinese Munjac (Muntiacus reevesi) visiting artificial mineral licks during autumn. Chin. J. Zool. 2015, 50, 169–175. [Google Scholar]
- Viana, D.S.; Granados, J.E.; Fandos, P.; Pérez, J.M.; Cano-Manuel, F.J.; Burón, D.; Fandos, G.; Aguado, M.Á.P.; Figuerola, J.; Soriguer, R. Linking seasonal home range size with habitat selection and movement in a mountains ungulate. Mov. Ecol. 2018, 6, 1. [Google Scholar] [CrossRef]
- Semenzato, P.; Cagnacci, F.; Ossi, F.; Ecel, E.; Morellet, N.; Hewison, A.J.M.; Sturaro, E.; Ramanzin, M. Behavioural heat-stress compensation in a cold-adapted ungulate: Forage-mediated responses to warming Alpine summers. Ecol. Lett. 2021, 24, 1556–1568. [Google Scholar] [CrossRef]
- Shakeri, Y.N.; White, K.S.; Waite, J.N. Staying close to home: Ecological constraints on space use and range fidelity in a mountain ungulate. Ecol. Evol. 2021, 11, 11051–11064. [Google Scholar] [CrossRef]
- Karanth, K.K. Wildlife in the matrix: Spatio-temporal patterns of herbivore occurrence in Karnataka, India. Environ. Manag. 2016, 57, 189–206. [Google Scholar] [CrossRef]
- Nathan, R.; Spiegel, O.; Fortmann-Roe, S.; Harel, R.; Wikelski, M.; Getz, W.M. Using triaxial acceleration data to identify behavioral modes of free-ranging animals: General concepts and tools illustrated for griffon vultures. J. Exp. Biol. 2012, 215, 986–996. [Google Scholar] [CrossRef]
- Frey, S.; Fisher, J.T.; Burton, A.C.; Volpe, J.P. Investigating animal activity patterns and temporal niche partitioning using camera-trap data: Challenges and opportunities. Remote Sens. Ecol. Con. 2017, 3, 123–132. [Google Scholar] [CrossRef]
- Burton, A.C.; Neilson, E.; Moreira, D.; Ladle, A.; Steenweg, R.; Fisher, J.T.; Bayne, E.; Boutin, S. Review: Wildlife camera trapping: A review and recommendations for linking surveys to ecological processes. Methods. Ecol. Evol. 2015, 52, 675–685. [Google Scholar] [CrossRef]
- Rowcliffe, J.M.; Kays, R.; Kranstauber, B.; Carbone, C.; Jansen, P.A. Quantifying levels of animal activity using camera trap data. Methods Ecol. Evol. 2014, 5, 1170–1179. [Google Scholar] [CrossRef]
- Massé, A.; Côté, S. Spatiotemporal variations in resources affect activity and movement patterns of white-tailed deer (Odocoileus virginianus) at high density. Can. J. Zool. 2013, 91, 252–263. [Google Scholar] [CrossRef]
- de Satgé, J.; Teichman, K.; Cristescu, B. Competition and coexistence in a small carnivore guild. Oecologia 2017, 184, 873–884. [Google Scholar] [CrossRef]
- Houngbégnon, F.G.A.; Cornelis, D.; Vermeulen, C.; Sonké, B.; Ntie, S.; Fayoll, A.; Fonteyn, D.; Lhoest, S.; Evrard, Q.; Yapi, F.; et al. Daily activity patterns and co-occurrence of duikers revealed by an intensive camera trap survey across central African Rainforests. Animals 2020, 10, 2200. [Google Scholar] [CrossRef]
- Mori, E.; Lovari, S.; Cozzi, F.; Gabbrielli, C.; Giari, C.; Torniai, L.; Romeo, G.; Ferretti, F.; Fattorini, N. Safety or satiety? Spatiotemporal behaviour of a threatened herbivore. Mmm. Biol. 2020, 100, 49–61. [Google Scholar] [CrossRef]
- Tian, C.; Zhang, Y.Y.; Liu, Z.X.; Dayananda, B.; Fu, X.B.; Yuan, D.; Tua, Z.B.; Luo, C.P.; Li, J.Q. Temporal niche patterns of large mammals in Wanglang nature reserve, China. Globa. Ecol. Conserv. 2020, 22, e01015. [Google Scholar] [CrossRef]
- Viviano, A.; Mori, E.; Fattorini, N.; Mazza, G.; Lazzeri, L.; Panichi, A.; Strianese, L.; Mohamed, W.F. Spatiotemporal Overlap between the European Brown Hare and Its Potential Predators and Competitors. Animals 2021, 11, 562. [Google Scholar] [CrossRef]
- Li, J.; Li, D.Q.; Hacker, C.; Dong, W.; Wu, B.; Xue, Y.D. Spatial co-occurrence and temporal activity patterns of sympatric mesocarnivores guild in Qinling Mountains. Glob. Ecol. Conserv. 2022, 36, e02129. [Google Scholar] [CrossRef]
- Sheng, M.C. Biodiversity Research and Conservation on Qinling Mountains in China; Science Press: Beijing, China, 2010. [Google Scholar]
- He, B.S.; Sun, R.Q.; Cheng, P.; Dong, W.; Wang, J.; Wang, D.J.; Li, S. Baseline survey of mammal and bird diversity using camera-trapping in the Changqing National Nature Reserve of Shaanxi Province. Acta. Theriol. Sinica 2016, 36, 348–356. [Google Scholar]
- Jia, X.D.; Liu, X.H.; Yang, X.Z.; Wu, P.F.; Melissa, S.; Cai, Q.; He, X.B.; Zhu, Y. Seasonal activity patterns of ungulates in Qinling Mountains based on camera-trap data. Diodivers. Sci. 2014, 22, 737–745. [Google Scholar]
- Li, S.; Zhang, X.F.; Chen, P.; Wang, J.; Xiang, D.Q.; Dong, W.; Zhang, X.M.; He, B.S.; Sun, R.Q.; Zhao, N.X.; et al. The community structure and elevational patterns of forest ungulates at the southern slope of the Qinling Mountains, China. Chin. J. Zool. 2014, 49, 633–643. [Google Scholar]
- Ren, Y.; Yang, X.Z.; Wang, X.J. Wildlife Resource in Changqing National Nature Reserve; Northwestern University Press: Xi’an, China, 2002. [Google Scholar]
- Zhao, N.X.; Zhang, X.M.; Dong, W. Field Guide to Nature History and Wildlife of Qinling Mountains; Encyclopedia of China Publishing House: Beijing, China, 2014. [Google Scholar]
- Li, J.; Wang, X.L.; Yang, M.Y.; Cheng, D.X.; Wang, X.J.; Luo, P.; Liu, F.; Xue, Y.D.; Li, G.L.; Zhang, Y.G.; et al. Construction progress of camera-trapping database from the Nature Reserves Biological Specimen Resources Sharing Sub-platform. Biodivers. Sci. 2020, 28, 1081–1089. [Google Scholar] [CrossRef]
- Li, S.; Mcshea, W.J.; Wang, D.J.; Shao, L.K.; Shi, X.G. The used of infrared-triggered cameras for surveying phasianids in Sichuan Province, China. IBIS 2010, 152, 299–309. [Google Scholar] [CrossRef]
- Davis, C.; Rich, L.N.; Farris, Z.J.; Kelly, M.J.; di Bitetti, M.S.; Blanco, Y.D.; Albanesi, S.; Farhadinia, M.S.; Gholikhani, N.; Hamel, S.; et al. Ecological correlates of the spatial co-occurrence of sympatric mammalian carnivores worldwide. Ecol. Lett. 2018, 21, 1401–1412. [Google Scholar] [CrossRef]
- Zeng, Z.G.; Skidmore, A.K.; Song, Y.L.; Wang, T.J.; Gong, H.S. Seasonal altitudinal movements of golden takin in the Qinling Mountains of China. J. Wildl. Manag. 2008, 72, 611–617. [Google Scholar] [CrossRef]
- Lesmeister, D.B.; Nielsen, C.K.; Schauber, E.M.; Hellgren, E.C. Spatial and temporal structure of a mesocarnivore guild in midwestern north America. Wildl. Monogr. 2015, 191, 1–61. [Google Scholar] [CrossRef]
- Sørensen, T. A method of establishing groups of equal amplitude in plant sociology based on similarity of species content and its application to analyses of the vegetation on Danish commons. Kong. Dan. Vidensk. Selsk. Biol. Skr. 1948, 5, 1–34. [Google Scholar]
- Torretta, E.; Serafini, M.; Puopolo, F.; Schenone, L. Spatial and temporal adjustments allowing the coexistence among carnivores in Liguria (N-W Italy). Acta Ethol 2015, 19, 123–132. [Google Scholar] [CrossRef]
- Blake, J.G.; Mosquera, D.; Guerra, J.; Loiselle, B.A.; Romo, D.; Swing, K. Mineral licks as diversity hotspots in lowland forest of eastern Ecuador. Diversity 2011, 3, 217–234. [Google Scholar] [CrossRef]
- Mella-Méndez, I.; Flores-Peredo, R.; Pérez-Torres, J.; Hernández-González, S.; González-Urbie, D.U.; Bolívar-Cimé, B.S. Activity patterns and temporal niche partitioning of dogs and medium-sized wild mammals in urban parks of Xalapa, Mexico. Urban. Ecosyst. 2019, 22, 1061–1070. [Google Scholar] [CrossRef]
- Manly, B.F.J.; McDonald, L.L.; Thomas, D.L.; McDonald, T.L.; Erickson, W.P. Resource Selection by Animals: Statistical Design and Analysis for Field Studies, 2nd ed.; Springer: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Bu, H.L.; Wang, F.; McShea, W.J.; Lu, Z.; Wang, D.J.; Li, S. Spatial co-occurrence and activity patterns of mesocarnivores in the temperate forests of Southwest China. PLoS ONE 2016, 11, e0164271. [Google Scholar] [CrossRef]
- Linkie, M.; Ridout, M.S. Assessing tiger–prey interactions in Sumatran rainforests. J. Zool. 2011, 3, 224–229. [Google Scholar] [CrossRef]
- Riduot, M.S.; Linkie, M. Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Environ. Stat. 2009, 14, 322–337. [Google Scholar] [CrossRef]
- Meredith, M.; Ridout, M. Overview of the “Overlap” Package. 2021. Available online: http://cran.cs.wwu.edu/web/packages/overlap/vignettes/overlap.pdf (accessed on 10 November 2021).
- Finnegan, S.P.; Gantchoff, M.G.; Hill, J.; Silveira, L.; Tôrres, N.M.; Jácomo, A.T.; Uzal, A. “When the felid’s away, the msocarnivores play”: Seasonal temporal segregation in a neotropical carnivore guild. Mamm. Biol. 2021, 101, 631–638. [Google Scholar] [CrossRef]
- Monterroso, P.; Alves, P.C.; Ferreras, P. Plasticity in circadian activity patterns of mesocarnivores in Southwestern Europe: Implications for species coexistence. Behav. Ecol. Sociobiol. 2014, 68, 1403–1417. [Google Scholar] [CrossRef]
- Agha, M.; Batter, T.; Bolas, E.C.; Collins, A.C.; Da Rocha, D.G.; Monteza-Moreno, C.M.; Preckler-Quisquater, S.; Sollmann, R. A review of wildlife camera trapping trends across Africa. Afr. J. Ecol. 2018, 56, 694–701. [Google Scholar] [CrossRef]
- Li, P.; Zhang, Z.J.; Yang, H.; Wei, W.; Zhou, H.; Hong, M.S.; Fu, M.X.; Song, X.Q.; Yu, J. Study on the activity rhythms of ungulates in Daxiangling Nature Reserve based on infrared camera trapping. J. Sichuan Forest. Sci. Technol. 2021, 42, 18–23. [Google Scholar]
- Sun, J.X.; Li, J.Q.; Wang, Y.Q.; Li, S.; Guan, T.P.; Wang, J.; Xia, W.C.; Xu, H.G. Study on the activity rhythms of nine ungulates in Summer and Autumn in Sichuan. J. Ecol. Rural Environ. 2018, 34, 1003–1009. [Google Scholar]
- Liu, M.X.; Zhu, B.Q.; Wang, Y.J.; Zengren, Z.M.; Fang, C.; Jin, G.X.; Leng, Z.C.; Guan, T.P.; Sun, Y.Y. Activity rhythm and seasonal changes of Elaphodus cephalophus in Baihe National Nature Reserve, Sichuan Province. J. Sichuan Forest. Sci. Technol. 2021, 42, 27–32. [Google Scholar]
- Peng, K.; Chen, X.; Weng, P.; Wei, Y.; Yang, Z.S.; Dai, Q. The activity rhythm survey of ungulates in Baishuihe national nature reserve based on infrared camera trapping. J. Sichuan Forest. Sci. Technol. 2021, 42, 76–82. [Google Scholar]
- Yang, C.; Wang, Y.Q.; Huang, X.F.; Yuan, X.D.; Zhou, H.Y.; Fang, H.C.; Li, D.Y.; Li, J.Q. Activity rhythm of Muntiacus reevesi based on infrared camera technology. J. Guangxi Normal Univ. 2021, 39, 65–70. [Google Scholar]
- Marinho, P.H.; Fonseca, C.R.; Sarmento, P.; Fonseca, C.; Venticinque, E.M. Temporal niche overlap among mesocarnivores in a Caatinga dry forest. Eur. J. Wildl. Res. 2020, 66, 34. [Google Scholar] [CrossRef]
- Zeng, Z.G.; Song, Y.L. Daily activity rhythm and time budget of golden takin in spring and summer. Acta Theriol. Sinica 2001, 21, 7–13. [Google Scholar]
- Sanusi, M.A.M.; Shukor, M.A.; Juliana, W.A.W.; Traeholt, C. Activity pattern of selected ungulates at Krau Wildlife Reserve. AIP Conf. Proc. 2013, 1571, 325–330. [Google Scholar]
- Liu, X.H.; Wu, P.F.; Shao, X.M.; Songer, M.; Cai, Q.; He, X.B.; Zhu, Y. Diversity and activity patterns of sympatric animals among four types of forest habitat in Guanyinshan Nature Reserve in the Qinling Mountains, China. Environ. Sci. Pollut. Res. Vol. 2017, 24, 16465–16477. [Google Scholar] [CrossRef]
- Espinosa, S.; Salvador, J. Hunters’ landscape accessibility and daily activity of ungulates in Yasuní Biosphere Reserve, Ecuador. Therya 2017, 8, 45–52. [Google Scholar] [CrossRef][Green Version]
- Ou, S.H.; Sheng, H.L.; Lu, H.J. Diet of the black muntjac and tufted deer. J. Shanghai Normal Univ. 1981, 1, 111–115. [Google Scholar]
- Sheng, H.L.; Lu, H.J. Distribution, habitats and resource status of the tufted deer (Elaphodus cepholophus). Acta Zool. Sinica 1982, 26, 307–311. [Google Scholar]
- Sheng, H.L.; Wang, P.C. Ecology and utilization of Muntjac reeves. Chin. J. Zool. 1976, 1, 39–40. [Google Scholar]
- Liu, H.; Li, L.; Ma, Q.; Wang, D.M.; Zhang, S.Q.; Zhu, Y.P. Review on wild boar research. Sichuan J. Zool. 2011, 30, 310–313. [Google Scholar]
- Li, Y.; Viña, A.; Yang, W.; Chen, X.; Zhang, J.; Ouyang, Z.Y.; Liang, Z.; Liu, J.G. Effects of conservation policies on forest cover change in giant panda habitat regions, China. Land Use Policy 2013, 33, 42–53. [Google Scholar] [CrossRef]
- Zhang, P.C.; Shao, G.F.; Zhao, G.; Master, D.C.L.; Parker, G.R.; Dunning, J.B.; Li, Q.L. China’s forest policy for the 21st century. Science 2007, 288, 2135–2136. [Google Scholar] [CrossRef]
- Morishima, K.; Nakano, T.; Aizawa, M. Sika deer presence affects of the host-parasite interface of a Japanese land leech. Ecol. Evol. 2020, 10, 6030–6038. [Google Scholar] [CrossRef]
- Zhu, S.N.; Song, H.D.; Wang, K.; Zong, C.; He, S.W. Cause of golden takin attacking people in the villages. J. Econ. Anim. 2015, 19, 140–146. [Google Scholar]
- Wang, L.; Bu, S.H.; Song, H.D.; Li, G.C.; Zheng, X.L. Research on the villagers’ tolerance to wild boars in the Qinling Mountainous region of Xi’an, Shannxi. Sichuan J. Zool. 2020, 39, 563–571. [Google Scholar]
- Steinmetz, R.; Seuaturien, N.; Intanajitjuy, P.; Inrueang, P.; Prempree, K. The effects of prey depletion on dietary niches of sympatric apex predators in Southeast Asia. Integr. Zool. 2020, 16, 19–32. [Google Scholar] [CrossRef]
- Xu, W.; Pimm, S.L.; Du, A.; Su, Y.; Ouyang, Z.Y. Transforming protected area management in China. Trends Ecol. Evol. 2019, 34, 762–766. [Google Scholar] [CrossRef]
- Li, S.; McShea, W.J.; Wang, D.J.; Gu, X.D.; Zhang, X.F.; Zhang, L.; Shen, X.L. Retreat of large carnivores across the giant panda distribution range. Nat. Ecol. Evol. 2020, 40, 1327–1331. [Google Scholar] [CrossRef]
- Tan, W.S.; Hamzah, N.B.A.; Saaban, S.; Zawakhir, N.A.; Rao, Y.; Jamaluddin, N.; Cheong, F.; Khalid, N.B.; Mohdsaat, N.I.; Zaidee Ee, E.N.B.; et al. Observations of occurrence and daily activity patterns of ungulates in the Endau Rompin Landscape, peninsular Malaysia. J. Threat. Taxa 2018, 10, 11245–11253. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | No. of Sites | No. of 2 km × 2 km Cells | No. of Independent Detections (n) | Capture Rate per 1000 Camera Trap Days (Mean ± SD) |
|---|---|---|---|---|
| Wild boar (Sus scrofa) | 495 | 87 | 5827 | (7.05 ± 8.52) a |
| Forest musk deer (Moschus berezovskii) | 33 | 21 | 57 | (1.12 ± 1.38) b |
| Tufted deer (Elaphodus cephalophus) | 266 | 76 | 1029 | (2.74 ± 3.60) b |
| Reeves’ muntjac (Muntiacus reevesi) | 275 | 75 | 4799 | (10.01 ± 14.17) c |
| Golden takin (Budorcas taxicolor bedfordi) | 382 | 81 | 3323 | (5.24 ± 6.47) a |
| Mainland serow (Capricornis milneedwardsii) | 33 | 46 | 46 | (1.00 ± 1.07) b |
| Chinese goral (Naemorhedus griseus) | 151 | 55 | 503 | (2.29 ± 2.76) b |
| Species | Models | Number of Parameters | AIC | ΔAIC | AICWt | ψ | Pr |
|---|---|---|---|---|---|---|---|
| Wild boar | psi (Disset); p (Season + Veg) | 5 | 3919.84 | 0.00 | 0.14 | 0.889 | 0.599 |
| psi (Disset + Veg); p (Season + Veg) | 6 | 3919.85 | 0.10 | 0.14 | 0.887 | 0.600 | |
| psi (Disset + Disriv); p (Season + Veg) | 6 | 3920.21 | 0.37 | 0.12 | 0.888 | 0.599 | |
| psi (Ele + Veg); p (Season + Veg) | 6 | 3921.76 | 1.92 | 0.05 | 0.887 | 0.600 | |
| psi (Ele); p (Season + Veg) | 5 | 3921.79 | 1.95 | 0.05 | 0.889 | 0.599 | |
| psi (Ele + Disset); p (Season + Veg) | 6 | 3921.83 | 1.99 | 0.05 | 0.889 | 0.600 | |
| Model average | 0.888 | 0.599 | |||||
| Forest musk deer | psi (Ele + Veg); p (Veg) | 5 | 437.24 | 0.00 | 0.20 | 0.141 | 0.114 |
| psi (Ele + Veg); p (Season) | 5 | 437.29 | 0.04 | 0.20 | 0.140 | 0.107 | |
| psi (Ele); p (Veg) | 4 | 437.76 | 0.52 | 0.16 | 0.190 | 0.075 | |
| psi (Ele + Disriv); p (Veg) | 5 | 439.03 | 1.79 | 0.08 | 0.186 | 0.077 | |
| psi (Ele + Veg); p (Season + Veg) | 6 | 439.24 | 1.99 | 0.07 | 0.141 | 0.115 | |
| Model average | 0.160 | 0.098 | |||||
| Tufted deer | psi (.); p (Season + Veg) | 4 | 2302.17 | 0.00 | 0.23 | 0.554 | 0.277 |
| psi (Ele); p (Season + Veg) | 5 | 2303.45 | 1.27 | 0.12 | 0.553 | 0.277 | |
| psi(Veg);p (Season + Veg) | 5 | 2303.62 | 1.45 | 0.11 | 0.552 | 0.278 | |
| psi (Disriv); p (Season + Veg) | 5 | 2303.65 | 1.48 | 0.11 | 0.553 | 0.277 | |
| psi (Disset); p (Season + Veg) | 5 | 2303.04 | 1.86 | 0.09 | 0.554 | 0.277 | |
| Model average | 0.553 | 0.277 | |||||
| Muntjac deer | psi (Ele + Veg); p (Veg) | 5 | 2703.02 | 0.00 | 0.41 | 0.486 | 0.584 |
| psi (Disset + Veg); p (Veg) | 5 | 2704.15 | 1.13 | 0.24 | 0.485 | 0.584 | |
| psi (Ele + Veg); p (Season + Veg) | 6 | 2705.01 | 1.99 | 0.15 | 0.486 | 0.584 | |
| Model average | 0.486 | 0.584 | |||||
| Golden takin | psi (Ele + Veg); p (Veg) | 5 | 3487.72 | 0.00 | 0.17 | 0.713 | 0.440 |
| psi (Ele); p (Veg) | 4 | 3488.39 | 0.67 | 0.12 | 0.712 | 0.441 | |
| psi (Ele + Veg); p (Season + Veg) | 6 | 3488.76 | 1.03 | 0.10 | 0.714 | 0.439 | |
| psi (Ele); p (.) | 3 | 3489 | 1.28 | 0.09 | 0.710 | 0.442 | |
| psi (Ele + Veg); p (.) | 4 | 3489.27 | 1.55 | 0.08 | 0.712 | 0.442 | |
| psi (Ele); p (Season + Veg) | 5 | 3489.36 | 1.64 | 0.07 | 0.711 | 0.440 | |
| Model average | 0.712 | 0.441 | |||||
| Mainland serow | psi (Disset + Veg); p (.) | 4 | 431.6 | 0.00 | 0.13 | 0.201 | 0.068 |
| psi (Disset + Veg); p (Veg) | 5 | 431.82 | 0.22 | 0.12 | 0.217 | 0.090 | |
| psi (Disset + Veg); p (Season + Veg) | 6 | 432.06 | 0.46 | 0.11 | 0.226 | 0.092 | |
| psi (Disset + Veg); p (Season) | 5 | 432.34 | 0.74 | 0.09 | 0.208 | 0.066 | |
| psi (Ele + Veg); p (Season + Veg) | 6 | 433.02 | 1.42 | 0.07 | 0.268 | 0.089 | |
| psi (Ele + Veg); p (.) | 4 | 433.43 | 1.83 | 0.05 | 0.208 | 0.066 | |
| psi (Veg); p (Veg) | 4 | 433.47 | 1.87 | 0.05 | 0.254 | 0.086 | |
| Model average | 0.226 | 0.080 | |||||
| Chinese goral | psi (Veg); p (Season + Veg) | 5 | 1500.53 | 0.00 | 0.19 | 0.322 | 0.254 |
| psi (.); p (Season + Veg) | 4 | 1501.3 | 0.77 | 0.13 | 0.311 | 0.262 | |
| psi (Veg); p (Season) | 4 | 1501.72 | 1.19 | 0.10 | 0.322 | 0.254 | |
| psi (Disset + Veg); p (Season + Veg) | 6 | 1502.3 | 1.77 | 0.08 | 0.311 | 0.262 | |
| psi (Ele + Veg); p (Season + Veg) | 6 | 1502.52 | 1.99 | 0.07 | 0.322 | 0.254 | |
| psi (Disriv + Veg); p (Season + Veg) | 6 | 1502.53 | 1.99 | 0.07 | 0.323 | 0.254 | |
| Model average | 0.318 | 0.257 |
| Species | Model Component | Covariates | Estimate (β) | SE | Z | p |
|---|---|---|---|---|---|---|
| Wild boar | Occupancy | Intercept | 2.11 | 0.15 | 13.86 | <0.001 *** |
| Disset | −0.24 | 0.17 | 1.36 | 0.174 | ||
| Veg | 0.07 | 0.13 | 0.55 | 0.581 | ||
| Disriv | −0.04 | 0.10 | 0.39 | 0.698 | ||
| Ele | −0.04 | 0.13 | 0.33 | 0.740 | ||
| Detection | Intercept | 0.41 | 0.04 | 9.97 | <0.001 *** | |
| Veg | 0.28 | 0.04 | 6.70 | <0.001 *** | ||
| Season | −0.08 | 0.04 | 2.02 | 0.0436* | ||
| Forest musk deer | Occupancy | Intercept | −2.05 | 0.50 | 4.12 | <0.001 *** |
| Ele | 0.66 | 0.23 | 2.89 | 0.003 ** | ||
| Veg | 0.62 | 0.55 | 1.12 | 0.263 | ||
| Disriv | −0.02 | 0.10 | 0.22 | 0.823 | ||
| Detection | Intercept | −2.30 | 0.49 | 4.72 | <0.001 *** | |
| Veg | 0.21 | 0.53 | 0.40 | 0.689 | ||
| Season | 0.00 | 0.12 | 0.03 | 0.973 | ||
| Tufted deer | Occupancy | Intercept | 0.22 | 0.13 | 1.70 | 0.090 |
| Ele | 0.02 | 0.06 | 0.29 | 0.775 | ||
| Veg | 0.01 | 0.06 | 0.25 | 0.808 | ||
| Disriv | −0.01 | 0.05 | 0.25 | 0.807 | ||
| Disset | 0.01 | 0.04 | 0.13 | 0.898 | ||
| Detection | Intercept | −1.09 | 0.08 | 12.12 | <0.001 *** | |
| Season | −0.76 | 0.08 | 9.42 | <0.001 *** | ||
| Veg | 0.19 | 0.07 | 2.93 | 0.003 ** | ||
| Muntjac deer | Occupancy | Intercept | −0.08 | 0.09 | 0.85 | 0.40 |
| Ele | −0.35 | 0.24 | 1.45 | 0.15 | ||
| Veg | 0.69 | 0.11 | 6.56 | <0.001 *** | ||
| Disset | −0.14 | 0.22 | 0.63 | 0.53 | ||
| Detection | Intercept | 0.34 | 0.06 | 5.95 | <0.001 *** | |
| Veg | 0.15 | 0.06 | 2.48 | 0.013 * | ||
| Season | 0.00 | 0.02 | 0.04 | 0.969 | ||
| Golden takin | Occupancy | Intercept | 1.18 | 0.14 | 8.60 | <0.001 *** |
| Ele | 1.22 | 0.17 | 7.18 | <0.001 *** | ||
| Veg | −1.05 | 0.13 | 0.78 | 0.436 | ||
| Detection | Intercept | −0.24 | 0.05 | 5.04 | <0.001 *** | |
| Veg | 0.06 | 0.05 | 1.10 | 0.272 | ||
| Season | 0.01 | 0.03 | 0.40 | 0.692 | ||
| Mainland serow | Occupancy | Intercept | −1.48 | 0.45 | 3.25 | 0.001 ** |
| Disset | −0.33 | 0.29 | 1.15 | 0.252 | ||
| Veg | 0.85 | 0.51 | 1.66 | 0.096 | ||
| Ele | −0.07 | 0.18 | 0.39 | 0.690 | ||
| Detection | Intercept | −2.56 | 0.39 | 6.64 | <0.001 *** | |
| Veg | −0.34 | 0.42 | 0.81 | 0.420 | ||
| Season | −0.10 | 0.17 | 0.59 | 0.558 | ||
| Chinese goral | Occupancy | Intercept | −0.77 | 0.13 | 5.84 | <0.001 *** |
| Veg | 0.11 | 0.14 | 0.76 | 0.450 | ||
| Disset | 0.04 | 0.10 | 0.42 | 0.677 | ||
| Ele | −0.01 | 0.09 | 0.07 | 0.946 | ||
| Disriv | 0.00 | 0.03 | 0.03 | 0.980 | ||
| Detection | Intercept | −1.10 | 0.13 | 8.75 | <0.001 *** | |
| Season | −0.34 | 0.11 | 3.07 | 0.021 * | ||
| Veg | 0.26 | 0.12 | 2.13 | 0.035 * |
 Wild boar Wild boar | Δ = 0.54 (0.43–0.63) | Δ = 0.83 (0.79–0.84) | Δ = 0.79 (0.77–0.80) | Δ = 0.84 (0.81–0.84) | Δ = 0.84 (0.72–0.91) | Δ = 0.86 (0.82–0.89) |
| Sij = 0.34 |  Forest musk deer Forest musk deer | Δ = 0.69 (0.57–0.78) | Δ = 0.66 (0.54–0.76) | Δ = 0.64 (0.52–0.73) | Δ = 0.62 (0.46–0.76) | Δ = 0.59 (0.46–0.69) |
| Sij = 0.93 | Sij = 0.43 |  Tufted deer Tufted deer | Δ = 0.90 (0.87–0.92) | Δ = 0.88 (0.84–0.90) | Δ = 0.82 (0.80–0.94) | Δ = 0.86 (0.82–0.90) |
| Sij = 0.93 | Sij = 0.42 | Sij = 0.90 |  Reeves muntjac Reeves muntjac | Δ = 0.89 (0.87–0.91) | Δ = 0.84 (0.81–0.94) | Δ = 0.90 (0.87–0.93) |
| Sij = 0.96 | Sij = 0.41 | Sij = 0.91 | Sij = 0.88 |  Golden takin Golden takin | Δ = 0.83 (0.81–0.93) | Δ = 0.88 (0.85–0.92) |
| Sij = 0.44 | Sij = 0.26 | Sij = 0.46 | Sij = 0.46 | Sij = 0.45 |  Mainland serow Mainland serow | Δ = 0.84 (0.83–0.95) |
| Sij = 0.77 | Sij = 0.39 | Sij = 0.76 | Sij = 0.74 | Sij = 0.81 | Sij = 0.40 |  Chinese goral Chinese goral |
| Species | n(wi) in Time Period | Kruskal-Wallis Tests (χ2, df = 2) | ||
|---|---|---|---|---|
| Diurnal | Nocturnal | Crepuscular | ||
| Wild boar (Sus scrofa) | 4368(1.71) | 668(0.29) | 791(0.81) | 360.99, p < 0.001 |
| Forest musk deer (Moschus berezovskii) | 29(1.16) | 12(0.53) | 16(1.68) | 3.44, p = 0.179 |
| Tufted deer (Elaphodus cephalophus) | 661(1.47) | 138(0.34) | 230(1.34) | 4.56, p = 0.102 |
| Reeves’ muntjac (Muntiacus reevesi) | 2747(1.31) | 911(0.48) | 1141(1.43) | 509.63, p < 0.001 |
| Golden takin (Budorcas taxicolor bedfordi) | 1992(1.37) | 624(0.47) | 707(1.28) | 198.64, p < 0.001 |
| Mainland serow (Capricornis milneedwardsii) | 27(1.34) | 8(0.44) | 11(1.43) | 4.98, p = 0.083 |
| Chinese goral (Naemorhedus griseus) | 315(1.43) | 83(0.42) | 105(1.25) | 72.69, p < 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Xue, Y.; Liao, M.; Dong, W.; Wu, B.; Li, D. Temporal and Spatial Activity Patterns of Sympatric Wild Ungulates in Qinling Mountains, China. Animals 2022, 12, 1666. https://doi.org/10.3390/ani12131666
Li J, Xue Y, Liao M, Dong W, Wu B, Li D. Temporal and Spatial Activity Patterns of Sympatric Wild Ungulates in Qinling Mountains, China. Animals. 2022; 12(13):1666. https://doi.org/10.3390/ani12131666
Chicago/Turabian StyleLi, Jia, Yadong Xue, Mingfu Liao, Wei Dong, Bo Wu, and Diqiang Li. 2022. "Temporal and Spatial Activity Patterns of Sympatric Wild Ungulates in Qinling Mountains, China" Animals 12, no. 13: 1666. https://doi.org/10.3390/ani12131666
APA StyleLi, J., Xue, Y., Liao, M., Dong, W., Wu, B., & Li, D. (2022). Temporal and Spatial Activity Patterns of Sympatric Wild Ungulates in Qinling Mountains, China. Animals, 12(13), 1666. https://doi.org/10.3390/ani12131666