
Casting the Net Widely for Change in Animal Welfare: The Plight of Birds in Zoos, Ex Situ Conservation, and Conservation Fieldwork

Abstract
Simple Summary
Abstract
1. Introduction
2. Birds in Captive Environments: Identifying or Avoiding Behavioral Problems
2.1. General Behavioral Problems and Their Causes
2.2. Pain and Emotions
“The experiments involved captures of wild flocking songbirds, confining them to an artificial laboratory setting singly or in pairs. To induce (further) stress, the experimenter yelled at them, rattled the cages, and rolled the cages swiftly back and forth to prevent perching. In one experiment, birds held captive were so distressed they lost 11% of their body weight within 5 days. In a wounding experiment, although the experimenter used anesthesia before inflicting the injury, no pain relief was provided, which meant the birds woke up in pain from the wounds. In oil feeding experiments, crude oil was fed to one group of birds while a second/control group was not. When the two groups of birds were compared, both had been under so much stress and pain that they experienced the same rate of weight loss. Moreover, the experiments used sparrows. There is little correlation between sparrows and aquatic birds, the species generally affected by oil spills, and studies of penguins and ducks have produced widely varying results. The birds in those experiments underwent prolonged captivity and repeated painful injections and stressful anesthetic episodes before they were killed”.[76]
2.3. Enrichment
2.3.1. Physical Environment
2.3.2. Multispecies Housing
2.3.3. Problematic Medical Interventions
2.3.4. Veterinary Response to Interventionist Practices
- (1)
- The procedure(s) is deemed necessary for the safety, health, husbandry, and well-being of the bird(s) and cannot be accomplished by other nonsurgical means such as an avicultural husbandry management practice;
- (2)
- The procedure(s) is humanely performed in a valid research setting where such a procedure(s) has been approved by an institutional animal care and use committee or an appropriate oversight organization that considers the procedure(s) necessary for the study; and
- (3)
- The procedure(s) is deemed necessary by the administering veterinarian and is not on the list of condoned procedures in this statement [158].
3. In Situ and Ex Situ Conservation
3.1. Some General Methods for Field Data
“Sound science requires animal subjects to be physically, physiologically, and behaviorally unharmed. Accordingly, publication of methods that contravenes animal welfare principles risks perpetuating inhumane approaches and bad science”.[171]
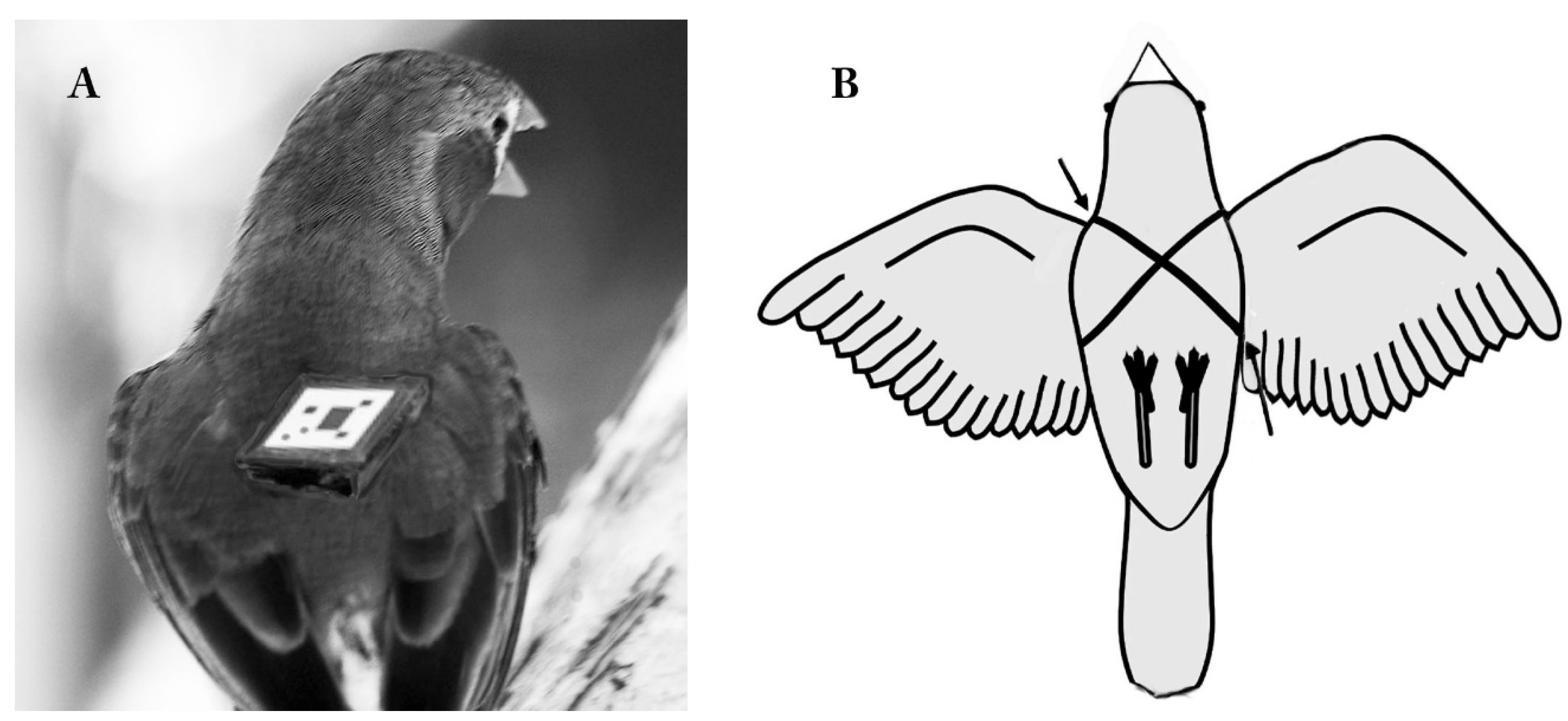
3.2. Conservation and the Demand for Data: The Technological ‘Solution’ (Telemetry)
“Recent technological advances facilitated the continuous improvement of avian-tracking devices allowing the study of individual movement patterns in ever-increasing detail. The emergence of cutting-edge tracking devices caused great leaps in the study of movement ecology in the past couple of decades, thereby increasing our knowledge about the global space-use of wide-ranging birds”.[231]
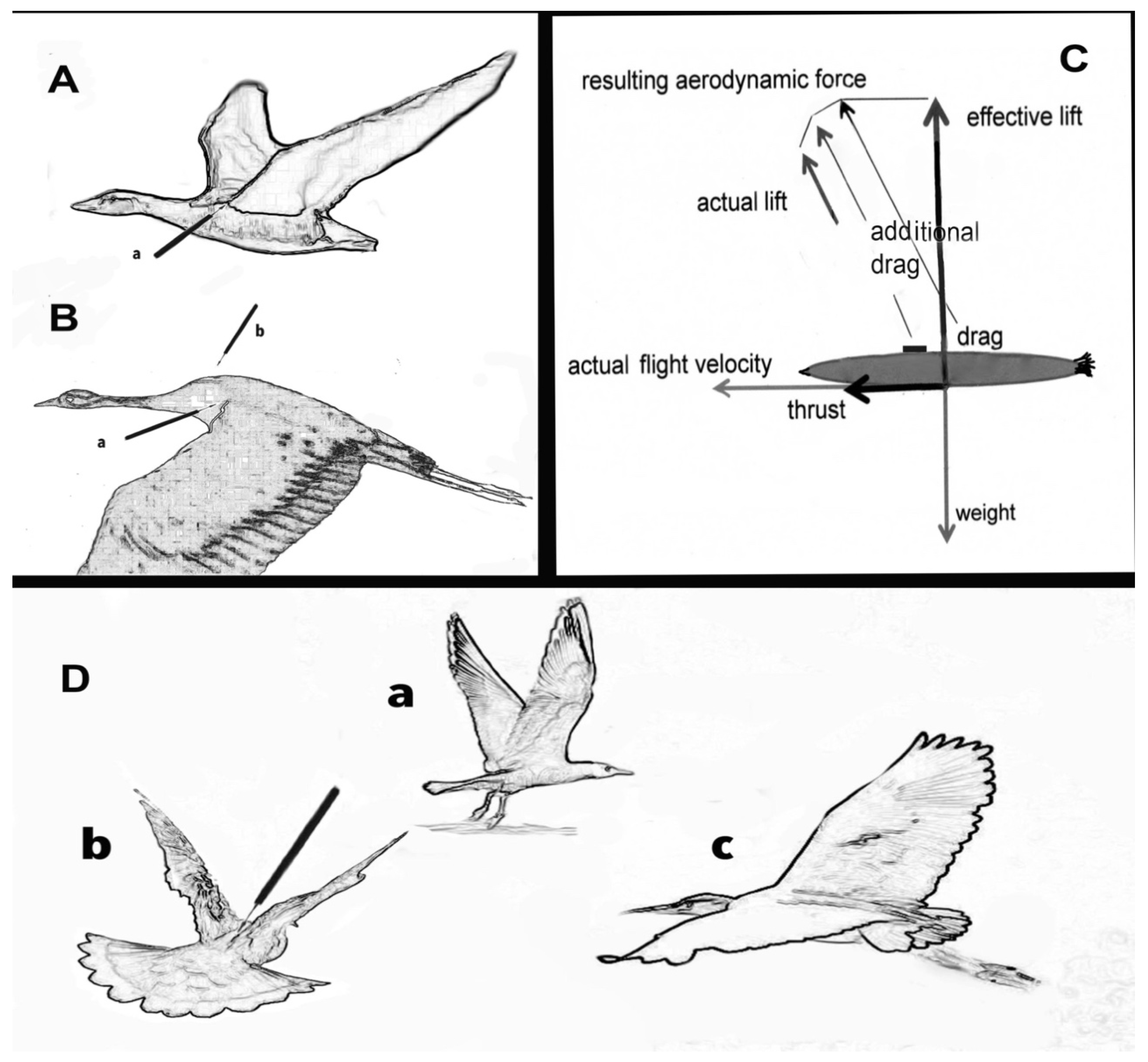
3.3. Some Harmful Effects of Telemetry
4. Solutions
4.1. Telemetry and Ex Situ Conservation
- (1)
- The manufacture of telemetry items for research: I am aware that the manufacturers of these devices are not to blame although their products may vary from relatively innocuous to dangerous. The problem lies fairly and squarely with the institutions giving permission to individual researchers to use specific tools for their research and some blame must also go to the researchers themselves who could express concerns and could report these to manufacturers directly, as surprisingly few may have done. Equally, journals can play their part by not just accepting an ethics approval statement from the institution of the researcher but by making their own separate ethics statements that preclude field practices known to be harmful.
- (2)
- We need more research of the effects of these devices on the animals’ behavior. For example, some avian species, resident or migratory, may be affected in different ways. I am not aware that there has been detailed research conducted on the effects of signal transmission although we know of birds detecting electromagnetic fields [260].
- (3)
- The AAV (American Avian Veterinarians), as was shown above, with its position statement has had a powerful effect on some zoo practices. It could easily add another position statement, such as: that it does not support any procedures that attach a foreign object to a bird on a temporary or prolonged basis unless it can be shown that such attachment has few if any deleterious effects that would impact on overall health, cause prolonged pain or permanently and irrevocably alter avian normal function or even result in death. (author’s wording).
4.2. Ex Situ Conservation and the Problem of the Technology Employed
- (1)
- limiting the amount of time such devices (no matter how improved in weight) should stay on a bird;
- (2)
- ensuring to never use devices on females in any studies, and most decisively not around breeding time;
- (3)
- abolishing practices/devices that lower reproductive rates, have been shown to lead to serious physical health issues, long term suffering or death in a known percentage of birds or in specific species.
- (4)
- explore the possibility of smaller and more aerodynamically shaped devices, more in the shape of pencils rather than in the shape of small match boxes.
- (5)
- ensuring that no device is on any bird beyond a set period of time (the shorter the better) and that there are safe and predictable ways in which such devices will self-release/self-destruct; at the moment it is not always clear how and when such devices, once fitted, will actually be removed (and by what means).
- (6)
- with the decline of migratory birds, in particular, there is a heightened need to learn where their flight paths are so that such flight paths can be protected. Under such circumstance it would be twice as important to go back to manufacturers and seek innovation of new and significantly smaller and lighter tracking devices that are tested extensively and meet new standards, i.e., are proven not to cause friction in flight or damage wings or other parts of the body or leave them so weak that they cannot complete their, often arduous, migration flights [222].
4.3. The Zoo Environment and Ex Situ Conservation
5. Conclusions
- (1)
- to support more rigorous science and
- (2)
- to further improve bird welfare [263].
Funding
Data Availability Statement
Conflicts of Interest
References
- Rosenberg, K.V.; Dokter, A.M.; Blancher, P.J.; Sauer, J.R.; Smith, A.C.; Smith, P.A.; Stanton, J.C.; Panjabi, A.; Helft, L.; Parr, M.; et al. Decline of the North American avifauna. Science 2019, 366, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Prowse, T.A.A.; O’Connor, P.J.; Collard, S.J.; Peters, K.J.; Possingham, H.P. Optimising monitoring for trend detection after 16 years of woodland-bird surveys. J. Appl. Ecol. 2021, 58, 1090–1100. [Google Scholar] [CrossRef]
- Yom-Tov, Y. Global warming and body mass decline in Israeli passerine birds. Proc. R. Soc. B Biol. Sci. 2001, 268, 947–952. [Google Scholar] [CrossRef]
- Chan, D.T.C.; Poon, E.S.K.; Wong, A.T.C.; Sin, S.Y.W. Global trade in parrots–Influential factors of trade and implications for conservation. Global Ecol. Conserv. 2021, 30, e01784. [Google Scholar] [CrossRef]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Bedford, E. Number of Pet Birds in the U.S. 2000–2017. Statistics and Market Data on Consumer Goods & FMCG—Pets and Animal Supplies (Statistics and Facts on Pets & Animal Supplies). 2019. Available online: http://www.Statistica.com (accessed on 18 June 2021).
- Born Free Foundation. International Animal Welfare and Conservation Charity. Available online: http://www.bornfree.org.uk (accessed on 18 June 2021).
- Clarke, R.V.; Rolf, A. Poaching, habitat loss and the decline of neotropical parrots: A comparative spatial analysis. J. Exp. Criminol. 2013, 9, 333–353. [Google Scholar] [CrossRef]
- Wright, T.F.; Toft, C.A.; Enkerlin-Hoeflich, E.; Gonzalez-Elizondo, J.; Albornoz, M.; Rodríguez-Ferraro, A.; Rojas-Suárez, F.; Sanz, V.; Trujillo, A.; Beissinger, S.R.; et al. Nest Poaching in Neotropical Parrots. Conserv. Biol. 2001, 15, 710–720. [Google Scholar] [CrossRef]
- Shemesh, I.I.; Rozen-Zvi, B.; Kalechman, Y.; Gafter, U.; Sredni, B. Prevents Diabetic Nephropathy Progression and Mesangial Cell Dysfunction: Regulation of the AKT Downstream Pathway. PLoS ONE 2014, 9, e114287. [Google Scholar] [CrossRef]
- Brown, M.B.; Brown, C.R. Blood Sampling Reduces Annual Survival in Cliff Swallows (Petrochelidon pyrrhonota). Auk 2009, 126, 853–861. [Google Scholar] [CrossRef]
- Arnold, J.M.; Oswald, S.A.; Voigt, C.C.; Palme, R.; Braasch, A.; Bauch, C.; Becker, P.H. Taking the stress out of blood collection: Comparison of field blood-sampling techniques for analysis of baseline corticosterone. J. Avian Biol. 2008, 39, 588–592. [Google Scholar] [CrossRef]
- Fair, J.E.P.; Jones, J. (Eds.) Guidelines to the Use of Wild Birds in Research; Ornithological Council: Washington, DC, USA, 2010. [Google Scholar]
- Buck, V. Who will start the 3Rs ball rolling for animal welfare? Nature 2007, 446, 856. [Google Scholar] [CrossRef]
- Würbel, H. Publications should include an animal-welfare section. Nature 2007, 446, 257. [Google Scholar] [CrossRef] [PubMed]
- Osborne, N.J.; Payne, D.; Newman, M.L. Journal Editorial Policies, Animal Welfare, and the 3Rs. Am. J. Bioeth. 2009, 9, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Leus, K.; Bingaman Lackey, L.; van Lint, W.; de Man, D.; Riewald, S.; Veldkam, A.; Wijmans, J. Sustainability of European Association of Zoos and Aquaria Bird and Mammal Populations. WAZA Mag. 2011, 12, 11–14. [Google Scholar]
- Kalmar, I.D.; Janssens, G.P.; Moons, C.P. Guidelines and Ethical Considerations for Housing and Management of Psittacine Birds Used in Research. ILAR J. 2010, 51, 409–423. [Google Scholar] [CrossRef] [PubMed]
- Lindsjö, J.; Fahlman, Å.; Törnqvist, E. Animal Welfare from Mouse to Moose—Implementing The Principles of The 3rs in Wildlife Research. J. Wildl. Dis. 2016, 52, S65–S77. [Google Scholar] [CrossRef]
- Soulsbury, C.; Gray, H.; Smith, L.; Braithwaite, V.; Cotter, S.; Elwood, R.W.; Wilkinson, A.; Collins, L.M. The welfare and ethics of research involving wild animals: A primer. Methods Ecol. Evol. 2020, 11, 1164–1181. [Google Scholar] [CrossRef]
- Huss, R.J. Valuing Man’s and Woman’s Best Friend: The Moral and Legal Status of Companion Animals. Marquette Law Rev. 2002, 86, 47. [Google Scholar]
- Seibert, L. Mental Health Issues in Captive Birds. In Mental Health and Well-being in Animals, 2nd ed.; McMillan, F.D., Ed.; CABI Publishers: Wallingford, UK, 2020; pp. 291–301. [Google Scholar]
- Dickens, M.J.; Bentley, G.E. Stress, captivity, and reproduction in a wild bird species. Horm. Behav. 2014, 66, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Bateson, M.; Matheson, S.M. Performance on a categorisation task suggests that removal of environmental enrichment induces ‘pessimism’ in captive European starlings (Sturnus vulgaris) Anim. Welf. South Mimms Engl. 2007, 16, S33–S36. [Google Scholar]
- Apfelbeck, B.; Raess, M. Behavioural and hormonal effects of social isolation and neophobia in a gregarious bird species, the European starling (Sturnus vulgaris). Horm. Behav. 2008, 54, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Aydinonat, D.; Penn, D.J.; Smith, S.; Moodley, Y.; Hoelzl, F.; Knauer, F.; Schwarzenberger, F. Social Isolation Shortens Telomeres in African Grey Parrots (Psittacus erithacus erithacus). PLoS ONE 2014, 9, e93839. [Google Scholar] [CrossRef]
- Mellor, D.J. Updating animal welfare thinking: Moving beyond the “Five Freedoms” towards “a Life Worth Living”. Animals 2016, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Lindenmayer, D.; Pope, M.; Cunningham, R.; Donnelly, C.; Nix, H. Roosting of the Sulphur-Crested Cockatoo Cacatua galerita. Emu-Austral Ornithol. 1996, 96, 209–212. [Google Scholar] [CrossRef]
- Grindlinger, H.M.; Ramsey, E. Compulsive feather picking in birds. Arch. Gen. Psychiat. 1991, 48, 857. [Google Scholar] [CrossRef]
- Meehan, C.; Garner, J.; Mench, J. Isosexual pair housing improves the welfare of young Amazon parrots. Appl. Anim. Behav. Sci. 2003, 81, 73–88. [Google Scholar] [CrossRef]
- Marino, A. Tips and Tricks for Foraging and Enrichment. J. Avian Med. Surg. 2018, 32, 152–156. [Google Scholar] [CrossRef]
- Gaunt, A.S.; Hikida, R.S.; Jehl, J.R. Rapid Atrophy and Hypertrophy of an Avian Flight Muscle. Auk 1990, 107, 649–659. [Google Scholar] [CrossRef]
- Mellor, E.; Brilot, B.; Collins, S. Abnormal repetitive behaviours in captive birds: A Tinbergian review. Appl. Anim. Behav. Sci. 2018, 198, 109–120. [Google Scholar] [CrossRef]
- Lesku, J.A.; Vyssotski, A.L.; Martinez-Gonzalez, D.; Wilzeck, C.; Rattenborg, N.C. Local sleep homeostasis in the avian brain: Convergence of sleep function in mammals and birds? Proc. R. Soc. B. 2011, 278, 2419–2428. [Google Scholar] [CrossRef]
- Cooper, C.E.; Withers, P.; Hurley, L.; Griffith, S. The Field Metabolic Rate, Water Turnover, and Feeding and Drinking Behavior of a Small Avian Desert Granivore During a Summer Heatwave. Front. Physiol. 2019, 10, 1405. [Google Scholar] [CrossRef] [PubMed]
- Cornelius, J.M.; Perreau, G.; Bishop, V.R.; Krause, J.S.; Smith, R.; Hahn, T.P.; Meddle, S.L. Social information changes stress hormone receptor expression in the songbird brain. Horm. Behav. 2017, 97, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Hodinka, B.; Ashley, N.T. Effect of sleep loss on executive function and plasma corticosterone levels in an arctic-breeding songbird, the Lapland longspur (Calcarius lapponicus). Horm. Behav. 2020, 122, 104764. [Google Scholar] [CrossRef]
- McMillan, F.D. The Concept of Quality of Life in Animals. In Mental Health and Well-Being in Animals; McMillan, F.D., Ed.; CABI Publishers: Wallingford, UK, 2005; pp. 181–200, 291–301. [Google Scholar]
- Fisher, H. The Nutrition of Birds. In Avian Biology; Elsevier: Amsterdam, The Netherlands, 1972; pp. 431–469. [Google Scholar] [CrossRef]
- Baldwin, A.L.; Schwartz, G.E.; Hopp, D.H. Are investigators aware of environmental noise in animal facilities and that this noise may affect experimental data? J. Am. Assoc. Lab. Anim. Sci. 2007, 46, 45–51. [Google Scholar] [PubMed]
- Aengus, W.L.; Millam, J.R. Taming parent-reared orange-winged Amazon parrots by neonatal handling. Zoo Biol. 1999, 18, 177–187. [Google Scholar] [CrossRef]
- Feenders, G.; Bateson, M. Hand rearing affects emotional responses but not basic cognitive performance in European starlings. Anim. Behav. 2013, 86, 127–138. [Google Scholar] [CrossRef][Green Version]
- Mason, G.; Rushen, J. Stereotypic Animal Behaviour: Fundamentals and Applications to Welfare; CABI: Wallingford, UK, 2008. [Google Scholar]
- Greenwell, P.J.; Montrose, V.T. The gray matter: Prevention and reduction of abnormal behavior in companion gray parrots (Psittacus erithacus). J. Vet. Behav. 2017, 20, 44–51. [Google Scholar] [CrossRef]
- South, J.M.; Pruett-Jones, S. Patterns of flock size, diet, and vigilance of naturalized Monk parakeets in Hyde Park, Chicago. Condor 2000, 102, 848–854. [Google Scholar] [CrossRef]
- Doane, B.M.; Qualkinbush, T. My Parrot, My Friend: An Owner’s Guide to Parrot Behavior; Howell Book House @ Turner Publishing: Nashville, TN, USA, 1994. [Google Scholar]
- Duque, J.F.; Rasmussen, T.; Rodriguez, A.; Stevens, J.R. The role of mesotocin on social bonding in pinyon jays. Ethology 2020, 126, 165–175. [Google Scholar] [CrossRef]
- Zeigler-Hill, V.; Highfill, L. The Interpersonal Circumplex: A Complementary Approach for Understanding Animal Personality. In Personality in Nonhuman Animals; Vonk, J., Weiss, A., Kuczaj, S.A., Eds.; Springer: Berlin, Germany, 2017; pp. 39–53. [Google Scholar]
- Richter, S.H.; Hintze, S. From the individual to the population–and back again? Emphasising the role of the individual in animal welfare science. App. Anim. Behav. Sci. 2019, 212, 1–8. [Google Scholar] [CrossRef]
- Clark, F. Cognitive enrichment and welfare: Current approaches and future directions. Anim. Behav. Cogn. 2017, 4, 52–71. [Google Scholar] [CrossRef]
- Hopper, L.M. Cognitive research in zoos. Curr. Opin. Behav. Sci. 2017, 16, 100–110. [Google Scholar] [CrossRef]
- Rogers, L.J.; Kaplan, G. Does Functional Lateralization in Birds Have any Implications for Their Welfare? Symmetry 2019, 11, 1043. [Google Scholar] [CrossRef]
- Spinka, M.; Wemelsfelder, F. Environmental challenge and animal agency. In Animal Welfare, 3rd ed.; Appleby, M.C., Olsson, I.A.S., Galindo, F., Eds.; CABI International: Wallingford, UK, 2018; pp. 39–55. [Google Scholar] [CrossRef]
- Pepperberg, I.M. Acquisition of the same/different concept by an African Grey parrot (Psittacus erithacus): Learning with respect to categories of color, shape, and material. Learn. Behav. 1987, 15, 423–432. [Google Scholar] [CrossRef]
- Pepperberg, I.M. Numerical competence in an African Gray parrot (Psittacus erithacus). J. Comp. Psych. 1994, 108, 36–44. [Google Scholar] [CrossRef]
- Evans, G.W. Environmental stress and health. In Handbook of Health Psychology; Revenson, T.A., Gurung, R.A.R., Eds.; Angus & Robertson: Sydney, NSW, Australia, 2001; pp. 365–385. [Google Scholar]
- Svendsen, L. Boredom Is in Your Mind; Springer: Berlin/Heidelberg, Germany, 2019; pp. 135–147. [Google Scholar] [CrossRef]
- Auersperg, A.M.I.; Köck, C.; O’Hara, M.; Huber, L. Tool making cockatoos adjust the lengths but not the widths of their tools to function. PLoS ONE 2018, 13, e0205429. [Google Scholar] [CrossRef]
- Rössler, T.; Mioduszewska, B.; O’Hara, M.; Huber, L.; Prawiradilaga, D.M.; Auersperg, A.M.I. Using an Innovation Arena to compare wild-caught and laboratory Goffin’s cockatoos. Sci. Rep. 2020, 10, 8681. [Google Scholar] [CrossRef] [PubMed]
- Laschober, M.; Mundry, R.; Huber, L.; Schwing, R. Kea (Nestor notabilis) show flexibility and individuality in within-session reversal learning tasks. Anim. Cogn. 2021, 24, 1339–1351. [Google Scholar] [CrossRef]
- Mettke-Hofmann, C.; Winkler, H.; Leisler, B. The Significance of Ecological Factors for Exploration and Neophobia in Parrots. Ethology 2002, 108, 249–272. [Google Scholar] [CrossRef]
- Papini, M.R.; Penagos-Corzo, J.C.; Pérez-Acosta, A.M. Avian Emotions: Comparative Perspectives on Fear and Frustration. Front. Psychol. 2019, 9, 2707. [Google Scholar] [CrossRef]
- Story Carter, J. Nine in 10 Zoos ‘Failing’ Welfare Standards; ABC Radio National (Blueprint with Jonathan Green, Online abc.net.au): Sydney, NSW, Australia, 2016. [Google Scholar]
- Ebisawa, K.; Nakayama, S.; Pai, C.; Kinoshita, R.; Koie, H. Prevalence and risk factors for feather damaging behavior in psittacine birds: Analysis of a Japanese nationwide survey. PLoS ONE 2021, 16, e0254610. [Google Scholar] [CrossRef]
- Ward, S.J.; Sherwen, S.; Clark, F.E. Advances in Applied Zoo Animal Welfare Science. J. Appl. Anim. Welf. Sci. 2018, 21, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Farm Animal Welfare Council (FAWC). Five Freedoms. UK Natl Arch. 1979. Available online: http://webarchive.nationalarchives.gov.uk/20121010012427/ (accessed on 18 June 2021).
- McCulloch, S.P. A Critique of FAWC’s Five Freedoms as a Framework for the Analysis of Animal Welfare. J. Agric. Environ. Ethic. 2012, 26, 959–975. [Google Scholar] [CrossRef]
- Botreau, R.; Veissier, I.; Butterworth, A.; Bracke, M.B.; Keeling, L.J. Definition of criteria for overall assessment of animal welfare. Anim. Welf. Potters Bar 2007, 16, 225. [Google Scholar]
- Webster, J. Animal welfare: Freedoms, dominions and “a life worth living”. Animals 2016, 6, 35. [Google Scholar] [CrossRef] [PubMed]
- Macmillen, R.E.; Hinds, D.S. Water Economy of Granivorous Birds: California House Finches. Condor 1998, 100, 493–503. [Google Scholar] [CrossRef]
- Nikolov, S.; Kanakov, D. Influencing factors leading to damaging behavior—Feather pecking and cannibalism in game birds. Trakia J. Sci. 2020, 18, 377–387. [Google Scholar] [CrossRef]
- Malik, A.; Valentine, A. Pain in birds: A review for veterinary nurses. Vet. Nurs. J. 2017, 33, 11–25. [Google Scholar] [CrossRef]
- Dawkins, M.S. Behavioural deprivation: A central problem in animal welfare. Appl. Anim. Behav. Sci. 1988, 20, 209–225. [Google Scholar] [CrossRef]
- Tiokhin, L. Do symptoms of illness serve signaling functions? Quart. Rev. Biol. 2016, 91, 177–195. [Google Scholar] [CrossRef]
- Goldberg, M.E. How to be a pain management advocate for exotic and zoo animals. Vet. Nurse 2017, 8, 389–397. [Google Scholar] [CrossRef]
- Grimm, D. PETA targets early-career wildlife researcher. Science 2017, 357, 1087. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G. Bird Minds. Cognition and Behaviour of Australian Native Birds; CSIRO Publishing: Melbourne, VIC, Australia, 2015. [Google Scholar]
- Langford, J.M. Avian bedlam: Toward a biosemiosis of troubled parrots. Environ. Hum. 2017, 9, 84–107. [Google Scholar] [CrossRef]
- Bertin, A.; Beraud, A.; Lansade, L.; Blache, M.-C.; Diot, A.; Mulot, B.; Arnould, C. Facial display and blushing: Means of visual communication in blue- and-yellow macaws (Ara ararauna)? PLoS ONE 2018, 13, e0201762. [Google Scholar] [CrossRef]
- Carpenter, J.A. Neurobiology and Communication in Parrots with an Emphasis on Eye Signals. Master’s Thesis, Vienna University, Vienna, Austria, 2019. [Google Scholar]
- Both, C.; Dingemanse, N.J.; Drent, P.J.; Tinbergen, J.M. Pairs of extreme avian personalities have highest reproductive success. J. Anim. Ecol. 2005, 74, 667–674. [Google Scholar] [CrossRef]
- Nicolaus, M.; Tinbergen, J.M.; Bouwman, K.M.; Michler, S.P.M.; Ubels, R.; Both, C.; Kempenaers, B.; Dingemanse, N. Experimental evidence for adaptive personalities in a wild passerine bird. Proc. R. Soc. B Biol. Sci. 2012, 279, 4885–4892. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, W.; Serrano-Davies, E.; Quinn, J.L. Do personality and innovativeness influence competitive ability? An experimental test in the great tit. Behav. Ecol. 2017, 28, 1435–1444. [Google Scholar] [CrossRef]
- Silva, P.A.; Trigo, S.; Marques, C.I.; Cardoso, G.; Soares, M.C. Experimental evidence for a role of dopamine on avian personality traits. J. Exp. Biol. 2020, 223, jeb216499. [Google Scholar] [CrossRef]
- Luescher, A.U. (Ed.) Manual of Parrot Behavior; Blackwell Publishing: Oxford, UK, 2006. [Google Scholar]
- Van Zeeland, Y.R.; Spruit, B.M.; Rodenburg, T.B.; Riedstra, B.; van Hierden, Y.M.; Buitenhuis, B.; Korte, S.M.; Lumeij, J.T. Feather damaging behaviour in parrots: A review with consideration of comparative aspects. Appl. Anim. Behav. Sci. 2009, 121, 75–95. [Google Scholar] [CrossRef]
- Weary, D.M.; Fraser, D. Signalling need: Costly signals and animal welfare assessment. Appl. Anim. Behav. Sci. 1995, 44, 159–169. [Google Scholar] [CrossRef]
- Sol, D.; Duncan, R.P.; Blackburn, T.M.; Cassey, P.; Lefebvre, L. Big brains, enhanced cognition, and response of birds to novel environments. Proc. Natl. Acad. Sci. USA 2005, 102, 5460–5465. [Google Scholar] [CrossRef]
- Lambert, M.L.; Jacobs, I.; Osvath, M.; Von Bayern, A.M. Birds of a feather? Parrot and corvid cognition compared. Behaviour 2019, 156, 505–594. [Google Scholar] [CrossRef]
- Angelier, F.; Wingfield, J.C.; Tartu, S.; Chastel, O. Does prolactin mediate parental and life-history decisions in response to environmental conditions in birds? A review. Horm. Behav. 2016, 77, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Smiley, K.O.; Adkins-Regan, E. Factors that influence the onset of parental care in zebra finches: Roles for egg stimuli and prolactin. Behav. Process. 2018, 153, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Saha, I. Regulatory effects of prolactin on breeding and migratory behaviours in birds. South Asian J. Exp. Biol. 2021, 11, 337–344. [Google Scholar] [CrossRef]
- Maurer-Spurej, E. Circulating serotonin in vertebrates. Cell. Mol. Life Sci. 2005, 62, 1881–1889. [Google Scholar] [CrossRef]
- Van der Eijk, J.A.; Lammers, A.; Kjaer, J.B.; Rodenburg, T.B. Stress response, peripheral serotonin and natural antibodies in feather pecking genotypes and phenotypes and their relation with coping style. Physiol. Behav. 2019, 199, 1–10. [Google Scholar] [CrossRef]
- Hara, E.; Kubikova, L.; Hessler, N.A.; Jarvis, E.D. Role of the midbrain dopaminergic system in modulation of vocal brain activation by social context. Eur. J. Neurosci. 2007, 25, 3406–3416. [Google Scholar] [CrossRef]
- Chen, R.; Gadagkar, V.; Roeser, A.C.; Puzerey, P.A.; Goldberg, J.H. Movement signaling in ventral pallidum and dopaminergic midbrain is gated by behavioral state in singing birds. J. Neurophysiol. 2021, 125, 2219–2227. [Google Scholar] [CrossRef]
- Sasaki, A.; Sotnikova, T.D.; Gainetdinov, R.R.; Jarvis, E.D. Social Context-Dependent Singing-Regulated Dopamine. J. Neurosci. 2006, 26, 9010–9014. [Google Scholar] [CrossRef]
- Gadagkar, V.; Puzerey, P.A.; Chen, R.; Baird-Daniel, E.; Farhang, A.R.; Goldberg, J.H. Dopamine neurons encode performance error in singing birds. Science 2016, 354, 1278–1282. [Google Scholar] [CrossRef] [PubMed]
- Puig, M.; Rose, J.; Schmidt, R.; Freund, N. Dopamine modulation of learning and memory in the prefrontal cortex: Insights from studies in primates, rodents, and birds. Front. Neural Circuits 2014, 8, 93. [Google Scholar] [CrossRef] [PubMed]
- Rogers, L.J.; Vallortigara, G.; Andrew, R.J. Divided Brains: The Biology and Behaviour of Brain Asymmetries; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Pereira, A.G.; Farias, M.; Moita, M.A. Thalamic, cortical, and amygdala involvement in the processing of a natural sound cue of danger. PLoS Biol. 2020, 18, e3000674. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.-F.; Chaiken, M.; Zuo, M.; Miller, H. Nucleus Taenia of the Amygdala of Birds: Anatomical and Functional Studies in Ring Doves (Streptopelia risoria) and European Starlings (Sturnus vulgaris). Brain Behav. Evol. 1999, 53, 243–270. [Google Scholar] [CrossRef]
- Agnvall, B.; Katajamaa, R.; Altimiras, J.; Jensen, P. Is domestication driven by reduced fear of humans? Boldness, metabolism and serotonin levels in divergently selected red junglefowl (Gallus gallus). Biol. Lett. 2015, 11, 20150509. [Google Scholar] [CrossRef]
- Rogers, L.J. Relevance of brain and behavioural lateralization to animal welfare. Appl. Anim. Behav. Sci. 2010, 127, 1–11. [Google Scholar] [CrossRef]
- Seibert, L.M. Social Behavior of Psittacine Birds. In Manual of Parrot Behavior; Luescher, A.U., Ed.; Blackwell Publishing: Ames, IA, USA, 2006; pp. 43–48. [Google Scholar] [CrossRef]
- Policht, R.; Kowalczyk, A.; Łukaszewicz, E.; Hart, V. Hissing of geese: Caller identity encoded in a non-vocal acoustic signal. PeerJ 2020, 8, e10197. [Google Scholar] [CrossRef]
- Kaplan, G. Bird Bonds: Sex, Mate-Choice and Cognition in Australian Native Birds; (See especially section on emotions); CSIRO Publishing: Melbourne, VIC, Australia, 2019. [Google Scholar]
- McFarland, D.; Budgell, P. The thermoregulatory role of feather movements in the barbary dove (Streptopelia risoria). Physiol. Behav. 1970, 5, 763–771. [Google Scholar] [CrossRef]
- Ruxton, G.D.; Persons IV, W.S.; Currie, P.J. A continued role for signaling functions in the early evolution of feathers. Evolution 2017, 71, 797–799. [Google Scholar] [CrossRef]
- Bertin, A.; Beraud, A.; Lansade, L.; Mulot, B.; Arnould, C. Bill covering and nape feather ruffling as indicators of calm states in the Sulphur-crested cockatoo (Cacatua galerita). Behav. Process. 2020, 178, 104188. [Google Scholar] [CrossRef]
- Mellen, J.; Macphee, M.S. Philosophy of environmental enrichment: Past, present, and future. Zoo Biol. 2001, 20, 211–226. [Google Scholar] [CrossRef]
- Wells, D.L. Sensory stimulation as environmental enrichment for captive animals: A review. Appl. Anim. Behav. Sci. 2009, 118, 1–11. [Google Scholar] [CrossRef]
- Williams, I.; Hoppitt, W.; Grant, R. The effect of auditory enrichment, rearing method and social environment on the behavior of zoo-housed psittacines (Aves: Psittaciformes); implications for welfare. Appl. Anim. Behav. Sci. 2017, 186, 85–92. [Google Scholar] [CrossRef]
- Slater, M.N.; Hauber, M.E. Olfactory enrichment and scent cue associative learning in captive birds of prey. Zoo Biol. 2017, 36, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Potier, S.; Duriez, O.; Cunningham, G.B.; Bonhomme, V.; O’Rourke, C.; Fernández-Juricic, E.; Bonadonna, F. Visual field shape and foraging ecology in diurnal raptors. J. Exp. Biol. 2018, 221, jeb.177295. [Google Scholar] [CrossRef] [PubMed]
- Shepherdson, D. The role of environmental enrichment in the captive breeding and reintroduction of endangered species. In Creative Conservation; Springer Science and Business Media LLC.: Berlin, Germany, 1994; pp. 167–177. [Google Scholar]
- De Almeida, A.C.; Palme, R.; Moreira, N. How environmental enrichment affects behavioral and glucocorticoid responses in captive blue-and-yellow macaws (Ara ararauna). Appl. Anim. Behav. Sci. 2018, 201, 125–135. [Google Scholar] [CrossRef]
- Rojas-Ferrer, I.; Morand-Ferron, J. The impact of learning opportunities on the development of learning and decision-making: An experiment with passerine birds. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190496. [Google Scholar] [CrossRef]
- Meehan, C.L.; Garner, J.P.; Mench, J.A. Environmental enrichment and development of cage stereotypy in Orange-winged Amazon parrots (Amazona amazonica). Dev. Psychobiol. 2004, 44, 209–218. [Google Scholar] [CrossRef]
- Breen, A.J.; Lovie, K.E.; Guerard, C.; Edwards, S.C.; Cooper, J.; Healy, S.D.; Guillette, L.M. Juvenile socio-ecological environment shapes material technology in nest-building birds. Behav. Ecol. 2020, 31, 892–901. [Google Scholar] [CrossRef]
- Melfi, V.A. There are big gaps in our knowledge, and thus approach, to zoo animal welfare: A case for evidence-based zoo animal management. Zoo Biol. 2009, 28, 574–588. [Google Scholar] [CrossRef]
- Dunn, R.A. The Sooner the Better: The Arguments for the Use of Extended Welfare Assessment Grids in Animal Welfare Cases. Liverp. Law Rev. 2020, 41, 107–127. [Google Scholar] [CrossRef]
- Binding, S.; Farmer, H.; Krusin, L.; Cronin, K. Status of animal welfare research in zoos and aquariums: Where are we, where to next? J. Zoo Aquar. Res. 2020, 8, 166–174. [Google Scholar]
- Powell, D.M.; Watters, J.V. The Evolution of the Animal Welfare Movement in U.S. Zoos and Aquariums. Der Zool. Gart. 2017, 86, 219–234. [Google Scholar] [CrossRef]
- Maple, T.L.; Perdue, B.M. (Eds.) Zoo Animal Welfare; Springer: Berlin, Germany, 2013. [Google Scholar]
- Miller, L.J.; Vicino, G.A.; Sheftel, J.; Lauderdale, L.K. Behavioral Diversity as a Potential Indicator of Positive Animal Welfare. Animals 2020, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Appleby, M.; Hughes, B. Cages modified with perches and nests for the improvement of bird welfare. World’s Poult. Sci. J. 1990, 46, 38–40. [Google Scholar] [CrossRef]
- Reinert, S.E. Use of introduced perches by raptors: Experimental results and management implications. Raptor Res. 1984, 18, 25–29. [Google Scholar]
- Widén, P. Habitat Quality for Raptors: A Field Experiment. J. Avian Biol. 1994, 25, 219. [Google Scholar] [CrossRef]
- Tomé, R.; Dias, M.P.; Chumbinho, A.C.; Bloise, C. Influence of Perch Height and Vegetation Structure on the Foraging Behaviour of Little Owls Athene noctua: How to Achieve the Same Success in Two Distinct Habitats. Ardea 2011, 99, 17–26. [Google Scholar] [CrossRef]
- Van Hoek, C.S.; Ten Cate, C. Abnormal behavior in caged birds kept as pets. Appl. Anim. Welf. Sci. 1998, 1, 51–64. [Google Scholar] [CrossRef]
- Peng, S.; Broom, D.M. The Sustainability of Keeping Birds as Pets: Should Any Be Kept? Animals 2021, 11, 582. [Google Scholar] [CrossRef]
- Melfi, V.A.; Bowkett, A.; Plowman, A.B.; Pullen, K. Do zoo designers know enough about animals. In Innovation or Replication: Proceedings of the 6th International Symposium on Zoo Design; Whitley Wildlife Conservation Trust: Paignton, UK, 2005. [Google Scholar]
- Abrey, A. The management of a multi-species bird collection in a zoological park. In Avian Medicine; Butterworth-Heinemann: Oxford, UK, 2000; pp. 364–385. [Google Scholar] [CrossRef]
- Foulds-Davis, Y.L. Evaluating Breeding Performance in Mixed Species Bird Enclosures within European Zoos. Master’s Thesis, Liverpool John Moores University, Liverpool, UK, 2015. [Google Scholar]
- Dowes, K. Is there a visitor effect on behaviour and enclosure use of mixed bird species in a zoo enclosure? Plymouth Stud. Sci. 2012, 5, 38–60. [Google Scholar]
- Blanchett, M.; Finegan, E.; Atkinson, J. The Effects of Increasing Visitor and Noise Levels on Birds Within a Free-flight Aviary Examined Through Enclosure Use and Behavior. Anim. Behav. Cogn. 2020, 7, 49–69. [Google Scholar] [CrossRef]
- Lormant, F.; Ferreira, V.H.B.; Meurisse, M.; Lemarchand, J.; Constantin, P.; Morisse, M.; Cornilleau, F.; Parias, C.; Chaillou, E.; Bertin, A.; et al. Emotionality modulates the impact of chronic stress on memory and neurogenesis in birds. Sci. Rep. 2020, 10, 14620. [Google Scholar] [CrossRef] [PubMed]
- Pearce, D.; Pryke, S.R.; Griffith, S.C. Interspecific Aggression for Nest Sites: Model Experiments with Long-Tailed Finches (Poephila Acuticauda) and Endangered Gouldian Finches (Erythrura Gouldiae). The Auk 2011, 128, 497–505. [Google Scholar] [CrossRef]
- Yamamoto, J.T.; Shields, K.M.; Millam, J.R.; Roudybush, T.E.; Grau, C.R. Reproductive activity of force-paired cockatiels (Nymphicus hollandicus). Auk 1989, 106, 86–93. [Google Scholar]
- Fox, R.A.; Millam, J.R. The Use of Ratings and Direct Behavioural Observation to Measure Temperament Traits in Cockatiels (Nymphicus hollandicus). Ethology 2010, 116, 59–75. [Google Scholar] [CrossRef]
- Dhondt, A.A. Interspecific Competition in Birds; Oxford University Press: Oxford, UK, 2012; Volume 2. [Google Scholar]
- Harris, M.R.; Siefferman, L. Interspecific Competition Influences Fitness Benefits of Assortative Mating for Territorial Aggression in Eastern Bluebirds (Sialia sialis). PLoS ONE 2014, 9, e88668. [Google Scholar] [CrossRef]
- Fox, R.A.; Millam, J.R. Personality traits of pair members predict pair compatibility and reproductive success in a socially monogamous parrot breeding in captivity. Zoo Biol. 2014, 33, 166–172. [Google Scholar] [CrossRef]
- Covas, R.; Doutrelant, C.; Du Plessis, M.A. Experimental evidence of a link between breeding conditions and the decision to breed or to help in a colonial cooperative bird. Proc. R. Soc. B Biol. Sci. 2004, 271, 827–832. [Google Scholar] [CrossRef]
- Griffith, S.C.; Crino, O.L.; Andrew, S.C.; Nomano, F.Y.; Adkins-Regan, E.; Alonso-Alvarez, C.; Bailey, I.E.; Bittner, S.S.; Bolton, P.E.; Boner, W.; et al. Variation in reproductive success across captive populations: Methodological differences, potential biases and opportunities. Ethology 2017, 123, 1–29. [Google Scholar] [CrossRef]
- Andrew, S.; Munro, U. Cannibalism in the Australian pelican (Pelecanus conspicillatus) and Australian white ibis (Threskiornis molucca). Waterbirds 2008, 31, 632–635. [Google Scholar]
- Klausen, B. A mixed-species exhibit for African water birds (including pelicans, flamingos, spoonbills and storks) at Odense Zoo, Denmark: Breeding success, animal welfare and education. Int. Zoo Yearb. 2013, 48, 61–68. [Google Scholar] [CrossRef]
- Goss-Custard, J.D.; Triplet, P.; Sueur, F.; West, A.D. Critical thresholds of disturbance by people and raptors in foraging wading birds. Biol. Conserv. 2006, 127, 88–97. [Google Scholar] [CrossRef]
- Mumtaz, F.; Khan, M.I.; Zubair, M.; Dehpour, A.R. Neurobiology and consequences of social isolation stress in animal model—A comprehensive review. Biomed. Pharmacother. 2018, 105, 1205–1222. [Google Scholar] [CrossRef]
- Degernes, L.A.; Feduccia, A. Tenectomy of the Supracoracoideus Muscle to Deflight Pigeons (Columba livia) and Cockatiels (Nymphicus hollandicus). J. Avian Med. Surg. 2001, 15, 10–16. [Google Scholar] [CrossRef]
- Reese, L.; Baumgartner, K.; Von Fersen, L.; Merle, R.; Ladwig-Wiegard, M.; Will, H.; Haase, G.; Tallo-Parra, O.; Carbajal, A.; Lopez-Bejar, M.; et al. Feather Corticosterone Measurements of Greater Flamingos Living under Different Forms of Flight Restraint. Animals 2020, 10, 605. [Google Scholar] [CrossRef]
- Reese, L.; Ladwig-Wiegard, M.; von Fersen, L.; Haase, G.; Will, H.; Merle, R.; Encke, D.; Maegdefrau, H.; Baumgartner, K.; Thöne-Reineke, C. Deflighting zoo birds and its welfare considerations. Anim. Welf. 2020, 29, 69–80. [Google Scholar] [CrossRef]
- Anderson, M.J.; Robinson-Drummer, P.A. Examining the effects of directional orientation and deflighting on lateral behavior in captive Caribbean flamingos (Phoenicopterus ruber). J. Behav. Neurosci. Res. 2015, 12, 45–53. [Google Scholar]
- Rose, P.; Brereton, J.; Croft, D. Measuring welfare in captive flamingos: Activity patterns and exhibit usage in zoo-housed birds. Appl. Anim. Behav. Sci. 2018, 205, 115–125. [Google Scholar] [CrossRef]
- McMillan, F.D. (Ed.) Mental Health and Well-Being in Animals; CABI: Wallingford, UK, 2019. [Google Scholar]
- Jenkins, J.R. Feather picking and self-mutilation in psittacine birds. Vet. Clinics North Amer. Exotic Anim. Pract. 2001, 4, 651–667. [Google Scholar] [CrossRef]
- Association of Avian Veterinarians. Position Statement of the AAV on Permanent Anatomic Alterations of Avian Species; (Undated); Association of Avian Veterinarian: Denver, CO, USA; p. 1. Available online: https://cdn.ymaws.com/www.aav.org/resource/collection/5BC40BD0-9410-4924-AD81-30F0389E8435/alteration.pdf (accessed on 30 October 2021).
- Mallinson, J.J. A Sustainable Future for Zoos and Their Role in Wildlife Conservation. Hum. Dimens. Wildl. 2003, 8, 59–63. [Google Scholar] [CrossRef]
- Conde, D.A.; Flesness, N.; Colchero, F.; Jones, O.R.; Scheuerlein, A. An Emerging Role of Zoos to Conserve Biodiversity. Science 2011, 331, 1390–1391. [Google Scholar] [CrossRef] [PubMed]
- IUCN. IUCN Technical Guidelines on the Management of Ex-Situ Populations for Conservation; IUCN: Gland, Switzerland, 2002. [Google Scholar]
- Hoffmann, M.; Hilton-Taylor, C.; Angulo, A.; Böhm, M.; Brooks, T.M.; Butchart, S.H.M.; Carpenter, K.E.; Chanson, J.; Collen, B.; Cox, N.A.; et al. The Impact of Conservation on the Status of the World’s Vertebrates. Science 2010, 330, 1503–1509. [Google Scholar] [CrossRef]
- Pacífico, E.C.; Barbosa, E.A.; Filadelfo, T.; Oliveira, K.G.; Silveira, L.F.; Tella, J.L. Breeding to non-breeding population ratio and breeding performance of the globally Endangered Lear’s Macaw Anodorhynchus leari: Conservation and monitoring implications. Bird Conserv. Int. 2014, 24, 466–476. [Google Scholar] [CrossRef]
- Escalante-Pliego, P.; Arias-Montero, A.; Cortez-Contreras, E.; Cantú-Guzmán, J.C.; Rodríguez-Mouriño, C.M. Slow but sure assimilation to culture and ecotourism of reintroduced scarlet macaws in Los Tuxtlas, Veracruz, Mexico. J. Ecotourism 2019, 18, 368–377. [Google Scholar] [CrossRef]
- Vaughan, C.; Nemeth, N.M.; Cary, J.; Temple, S. Response of a Scarlet Macaw Ara macao population to conservation practices in Costa Rica. Bird Conserv. Int. 2005, 15, 119–130. [Google Scholar] [CrossRef]
- Trancoso, R. Changing Amazon deforestation patterns: Urgent need to restore command and control policies and market interventions. Environ. Res. Lett. 2021, 16, 041004. [Google Scholar] [CrossRef]
- Braverman, I. Conservation without nature: The trouble with in situ versus ex situ conservation. Geoforum 2014, 51, 47–57. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; Garcia, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human-induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef]
- Walters, J.R.; Derrickson, S.R.; Fry, D.M.; Haig, S.M.; Marzluff, J.M.; Wunderle, J.M., Jr. Status of the California condor (Gymnogyps californianus) and efforts to achieve its recovery. Auk 2010, 127, 969–1001. [Google Scholar] [CrossRef]
- Traylor-Holzer, K.; Leus, K.; Bauman, K. Integrated Collection Assessment and Planning (ICAP) workshop: Helping zoos move toward the One Plan Approach. Zoo Biol. 2019, 38, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Field, K.A.; Paquet, P.C.; Artelle, K.; Proulx, G.; Brook, R.K.; Darimont, C.T. Publication reform to safeguard wildlife from researcher harm. PLoS Biol. 2019, 17, e3000193. [Google Scholar] [CrossRef] [PubMed]
- Burden, N.; Chapman, K.; Sewell, F.; Robinson, V. Pioneering Better Science through the 3Rs: An Introduction to the National Centre for the Replacement, Refinement, and Reduction of Animals in Research (NC3Rs). J. Am. Assoc. Lab. Anim. Sci. 2015, 54, 198–208. [Google Scholar]
- Schechtman, L.M. Implementation of the 3Rs (refinement, reduction, and replacement): Validation and regulatory acceptance considerations for alternative toxicological test methods. ILAR J. 2002, 43 (Suppl. 1), S85–S94. [Google Scholar] [CrossRef] [PubMed]
- Charton, E.; Castle, P. Reduction, replacement and refinement of animal tests in the European Pharmacopoeia: Recent developments for monographs on biological substances and preparations. Pharmeuropa 2001, 13, 94–96. [Google Scholar]
- Bishop, L.J.; Nolen, A.L. Animals in research and education: Ethical issues. Kennedy Inst. Ethic. J. 2001, 11, 91–112. [Google Scholar] [CrossRef]
- Zemanova, M.A. Towards more compassionate wildlife research through the 3Rs principles: Moving from invasive to non-invasive methods. Wildl. Biol. 2020, 2020, 216291100. [Google Scholar] [CrossRef]
- Voss, M.; Shutler, D.; Werner, J. A Hard Look at Blood Sampling of Birds. Auk 2010, 127, 704–708. [Google Scholar] [CrossRef]
- Washburn, B.E.; Millspaugh, J.J.; Schulz, J.H.; Jones, S.B.; Mong, T. Using fecal glucocorticoids for stress assessment in mourning doves. Condor 2003, 105, 696–706. [Google Scholar] [CrossRef]
- Pérez, T.; Vázquez, J.F.; Quirós, F.; Domínguez, A. Improving non-invasive genotyping in capercaillie (Tetrao urogallus): Redesigning sexing and microsatellite primers to increase efficiency on faeces samples. Conserv. Genet. Resour. 2011, 3, 483–487. [Google Scholar] [CrossRef]
- Hoy, S.R.; Ball, R.E.; Lambin, X.; Whitfield, D.P.; Marquiss, M. Genetic markers validate using the natural phenotypic characteristics of shed feathers to identify individual northern goshawks Accipiter gentilis. J. Avian Biol. 2015, 47, 443–447. [Google Scholar] [CrossRef]
- Fairhurst, G.D.; Frey, M.D.; Reichert, J.F.; Szelest, I.; Kelly, D.M.; Bortolotti, G.R. Does environmental enrichment reduce stress? An integrated measure of corticosterone from feathers provides a novel perspective. PLoS ONE 2011, 6, e17663. [Google Scholar] [CrossRef]
- Vallant, S.; Niederstätter, H.; Berger, B.; Lentner, R.; Parson, W. Increased DNA typing success for feces and feathers of capercaillie (Tetrao urogallus) and black grouse (Tetrao tetrix). Ecol. Evol. 2018, 8, 3941–3951. [Google Scholar] [CrossRef] [PubMed]
- Dalén, L.; Götherström, A.; Meijer, T.; Shapiro, B. Recovery of DNA from Footprints in the Snow. Can. Field Nat. 2007, 121, 321–324. [Google Scholar] [CrossRef]
- Olah, G.; Heinsohn, R.G.; Brightsmith, D.J.; Espinoza, J.R.; Peakall, R. Validation of non-invasive genetic tagging in two large macaw species (Ara macao and A. chloropterus) of the Peruvian Amazon. Conserv. Genet. Resour. 2016, 8, 499–509. [Google Scholar] [CrossRef]
- Breed, D.; Meyer, L.C.R.; Steyl, J.C.A.; Goddard, A.; Burroughs, R.; Kohn, T.A. Conserving wildlife in a changing world: Understanding capture myopathy—A malignant outcome of stress during capture and translocation. Conserv. Physiol. 2019, 7, coz027. [Google Scholar] [CrossRef]
- Hanley, C.S.; Thomas, N.J.; Paul-Murphy, J.; Hartup, B.K. Exertional Myopathy in Whooping Cranes (Grus americana) with Prognostic Guidelines. J. Zoo Wildl. Med. 2005, 36, 489–497. [Google Scholar] [CrossRef]
- Ruder, M.G.; Noel, B.L.; Bednarz, J.C.; Keel, M.K. Exertional Myopathy in Pileated Woodpeckers (Dryocopus pileatus) Subsequent to Capture. J. Wildl. Dis. 2012, 48, 514–516. [Google Scholar] [CrossRef]
- Rogers, D.I.; Battley, P.F.; Sparrow, J.; Koolhaas, A.; Hassell, C.J. Treatment of capture myopathy in shorebirds: A successful trial in northwestern Australia. J. Field Ornithol. 2004, 75, 157–164. [Google Scholar] [CrossRef]
- McEntire, M.S.; Sanchez, C.R. Multimodal drug therapy and physical rehabilitation in the successful treatment of capture myopathy in a lesser flamingo (Phoeniconaias minor). J. Avian Medic. Surg. 2017, 31, 232–238. [Google Scholar] [CrossRef]
- McGowan, P.J.; Traylor-Holzer, K.; Leus, K. IUCN guidelines for determining when and how ex situ management should be used in species conservation. Conserv. Lett. 2017, 10, 361–366. [Google Scholar] [CrossRef]
- Zink, R.M.; Kale, H.W. Conservation genetics of the extinct dusky seaside sparrow Ammodramus maritimus nigrescens. Biol. Conserv. 1995, 74, 69–71. [Google Scholar] [CrossRef]
- Snyder, N.F.; Derrickson, S.R.; Beissinger, S.R.; Wiley, J.W.; Smith, T.B.; Toone, W.D.; Miller, B. Limitations of Captive Breeding in Endangered Species Recovery. Conserv. Biol. 1996, 10, 338–348. [Google Scholar] [CrossRef]
- National Research Council. The Scientific Bases for the Preservation of the Hawaiin Crow; National Academy Press: Washington, DC, USA, 1992. [Google Scholar]
- Tanimoto, A.M. The Vocal Repertoire of the Alāl (Corvus hawaiiensis) during Breeding Season: A Comparison between Current Captive and Past Wild Populations of the Native Hawaiian Crow. Ph.D. Thesis, University of Hawaii at Hilo, Hilo, HI, USA, 2014. [Google Scholar]
- Lloyd, N.; Traylor-Holzer, K.; Mickelberg, J.; Stephens, T.; Schroeder, M.; Heinrichs, J.; Diaz, B.P.; Moehrenschlager, A. Greater Sage-Grouse in Canada Population and Habitat Viability Assessment Workshop. In Final Report; IUCN SSC Conservation Breeding Specialist Group: Apple Valley, CA, USA; Minnesota, MN, USA, 2014. [Google Scholar]
- Coates, P.S.; Brussee, B.E.; Ricca, M.A.; Dudko, J.E.; Prochazka, B.G.; Espinosa, S.P.; Casazza, M.L.; Delehanty, D.J. Greater Sage-grouse (Centrocercus urophasianus) Nesting and Brood-Rearing Microhabitat in Nevada and Cali-Fornia—Spatial Variation in Selection and Survival Patterns (No. 2017–1087); US Geological Survey: Reston, VA, USA, 2007. [CrossRef]
- Parsons, L.A.; Runia, T.J.; Vincent, G.P.; Gregory, A.J.; Jenks, J.A. Greater Sage-Grouse survival varies with breeding season events in West Nile virus non-outbreak years. Condor 2021, 123, duab002. [Google Scholar] [CrossRef]
- LeBeau, C.W.; Johnson, G.D.; Holloran, M.J.; Beck, J.L.; Nielson, R.M.; Kauffman, M.E.; Rodemaker, E.J.; McDonald, T.L. Greater sage-grouse habitat selection, survival, and wind energy infrastructure. J. Wildl. Managm. 2017, 81, 690–711. [Google Scholar] [CrossRef]
- Hagen, C.A.; Sedinger, J.E.; Braun, C.E. Estimating sex-ratio, survival, and harvest susceptibility in greater sage-grouse: Making the most of hunter harvests. Wildl. Biol. 2018, 2018. [Google Scholar] [CrossRef]
- Foster, L.J.; Dugger, K.M.; Hagen, C.A.; Budeau, D.A. Potential effects of GPS transmitters on greater sage-grouse survival in a post-fire landscape. Wildl. Biol. 2018, 2018. [Google Scholar] [CrossRef]
- Dolman, P.M.; Collar, N.J.; Scotland, K.M.; Burnside, R.J. Ark or park: The need to predict relative effectiveness of ex situ and in situ conservation before attempting captive breeding. J. Appl. Ecol. 2015, 52, 841–850. [Google Scholar] [CrossRef]
- Cordero-Rivera, A. Behavioral Diversity (Ethodiversity): A Neglected Level in the Study of Biodiversity. Front. Ecol. Evol. 2017, 5, 7. [Google Scholar] [CrossRef]
- Curio, E. Conservation needs ethologv. Trends Ecol. Evol. 1996, 11, 260–263. [Google Scholar] [CrossRef]
- Griffin, A.S.; Galef, B.G., Jr. Social learning about predators: Does timing matter? Anim. Behav. 2005, 69, 669–678. [Google Scholar] [CrossRef]
- Curio, E. Cultural Transmission of Enemy Recognition by Birds. In Social Learning: Psychological and Biological Perspectives; Zentall, T.R., Galef, B.G., Jr., Eds.; Psychology Press: Hove, UK, 1988; pp. 75–97. [Google Scholar]
- Curio, E.; Ernst, U.; Vieth, W. The adaptive significance of avian mobbing. II. Cultural Transmission of enemy recognition in blackbirds: Effectiveness and some constraints. Z. Tierpsychol. 1978, 48, 184–202. [Google Scholar] [CrossRef]
- Slagsvold, T.; Wiebe, K.L. Social learning in birds and its role in shaping a foraging niche. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 969–977. [Google Scholar] [CrossRef]
- Janik, V.M.; Slater, P.J. The different roles of social learning in vocal communication. Anim. Behav. 2000, 60, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Guillette, L.M.; Scott, A.C.Y.; Healy, S. Social learning in nest-building birds: A role for familiarity. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152685. [Google Scholar] [CrossRef]
- Potvin, D.A.; Ratnayake, C.P.; Radford, A.N.; Magrath, R.D. Birds Learn Socially to Recognize Heterospecific Alarm Calls by Acoustic Association. Curr. Biol. 2018, 28, 2632.e4–2637.e4. [Google Scholar] [CrossRef] [PubMed]
- Guillette, L.M.; Healy, S.D. Social learning in nest-building birds watching live-streaming video demonstrators. Integr. Zool. 2019, 14, 204–213. [Google Scholar] [CrossRef]
- Crates, R.; Rayner, L.; Stojanovic, D.; Webb, M.; Terauds, A.; Heinsohn, R. Contemporary breeding biology of critically endangered Regent Honeyeaters: Implications for conservation. Ibis 2018, 161, 521–532. [Google Scholar] [CrossRef]
- Schwab, C.; Bugnyar, T.; Schloegl, C.; Kotrschal, K. Enhanced social learning between siblings in common ravens, Corvus corax. Anim. Behav. 2008, 75, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Uomini, N.; Fairlie, J.; Gray, R.D.; Griesser, M. Extended parenting and the evolution of cognition. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190495. [Google Scholar] [CrossRef]
- Dall, S.R.; Wright, J. Rich pickings near large communal roosts favor ‘gang’ foraging by juvenile common ravens, Corvus corax. PLoS ONE 2009, 4, e4530. [Google Scholar] [CrossRef] [PubMed]
- Taylor, G.; Ewen, J.G.; Clarke, R.H.; Blackburn, T.M.; Johnson, G.; Ingwersen, D. Video monitoring reveals novel threat to Critically Endangered captive-bred and released Regent Honeyeaters. Emu. Austral Ornithol. 2018, 118, 304–310. [Google Scholar] [CrossRef]
- Tilgar, V.; Moks, K.; Saag, P. Predator-induced stress changes parental feeding behavior in pied flycatchers. Behav. Ecol. 2010, 22, 23–28. [Google Scholar] [CrossRef]
- Perez, R.M.; Wilson, D.E.; Gruen, K.D. Survival and flight characteristics of captive-reared and wild northern bobwhite in South Texas. In Quail V: Proceedings of the Fifth National Quail Symposium; De Maso, S.J., Kuvleksy, W.P., Jr., Hernández, F., Berger, M.E., Eds.; Texas Parks and Wildlife Department: Austin, TX, USA, 2002; pp. 81–85. [Google Scholar]
- Berger-Tal, O.; Blumstein, D.T.; Carroll, S.; Fisher, R.N.; Mesnick, S.L.; Owen, M.A.; Saltz, D.; Claire, C.C.S.; Swaisgood, R.R. A systematic survey of the integration of animal behavior into conservation. Conserv. Biol. 2015, 30, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Barron, D.G.; Brawn, J.D.; Weatherhead, P.J. Meta-analysis of transmitter effects on avian behaviour and ecology. Methods Ecol. Evol. 2010, 1, 180–187. [Google Scholar] [CrossRef]
- Schlicht, E.; Kempenaers, B. The immediate impact of ringing, blood sampling and PIT Tags on the behaviour of blue tits Cyanistes caeruleus. Ardea 2018, 106, 39–98. [Google Scholar] [CrossRef]
- Lameris, T.K.; Müskens, G.J.D.M.; Kölzsch, A.; Dokter, A.M.; van der Jeugd, H.; Nolet, B.A. Effects of harness-attached tracking devices on survival, migration, and reproduction in three species of migratory waterfowl. Anim. Biotelem. 2018, 6, 7. [Google Scholar] [CrossRef]
- Pakanen, V.; Rönkä, N.; Leslie, T.R.; Blomqvist, D.; Koivula, K. Survival probability in a small shorebird decreases with the time an individual carries a tracking device. J. Avian Biol. 2020, 51, e02555. [Google Scholar] [CrossRef]
- Taylor, F.W. Scientific Management; Routledge: London, UK, 2003. [Google Scholar]
- Buechley, E.R.; Oppel, S.; Beatty, W.S.; Nikolov, S.C.; Dobrev, V.; Arkumarev, V.; Saravia, V.; Bougain, C.; Bounas, A.; Kret, E.; et al. Identifying critical migratory bottlenecks and high-use areas for an endangered migratory soaring bird across three continents. J. Avian Biol. 2018, 49, e01629. [Google Scholar] [CrossRef]
- McConnell, C. Identifying, Protecting and Managing Stopover Habitats for Wild Whooping Cranes on US Army Corps of Engineers Lakes. bioRxiv 2021. [Google Scholar] [CrossRef]
- Liu, Z.; Huang, Q.; Tang, G. Identification of urban flight corridors for migratory birds in the coastal regions of Shenzhen city based on three-dimensional landscapes. Landsc. Ecol. 2020, 36, 2043–2057. [Google Scholar] [CrossRef]
- Bauer, S.; Shamoun-Baranes, J.; Nilsson, C.; Farnsworth, A.; Kelly, J.F.; Reynolds, D.R.; Dokter, A.M.; Krauel, J.F.; Petterson, L.B.; Horton, K.G.; et al. The grand challenges of migration ecology that radar aeroecology can help answer. Ecography 2019, 42, 861–875. [Google Scholar] [CrossRef]
- Meyburg, B.U.; Angelov, I.; Azar, S. A corridor of soaring bird migration in Lebanon on the Eastern Mediterranean flyway. Sandgrouse 2020, 42, 46–58. [Google Scholar]
- Postman, N. Technopoly: The Surrender of Culture to Technology; Vintage Bks: New York, NY, USA, 2011. [Google Scholar]
- Kavelaars, M.M.; Stienen, E.; Matheve, H.; Buijs, R.-J.; Lens, L.; Müller, W. GPS tracking during parental care does not affect early offspring development in lesser black-backed gulls. Mar. Biol. 2018, 165, 87. [Google Scholar] [CrossRef]
- Martínez, A.E.; Parra, E.; Muellerklein, O.; Vredenburg, V.T. Fear-based niche shifts in neotropical birds. Ecology 2018, 99, 1338–1346. [Google Scholar] [CrossRef]
- Rösner, S.; Mussard-Forster, E.; Lorenc, T.; Müller, J. Recreation shapes a “landscape of fear” for a threatened forest bird species in Central Europe. Landsc. Ecol. 2013, 29, 55–66. [Google Scholar] [CrossRef]
- Hill, J.M.; Elphick, C.S. Are grassland passerines especially susceptible to negative transmitter impacts? Wildl. Soc. Bull. 2011, 35, 362–367. [Google Scholar] [CrossRef]
- Michael, S.; Gartrell, B.; Hunter, S. Humeral remodeling and soft tissue injury of the wings caused by backpack harnesses for radio transmitters in New Zealand takahē (Porphyrio hochstetteri). J. Wildl. Dis. 2013, 49, 552–559. [Google Scholar] [CrossRef]
- Trefry, S.A.; Diamond, A.W.; Jesson, L.K. Wing marker woes: A case study and meta-analysis of the impacts of wing and patagial tags. J. Ornithol. 2013, 154, 1–11. [Google Scholar] [CrossRef]
- Vandenabeele, S.P.; Wilson, R.; Grogan, A. Tags on seabirds; how seriously are we considering instrument-induced behaviors? Anim. Welf. 2011, 20, 559–571. [Google Scholar]
- Alarcón-Nieto, G.; Graving, J.M.; Klarevas-Irby, J.A.; Maldonado-Chaparro, A.A.; Mueller, I.; Farine, D.R. An automated barcode tracking system for behavioural studies in birds. Methods Ecol. Evol. 2018, 9, 1536–1547. [Google Scholar] [CrossRef]
- Croll, D.A.; Osmek, S.D.; Bengtson, J.L. An Effect of Instrument Attachment on Foraging Trip Duration in Chinstrap Penguins. Condor 1991, 93, 777–779. [Google Scholar] [CrossRef]
- Croll, D.A.; Gaston, A.J.; Burger, A.E.; Konnoff, D. Foraging Behavior and Physiological Adaptation for Diving in Thick-Billed Murres. Ecology 1992, 73, 344–356. [Google Scholar] [CrossRef]
- Galván, I.; Barba, E.; Piculo, R.; Cantó, J.L.; Cortés, V.; Monrós, J.S.; Atiénzar, F.; Proctor, H. Feather mites and birds: An interaction mediated by uropygial gland size? J. Evolut. Biol. 2008, 21, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Rueda, G. Preen oil and bird fitness: A critical review of the evidence. Biol. Rev. 2017, 92, 2131–2143. [Google Scholar] [CrossRef]
- Daly, M.K.; Saba, C.F.; Crochik, S.S.; Howerth, E.W.; Kosarek, C.E.; Cornell, K.K.; Roberts, R.E.; Northrup, N.C. Fibrosarcoma adjacent to the site of microchip implantation in a cat. J. Feline Med. Surg. 2008, 10, 202–205. [Google Scholar] [CrossRef]
- Newman, S.H.; Carter, H.R.; Whitworth, D.L.; Zinkl, J.G. Health Assessments and stress response of Xantus’s Murrelets to capture, handling, and radio-marking. Mar. Ornithol. 2005, 33, 147–154. [Google Scholar]
- Heatley, J.J.; Oliver, J.W.; Hosgood, G.; Columbini, S.; Tully, T.N. Serum Corticosterone Concentrations in Response to Restraint, Anesthesia, and Skin Testing in Hispaniolan Amazon Parrots (Amazona ventralis). J. Avian Med. Surg. 2000, 14, 172–176. [Google Scholar] [CrossRef]
- Machin, K.L. Assessment of pain in captive and free-ranging ducks after intra-abdominal transmitter placement. In Proceedings of the American Association of Zoo Veterinarians Annual Conference, Omaha, NE, USA, 17–22 October 1998; pp. 470–473. [Google Scholar]
- Whidden, S.E.; Williams, C.T.; Breton, A.R.; Buck, C.L. Effects of transmitters on the reproductive success of Tufted Puffins. J. Field Ornithol. 2007, 78, 206–212. [Google Scholar] [CrossRef]
- Thaxter, C.B.; Ross-Smith, V.H.; Clark, J.A.; Clark, N.A.; Conway, G.J.; Masden, E.A.; Wade, H.M.; Leat, E.H.K.; Gear, S.C.; Marsh, M.; et al. Contrasting effects of GPS device and harness attachment on adult survival of Lesser Black-backed Gulls, Larus fuscus and Great Skuas, Stercorarius skua. Ibis 2016, 158, 279–290. [Google Scholar] [CrossRef]
- Animal Ethics Infolink. Radio Tracking and GPS Tracking. Animal Research Review Panel Guideline 9. 2020. Available online: http://www.animalethics.org.au/policies-and-guidelines/wildlife-research/radio-tracking (accessed on 18 June 2021).
- Acharya, S.; Sahoo, S.K. PLGA nanoparticles containing various anticancer agents and tumor delivery by EPR effect. Adv. Drug Deliv. Rev. 2011, 63, 170–183. [Google Scholar] [CrossRef]
- Fallow, P.M.; Chiaradia, A.; Ropert-Coudert, Y.; Kato, A.; Reina, R.D. Flipper Bands Modify the Short-Term Diving Behavior of Little Penguins. J. Wildl. Manag. 2009, 73, 1348–1354. [Google Scholar] [CrossRef]
- Hudson, S.A. Effect of wing tags and testing hypotheses of sexual size dimorphism in frigatebirds. Ph.D. Thesis, University of New Brunswick, Fredericton, NB, Canada, 2015. [Google Scholar]
- Calvo, B.; Furness, R. A review of the use and the effects of marks and devices on birds. Ringing Migr. 1992, 13, 129–151. [Google Scholar] [CrossRef]
- Uher-Koch, B.D.; Schmutz, J.A.; Wright, K.G. Nest visits and capture events affect breeding success of Yellow-billed and Pacific loons. Condor 2015, 117, 121–129. [Google Scholar] [CrossRef]
- Möcklinghoff, L.; Schuchmann, K.-L.; Marques, M.I. New non-invasive photo-identification technique for free-ranging giant anteaters (Myrmecophaga tridactyla) facilitates urgently needed field studies. J. Nat. Hist. 2018, 52, 2397–2411. [Google Scholar] [CrossRef]
- McLean, W.R.; Goldingay, R.L.; Westcott, D.A. Visual lures increase camera-trap detection of the southern cassowary (Casuarius casuarius johnsonii). Wildl. Res. 2017, 44, 230. [Google Scholar] [CrossRef]
- O’Brien, T.G.; Kinnaird, M.F. A picture is worth a thousand words: The application of camera trapping to the study of birds. Bird Conserv. Int. 2008, 18, S144–S162. [Google Scholar] [CrossRef]
- Fontúrbel, F.E.; Rodríguez-Gómez, G.B.; Fernández, N.; García, B.; Orellana, J.I.; Castaño-Villa, G.J. Sampling understory birds in different habitat types using point counts and camera traps. Ecol. Indic. 2020, 119, 106863. [Google Scholar] [CrossRef]
- Randler, C.; Kalb, N. Distance and size matters: A comparison of six wildlife camera traps and their usefulness for wild birds. Ecol. Evol. 2018, 8, 7151–7163. [Google Scholar] [CrossRef]
- Larkin, R.P.; Sutherland, P.J. Migrating birds respond to Project Seafarer’s electromagnetic field. Science 1977, 195, 777–779. [Google Scholar] [CrossRef]
- Harrop, S. Climate Change, Conservation and the Place for Wild Animal Welfare in International Law. J. Environ. Law 2011, 23, 441–462. [Google Scholar] [CrossRef]
- Geen, G.R.; Robinson, R.A.; Baillie, S.R. Effects of tracking devices on individual birds—A review of the evidence. J. Avian Biol. 2019, 50, e01823. [Google Scholar] [CrossRef]
- Guhad, F. Introduction to the 3Rs (refinement, reduction and replacement). J. Am. Assoc. Lab. Anim. Sci. 2005, 44, 58–59. [Google Scholar]
- Westphal, K.R. Back to the 3 R’s: Rights, Responsibilities and Reasoning. SATS 2016, 17, 21–60. [Google Scholar] [CrossRef]
- Rösner, S.; Brandl, R.; Segelbacher, G.; Lorenc, T.; Müller, J. Non-invasive genetic sampling allows estimation of capercaillie numbers and population structure in the Bohemian Forest. Eur. J. Wildl. Res. 2014, 60, 789–801. [Google Scholar] [CrossRef]
- Gould, J.; Clulow, J.; Clulow, S. Using citizen science in the photo-identification of adult individuals of an amphibian based on two facial skin features. PeerJ 2021, 9, e11190. [Google Scholar] [CrossRef]
- Zdenek, C.N. Who’s Who of Palm Cockatoos: Evaluating Non-Invasive Techniques for Identification of Individual Palm Cockatoos (Probosciger aterrimus). Master’s Thesis, Australian National University (ANU), Canberra, ACT, Australia, 2012. [Google Scholar] [CrossRef]
- Gaglio, D.; Cook, T.R.; McInnes, A.; Sherley, R.; Ryan, P.G. Foraging plasticity in seabirds: A non-invasive study of the diet of greater crested terns breeding in the Benguela region. PLoS ONE 2018, 13, e0190444. [Google Scholar] [CrossRef]
- Adam, I.; Scharff, C.; Honarmand, M. Who is Who? Non-invasive Methods to Individually Sex and Mark Altricial Chicks. J. Vis. Exp. 2014, 87, e51429. [Google Scholar] [CrossRef]
- Bekoff, M. The importance of ethics in conservation biology: Let’s be ethicists, not ostriches. Endanger. Spec. Upd. 2002, 19, 23–26. [Google Scholar]
- Defenders of Wildlife. Available online: https://defenders.org/about (accessed on 30 October 2021).
- Greggor, A.L.; Vicino, G.A.; Swaisgood, R.R.; Fidgett, A.; Brenner, D.; Kinney, M.E.; Farabaugh, S.; Masuda, B.; Lamberski, N. Animal Welfare in Conservation Breeding: Applications and Challenges. Front. Vet. Sci. 2018, 5, 323. [Google Scholar] [CrossRef]
- Carey, M.J.; Meathrel, C.E.; May, N.A. A new method for the long-term attachment of data-loggers to shearwaters (Procellariidae). Emu. Austral Ornithol. 2009, 109, 310–315. [Google Scholar] [CrossRef]
- Elliott, K.; McFarlane-Tranquilla, L.; Burke, C.; Hedd, A.; Montevecchi, W.; Anderson, W. Year-long deployments of small geolocators increase corticosterone levels in murres. Mar. Ecol. Prog. Ser. 2012, 466, 1–7. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Needs (Physical) | Natural Habits | Captivity | Potential Outcomes Challenges Solutions | Sources | ||
|---|---|---|---|---|---|---|
| 1. Foraging | Ground/arboreal | Most food in single bowl | Boredom, lack of motivation, feather picking | work for food | Food search/Food variety | Lindenmayer et al. 1996 [28]; Grindlinger and Ramsey 1991 [29]; Meehan et al. 2003 [30]; Marino 2018 [31] |
| 2. Flight | In most species, flight is a daily activity | Usually impossible | Muscle atrophy | Provide opportunity and motivation | Allow for regular exercise | Gaunt et al. 1990 [32] |
| 3. Light | natural | Great variation | stress | Avoiding artificial light | Avoid harsh and direct light by providing half dark corners | Mellor et al.2018 [33] |
| 4. Sleep | Birds are the only animals, outside of mammals, known to engage in slow wave sleep and REM sleep | Problem if kept in lit living rooms after dark or kept awake by sudden noises and bright or flashing lights | Aggression; Sleep deprivation; Loss of appetite | Non-threateningEnvironment without artificial light | Finding a warm and quiet spot with suitable night light | Lesku et al. 2011 [34]; Cooper et al. 2019; [35]; Cornelius et al. 2018 [36]; Hodinka et al. 2020 [37] |
| 5. Predictability | Daily routines | unpredictability | loss of control | Taking time tabling seriously | Good time tabling of extra activities | McMillan 2005 [38] |
| 6. Nutritional needs | All four food groups | Commonly deficient | Depression, weight loss, fractures, even vomiting and diarrhea | Creating variety | Early training in what is edible | Fisher 2013 [39] |
| 7. Noise | Forest and plains levels of sounds | Noisy people, radio, television, and especially ultrasonic or computer sounds | Stress, fear, shock | Avoiding sudden noises and nearness to noise-producingtechnology | Reducing the noise and removing the source | Baldwin et al. 2007 [40] |
| 8. Parental care | Substantial; Time involvement | Deprivation of parent care | Effect on neural development and adult functioning. Increased anxiety, impulsivity, aggression, and behavioral abnormalities such as motor stereotypies | Justification for separation now doubted | Absence of parental care has no panacea, but negative effects can be mitigated by providing mentors of the same species and/or environmental enrichment | Aengus and Millam 1999 [41]; Feenders and Bateson 2013 [42]; Mason and Rushen 2008 [43]; Greenwell and Montrose 2017 [44] |
| 9. Attachments | Within pairs or flocks, pairs remain in close spatial contact | Single, or mismatched; Multi-species | Depression, Physical signs of illness | Companion/buddy System | Companion/buddy System; Isosexual pair housing | South and Pruett-Jones 2000 [45]; Doane and Qualkinbush 1994 [46]; Meehan et al. 2003 [43]; Duque et al. 2020 [47]. |
| 10. Variety | natural | Lack thereof | Less evidence | To make variety species appropriate | Increasing meaningful variety | Seibert 2020 [22] |
| 11. Personality | Vastly Different | Ignored too often | A range of abnormal behavior, incl. screaming, aggression | Important to first establish compatibility in multi-housing | Let birds choose their partners by themselves | Zeigler-Hill and Highfill 2017 [48]; Richter and Hintze 2019 [49] |
| 12. Cognitive needs/brain function | daily life stimulation, problem solving and decision making | of increasing importance | Not much known other than effects on ‘mood’ | A good deal known about corvids and parrots but assessing cognitive needs is far more difficult | Can only be assessed via behavior and that presupposes knowing the cognitive skills very precisely | Bateson and Matheson 2007 [24]; Clark 2017 [50]; Hopper 2017 [51]; Rogers and Kaplan 2019 [52] |
| 13. Knowledge | Learning from mentors, parents, experience | Limited to captive environment | Unsuitable for release if in a captive breedingprogram | Provide challenges (animal agency) | Exposure to trial-and-error tasks | Spinka and Wemelsfelder 2018 [53] |
| 14. Experience stimulation | varied | limited | Boredom | Expand environment | Stimulation is sensory or environmental but can also be social | Pepperberg 1987 [54]; 1994 [55]; Evans 2001 [56]; Swendsen 2019 [57] |
| 15. Problem-solving | regular | --- | Birds in research facilities often get set tasks of this kind and are often better off than captive birds without such stimulation | To re-engage the individual with its environment | Often successfully used by making food more difficult to access, exploiting skills the species is known to have | Auersperg et al. 2018 [58]; Rössler et al. 2020 [59]; Laschober et al. 2021 [60] |
| 16. Exploration | daily | limited | Stop locomotion | Prevent depression | Create and change areas that lend themselves to exploration, such as tree segments with loose bark | Mettke-Hofmann et al. 2002 [61] |
| 17. Danger, approach to novelty | Recognizing danger/neophobia | loss of curiosity or loss of interest | Fear, anxiety, or indifference (non-recognition) | Settings conducive to avoid visual constants that might indicate danger | Places for hiding and height to flee to | Mettke-Hofmann et al. 2002 [61]; Papini et al. 2019 [62] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaplan, G. Casting the Net Widely for Change in Animal Welfare: The Plight of Birds in Zoos, Ex Situ Conservation, and Conservation Fieldwork. Animals 2022, 12, 31. https://doi.org/10.3390/ani12010031
Kaplan G. Casting the Net Widely for Change in Animal Welfare: The Plight of Birds in Zoos, Ex Situ Conservation, and Conservation Fieldwork. Animals. 2022; 12(1):31. https://doi.org/10.3390/ani12010031
Chicago/Turabian StyleKaplan, Gisela. 2022. "Casting the Net Widely for Change in Animal Welfare: The Plight of Birds in Zoos, Ex Situ Conservation, and Conservation Fieldwork" Animals 12, no. 1: 31. https://doi.org/10.3390/ani12010031
APA StyleKaplan, G. (2022). Casting the Net Widely for Change in Animal Welfare: The Plight of Birds in Zoos, Ex Situ Conservation, and Conservation Fieldwork. Animals, 12(1), 31. https://doi.org/10.3390/ani12010031