
Influence of Different Regimes of Moderate Maternal Feed Restriction during Pregnancy of Primiparous Rabbit Does on Long-Term Metabolic Energy Homeostasis, Productive Performance and Welfare

 , , , , , and
, , , , , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
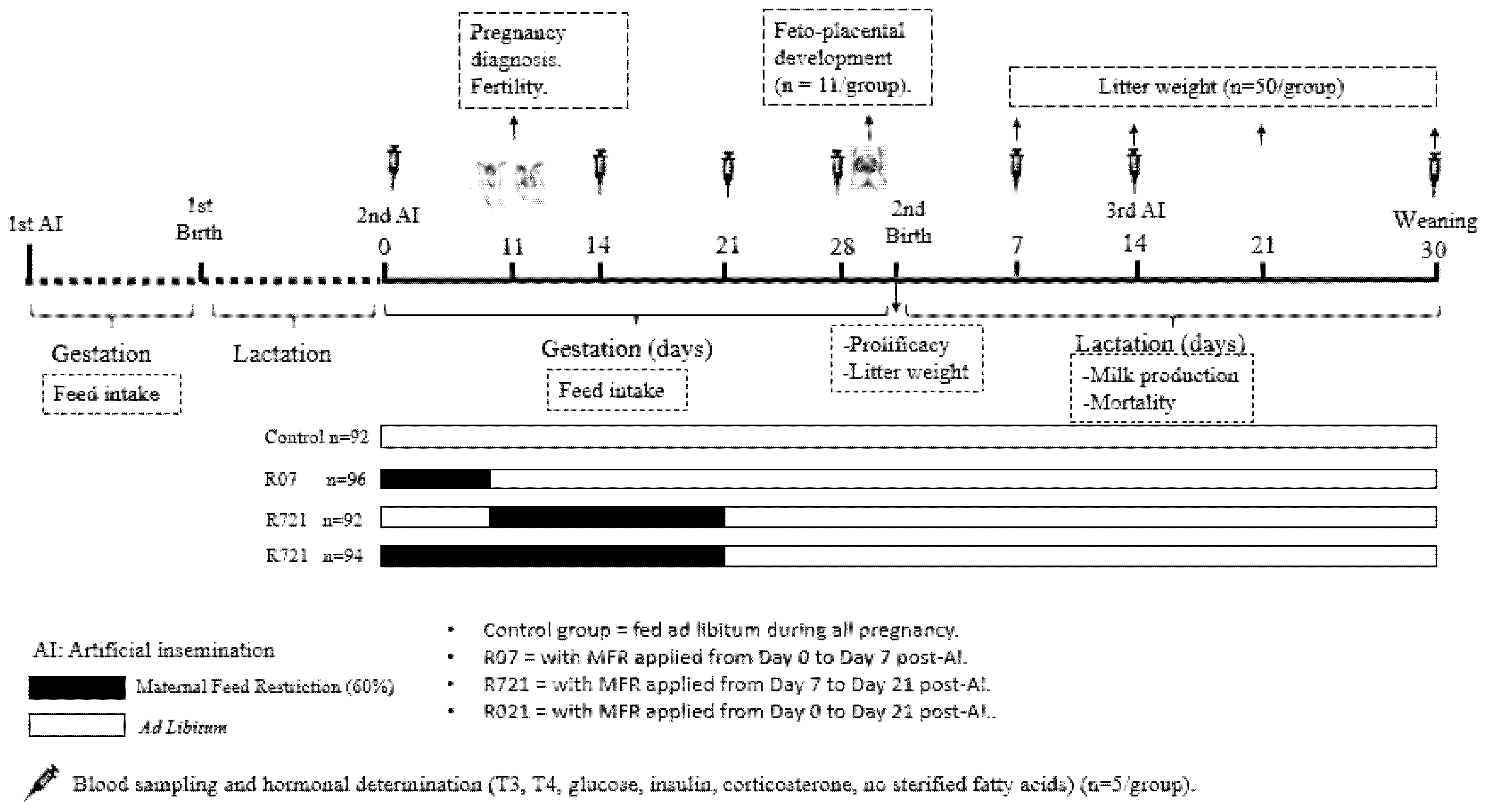
2.1. Experimental Design
- -
- Control group (n = 92) always fed ad libitum.
- -
- R07 (n = 96) with MFR applied from day 0 to day 7 post-AI.
- -
- R721 (n = 92) with MFR applied from day 7 to day 21 post-AI.
- -
- R021 (n = 94) with MFR applied from day 0 to day 21 post-AI.
2.2. Fetoplacental Study
2.3. Productive Outcome
2.4. Long-Term Maternal Metabolic and Hormonal Study
2.5. Statistical Analysis
3. Results
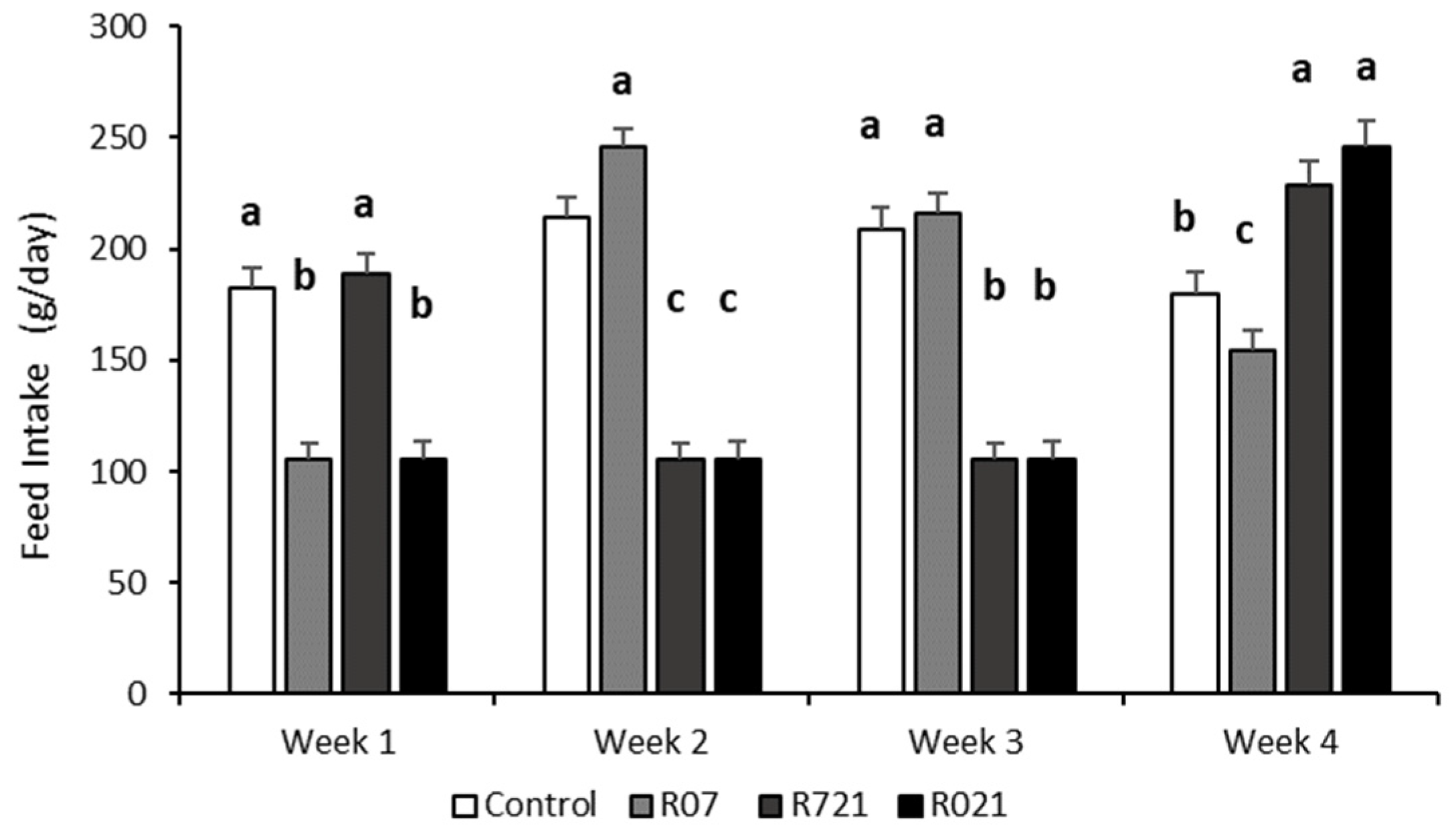
3.1. Feed Intake during Pregnancy
3.2. Fetoplacental Study
3.3. Productive Outcome
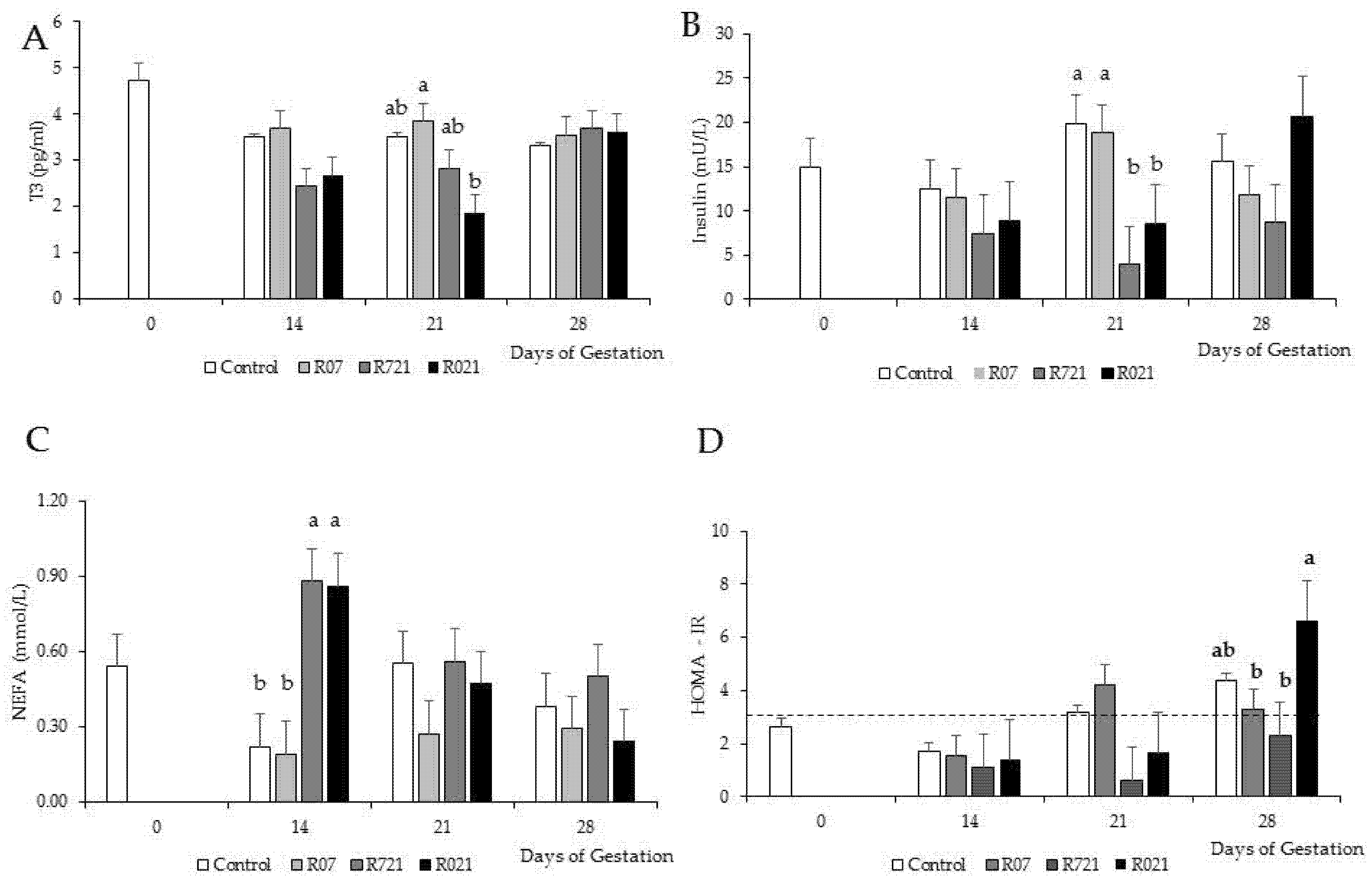
3.4. Long-Term Maternal Metabolic and Hormonal Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cartuche, L.; Pascual, M.; Gómez, E.A.; Blasco, A. Economic weights in rabbit meat production. World Rabbit. Sci. 2014, 22, 165–177. [Google Scholar] [CrossRef]
- Theau-Clément, M. Preparation of the rabbit doe to insemination: A review. World Rabbit. Sci. 2010, 15, 61–80. [Google Scholar] [CrossRef][Green Version]
- Fortun-Lamothe, L.; Rochambeau, H.; Lebas, F.; Tudela, F. Influence of the number of suckling young on reproductive per-formance in intensively rabbit does. In Proceedings of the 7th World Rabbit Congress, Valencia, Spain, 4–7 July 2000; pp. 125–132. [Google Scholar]
- Arias-Álvarez, M.; García-García, M.R.; Rebollar, P.G.; Revuelta, L.; Millán, P.; Lorenzo, P.L. Influence of metabolic status on oocyte quality and follicular characteristics at different post-partum periods in primiparous rabbit does. Theriogenology 2009, 72, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Sakr, O.G.; García-García, R.M.; Arias-Álvarez, M.; Lorenzo, P.L.; Millán, P.; Velasco, B.; Rebollar, P.G. Métodos de sincronización de celo en conejas primiparas lactantes a 25 días post-parto. Rev. Complut. Cienc. Vet. 2012, 6, 6–13. [Google Scholar]
- Partridge, G.G.; Lobley, G.E.; Fordyce, R.A. Energy and nitrogen metabolism of rabbits during pregnancy, lactation, and concurrent pregnancy and lactation. Br. J. Nutr. 1986, 56, 199–207. [Google Scholar] [CrossRef]
- Bauman, D.E.; Currie, W.B. Partitioning of Nutrients During Pregnancy and Lactation: A Review of Mechanisms Involving Homeostasis and Homeorhesis. J. Dairy Sci. 1980, 63, 1514–1529. [Google Scholar] [CrossRef]
- Rommers, J.M.; Kemp, B.; Meijerhof, R.; Noordhuizen, J.P.T.M. The effect of litter size before weaning on subsequent body development, feed intake, and reproductive performance of young rabbit does. J. Anim. Sci. 2001, 79, 1973–1982. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Tello, J.; Arias-Álvarez, M.; Jimenez-Martinez, M.A.; Garcia-Garcia, R.M.; Rodriguez, M.; Gonzalez, P.L.L.; Bermejo-Poza, R.; Gonzalez-Bulnes, A.; Rebollar, P.G. Competition for Materno-Fetal Resource Partitioning in a Rabbit Model of Undernourished Pregnancy. PLoS ONE 2017, 12, e0169194. [Google Scholar] [CrossRef]
- Partridge, G.G.; Daniels, Y.; Fordyce, R.A. The effects of energy intake during pregnancy in doe rabbits on pup birth weight, milk output and maternal body composition change in the ensuing lactation. J. Agric. Sci. 1986, 107, 697–708. [Google Scholar] [CrossRef]
- López-Tello, J.; Arias-Álvarez, M.; Jiménez-Martínez, M.A.; Barbero-fernábdez, A.; García, R.M.; Rodríguez, M.; Lorenzo, P.L.; Torres-Rovira, L.; Astiz, S.; González-Bulnes, A.; et al. The effects of sildenafil citrate and haemodynamics in a rabbit model of intaruterine growth restriction. Reprod. Fertil. Dev. 2016, 29, 1239–1248. [Google Scholar]
- Nafeaa, A.; Ahmed, S.A.E.; Fat Hallah, S. Effect of feed restriction during pregnancy on performance and productivity of New Zealand white rabbit does. Vet. Med. Int. 2011, 2011, 839737. [Google Scholar] [CrossRef][Green Version]
- Manal, A.F.; Tony, M.A.; Ezzo, O.H. Feed restriction of pregnant nulliparous rabbit does: Consequences on reproductive per-formance and maternal behaviour. Anim. Reprod. Sci. 2010, 120, 179–186. [Google Scholar] [CrossRef]
- Garcia-Garcia, R.; Arias-Alvarez, M.; Millan, P.; Francisco, M.R.; Rodriguez, A.S.; Lorenzo, P.; Rebollar, P. Gestation Food Restriction and Refeeding Compensate Maternal Energy Status and Alleviate Metabolic Consequences in Juvenile Offspring in a Rabbit Model. Nutrition 2021, 13, 310. [Google Scholar] [CrossRef]
- García-García, R.M.; Arias-Álvarez, M.; Rodríguez, M.; Sánchez-Rodríguez, A.; Formoso-Rafferty, N.; Lorenzo, P.L.; Rebollar, P.G. Effects of feed restriction during pregnancy on maternal reproductive outcome, foetal hepatic IGF gene expression and offspring performance in the rabbit. Animal 2021, in press. [Google Scholar]
- Mullur, R.; Liu, Y.-Y.; Brent, G.A. Thyroid Hormone Regulation of Metabolism. Physiol. Rev. 2014, 94, 355–382. [Google Scholar] [CrossRef] [PubMed]
- Nakrani, M.N.; Wineland, R.H.; Anjum, F. Physiology, Glucose Metabolism. In StatPearls; StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Aronoff, S.L.; Berkowitz, K.; Shreiner, B.; Want, L. Glucose Metabolism and Regulation: Beyond Insulin and Glucagon. Diabetes Spectr. 2004, 17, 183–190. [Google Scholar] [CrossRef]
- Emery, R.S.; Liesman, J.S.; Herdt, T.H. Metabolism of Long Chain Fatty Acids by Ruminant Liver. J. Nutr. 1992, 122, 832–837. [Google Scholar] [CrossRef]
- Brecchia, G.; Menchetti, L.; Cardinali, R.; Polisca, A.; Troisi, A.; Maranesi, M.; Boiti, C. Effects of fasting during pregnancy in rabbit does. In Proceedings of the 10th World Rabbit Congress, Sharm El-Sheikh, Egypt, 3–6 September 2012; pp. 341–345. [Google Scholar]
- Webster, J. Animal Welfare: Freedoms, Dominions and “A Life Worth Living”. Animals 2016, 6, 35. [Google Scholar] [CrossRef]
- Woodman, D. Reproductive Hormones In: Laboratory Animal Endocrinology; Wiley: Hoboken, NJ, USA, 1997; pp. 453–499. [Google Scholar]
- Helfenstein, T.; Fonseca, F.A.; Ihara, S.S.; Bottós, J.M.; Moreira, F.T.; Pott, H., Jr.; Farah, M.E.; Martins, M.C.; Izar, M.C. Impaired glucose tolerance plus hyperlip-idaemia induced by diet promotes retina microaneurysms in New Zealand rabbits. Int. J. Exp. Pathol. 2011, 92, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Enders, A.C.; Schlafke, S. Penetration of the uterine epithelium during implantation in the rabbit. Am. J. Anat. 1971, 132, 219–239. [Google Scholar] [CrossRef] [PubMed]
- Argente, M.J.; Santacreu, M.; Climent, A.; Blasco, A. Effects of intrauterine crowding on available uterine space per fetus in rabbits. Livest. Sci. 2008, 114, 211–219. [Google Scholar] [CrossRef]
- Godfrey, K.M.; Barker, D.J.; Aranceta, J.; Serra-Majem, L.; Ribas, L.; Pérez-Rodrigo, C. Fetal programming and adult health. Public Health Nutr. 2001, 4, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Matsuoka, T.; Mizoguchi, Y.; Serizawa, K.; Ishikura, T.; Mizuguchi, H.; Asano, Y. Effects of Stage and Degree of Restricted Feeding on Pregnancy Outcome in Rabbits. J. Toxicol. Sci. 2006, 31, 169–175. [Google Scholar] [CrossRef][Green Version]
- Rommers, J.M.; Meijerhof, R.; Noordhuizen, J.P.T.M.; Kemp, B. The effect of level of feeding in early gestation on reproductive success in young rabbit does. Anim. Reprod. Sci. 2004, 81, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Cappon, G.; Fleeman, T.; Chapin, R.; Hurtt, M. Effects of feed restriction during organogenesis on embryo-fetal development in rabbit. Birth Defects Res. Part B Dev. Reprod. Toxicol. 2005, 74, 424–430. [Google Scholar] [CrossRef]
- Clark, R.L.; Robertson, R.T.; Peter, C.P.; Bland, J.A.; Nolan, T.E.; Oppenheimer, L.; Bokelman, D.L. Association between adverse maternal and embryo-fetal effects in norfloxacin-treated and food-deprived rabbits. Fundam. Appl. Toxicol. 1986, 7, 272–286. [Google Scholar] [CrossRef]
- Robinson, J.S.; Owens, J.A.; Owens, P.C. Fetal Growth and Growth Retardation. Textbook of Fetal Physiology; Thorburn, G.D., Harding, R., Eds.; Oxford University Press: New York, NY, USA, 1994; pp. 83–94. [Google Scholar]
- Menchetti, L.; Brecchia, G.; Canali, C.; Cardinali, R.; Polisca, A.; Zerani, M.; Boiti, C. Food restriction during pregnancy in rabbits: Effects on hormones and metabolites involved in energy homeostasis and metabolic programming. Res. Vet.-Sci. 2015, 98, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Brecchia, G.; Bonanno, A.; Galeati, G.; Federici, C.; Maranesi, M.; Gobbetti, A.; Zerani, M.; Boiti, C. Hormonal and metabolic adaptation to fasting: Effects on the hypothalamic–pituitary–ovarian axis and reproductive performance of rabbit does. Domest. Anim. Endocrinol. 2006, 31, 105–122. [Google Scholar] [CrossRef]
- Hornick, J.; Van Eenaeme, C.; Gérard, O.; Dufrasne, I.; Istasse, L. Mechanisms of reduced and compensatory growth. Domest. Anim. Endocrinol. 2000, 19, 121–132. [Google Scholar] [CrossRef]
- Rommers, J.M.; Boiti, C.; Brecchia, G.; Meijerhof, R.; Noordhuizen, J.P.T.M.; Decuypere, E.; Kemp, B. Metabolic adaptation and hormonal regulation in young rabbit does during long-term caloric restriction and subsequent compensatory growth. Anim. Sci. 2004, 79, 255–264. [Google Scholar] [CrossRef]
- Buczkowska, E.O.; Jarosz-Chobot, P. Insulin effect on metabolism in skeletal muscles and the role of muscles in regulation of glucose homeostasis. Przegl. Lek. 2001, 58, 782–787. [Google Scholar]
- Fortun-Lamothe, L. Energy balance and reproductive performance in rabbit does. Anim. Reprod. Sci. 2006, 93, 1–15. [Google Scholar] [CrossRef]
- Garcia-Benasach, F. Diabetes Gestacional Análisis de la Influencia de Parámetros Clínicos y Ecográficos en los Resultados pe-rinatales. Ph.D. Thesis, Autonomic University of Madrid, Madrid, Spain, 2012. [Google Scholar]
- Buchanan, T.A.; Metzger, B.E.; Freinkel, N.; Bergman, R.N. Insulin sensitivity and B-cell responsiveness to glucose during late pregnancy in lean and moderately obese women with normal glucose tolerance or mild gestational diabetes. Am. J. Obstet. Gynecol. 1990, 162, 1008–1014. [Google Scholar] [CrossRef]
- Père, M.-C.; Etienne, M. Insulin sensitivity during pregnancy, lactation, and post-weaning in primiparous gilts1. J. Anim. Sci. 2007, 85, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Menchetti, L.; Andoni, E.; Barbato, O.; Canali, C.; Quattrone, A.; Vigo, D.; Codini, M.; Curone, G.; Brecchia, G. Energy homeostasis in rabbit does during pregnancy and pseudopregnancy. Anim. Reprod. Sci. 2020, 218, 106505. [Google Scholar] [CrossRef]
- Martínez-Paredes, E.; Ródenas, L.; Martínez-Vallespín, B.; Cervera, C.; Blas, E.; Brecchia, G.; Boiti, C.; Pascual, J.J. Effects of feeding programme on the performance and energy balance of nulliparous rabbit does. Animals 2012, 6, 1086–1095. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Habeeb, A.A.; EI-Masry, K.A. Hormonal pattern in pregnant rabbits and some productive aspects as affected by litter size at birth. 1991. Egypt J. Poult. Sci. 1991, 1, 429. [Google Scholar]
- Bober, M.A.; Dollah, A.; Becker, B.A.; Johnson, H.D. The influence of exogenous T3 on plasma prolactin, T3, TSH and milk production during heat stress in Holstein cows. J. Anim. Sci. 1980, 55, 339. [Google Scholar]
- Ganong, W.F. The Stress Response—A Dynamic Overview. Hosp. Pract. 1988, 23, 155–171. [Google Scholar] [CrossRef]
- Brunton, P.J.; Russell, J.A.; Douglas, A.J. Adaptive responses of the maternal hypothalamic-pituitary-adrenal axis during pregnancy and lactation. J. Neuroendocrinol. 2008, 20, 764–776. [Google Scholar] [CrossRef]
- Entringer, S.; Buss, C.; Shirtcliff, E.A.; Cammack, A.; Yim, I.S.; Chicz-DeMet, A.; Sandman, C.A.; Wadhwa, P.D. Attenuation of maternal psychophysiological stress responses and the maternal cortisol awakening response over the course of human pregnancy. Stress 2009, 13, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Tamashiro, K.L.; Moran, T.H. Perinatal environment and its influences on metabolic programming of offspring. Physiol. Behav. 2010, 100, 560–566. [Google Scholar] [CrossRef]
- Mastorakos, G.; Ilias, I. Maternal and Fetal Hypothalamic-Pituitary-Adrenal Axes During Pregnancy and Postpartum. Ann. N. Y. Acad. Sci. 2003, 997, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Szendrő, Z.; Trocino, A.; Hoy, S.; Xiccato, G.; Villagrá, A.; Maertens, L. A review of recent research outcomes on the housing of farmed domestic rabbits: Reproducing does. World Rabbit. Sci. 2019, 27, 1–14. [Google Scholar] [CrossRef]
- Kalaba, Z.M. Physiological Response and Stress Indicators of California Rabbits under Intensive Conditions in Egypt. Asian J. Poult. Sci. 2012, 6, 65–78. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Control n = 11 | R07 n = 11 | R721 n = 11 | R021 n = 11 | SEM | p Value | |
|---|---|---|---|---|---|---|
| LBW of does (g) | 4638 a | 4640 a | 4342 b | 4413 ab | 80.90 | 0.0254 |
| Corpora lutea | 12.2 | 12.3 | 13.1 | 13.4 | 0.58 | 0.4350 |
| Total fetuses/doe | 11.7 | 13.2 | 12.5 | 12.5 | 0.38 | 0.0814 |
| Implantation rate 1 (%) | 92.5 | 97 | 95.2 | 91 | 2.57 | 0.3684 |
| Fetal Viability 2 (%) | 94.6 | 84.9 | 90.3 | 90.1 | 9.13 | 0.0824 |
| Fetuses weights (g) | ||||||
| Total | 39.7 a | 38.4 ab | 37.4 b | 37.0 b | 1.78 | 0.0028 |
| Head | 9.32 a | 9.06 ab | 8.75 b | 8.71 b | 0.35 | 0.0001 |
| Trunk | 29.0 a | 28.0 ab | 27.0 b | 26.8 b | 1.43 | 0.0014 |
| Fetuses measures (mm) | ||||||
| CRL | 101.4 a | 99.5 ab | 98.7 b | 98.6 b | 1.83 | 0.0018 |
| OND | 29.0 a | 29.2 a | 28.7 ab | 28.3 b | 0.61 | 0.0032 |
| BPD | 19.3 | 19.2 | 19.3 | 19.0 | 0.52 | 0.5801 |
| TD | 20.6 b | 21.3 a | 20.8 b | 20.2 b | 0.71 | 0.0034 |
| Organs weights (g) | ||||||
| Brain | 0.913 | 0.901 | 0.892 | 0.901 | 0.04 | 0.6395 |
| Liver | 2.64 a | 2.39 b | 2.36 b | 2.39 b | 0.18 | 0.0097 |
| Heart | 0.221 | 0.207 | 0.219 | 0.209 | 0.02 | 0.0734 |
| Lungs | 0.134 a | 0.126 a | 0.122 b | 0.126 a | 0.08 | 0.0166 |
| Kidneys | 0.359 a | 0.337 ab | 0.323 b | 0.307 b | 0.02 | 0.0001 |
| Digestive tract | 0.208 | 0.215 | 0.207 | 0.197 | 0.17 | 0.0952 |
| Ratios | ||||||
| Brain | 0.024 | 0.024 | 0.024 | 0.025 | 0.00 | 0.4033 |
| Liver | 0.066 a | 0.061 b | 0.064 ab | 0.066 a | 0.00 | 0.0033 |
| Heart | 0.006 a | 0.005 b | 0.006 a | 0.006 a | 0.00 | 0.0028 |
| Lungs | 0.034 | 0.033 | 0.033 | 0.034 | 0.00 | 0.0942 |
| Kidneys | 0.009 a | 0.009 a | 0.009 a | 0.008 b | 0.00 | 0.0001 |
| Digestive tract | 0.053 | 0.056 | 0.056 | 0.053 | 0.00 | 0.1759 |
| Brain: Liver | 0.367 | 0.395 | 0.400 | 0.395 | 0.03 | 0.2273 |
| Control n = 11 | R07 n = 11 | R721 n = 11 | R021 n = 11 | SEM | p Value | |
|---|---|---|---|---|---|---|
| Whole placenta (g) | 5.16 ab | 5.21 a | 4.60 c | 4.81 bc | 0.30 | 0.0001 |
| Labyrinth | ||||||
| Weight (g) | 3.27 | 3.51 | 3.30 | 3.33 | 0.80 | 0.0809 |
| Length (mm) | 37.40 a | 35.80 a | 35.50 a | 33.90 b | 4.28 | 0.0001 |
| Width (mm) | 28.50 | 28.40 | 27.70 | 27.40 | 3.72 | 0.1286 |
| Thickness (mm) | 5.16 a | 5.17 a | 4.70 b | 5.05 ab | 0.99 | 0.0053 |
| Decidua | ||||||
| Weight (g) | 1.48 a | 1.55 a | 1.22 b | 1.40 a | 0.11 | 0.0001 |
| Length (mm) | 39.61 a | 37.32 b | 37.30 b | 36.09 b | 1.41 | 0.0001 |
| Width (mm) | 17.84 a | 18.76 a | 16.42 b | 18.39 a | 0.95 | 0.0001 |
| Thickness (mm) | 3.45 a | 3.52 a | 3.11 b | 3.27 ab | 0.26 | 0.0069 |
| Placental efficiency 1 | 7.52 ab | 7.34 b | 7.86 a | 7.45 ab | 0.34 | 0.0162 |
| Control | R07 | R721 | R021 | |||
|---|---|---|---|---|---|---|
| n = 92 | n = 96 | n = 92 | n = 94 | RMSE | p Value | |
| Fertility 1 (%) | 71.11 | 76.04 | 78.26 | 78.72 | 0.6129 | |
| Parturitions | 59 | 69 | 67 | 68 | ||
| Litter size | ||||||
| Born alive | 11.01 | 10.98 | 10.05 | 11.02 | 3.17 | 0.2904 |
| Stillborn | 0.50 | 0.43 | 0.32 | 0.24 | 1.00 | 0.5514 |
| Weaned (30 days post-partum) | 10.20 a | 10.54 a | 9.59 b | 10.04 ab | 1.14 | 0.0004 |
| Litter weight (g) 2 | ||||||
| At birth | 605.70 | 619.34 | 580.93 | 643.98 | 142.6 | 0.1276 |
| At 7 days post-partum | 1306 | 1361 | 1272 | 1288 | 216.2 | 0.1646 |
| At 14 days post-partum | 2290 | 2398 | 2260 | 2276 | 318.1 | 0.1040 |
| At 21 days p post-partum p | 3314 | 3394 | 3227 | 3304 | 381.2 | 0.1613 |
| At 30 days post-partum (weaning) | 5782 | 5708 | 5447 | 5484 | 1129 | 0.3592 |
| Milk production (kg) 3 | 8.29 | 8.49 | 8.07 | 8.26 | 9.53 | 0.1613 |
| Mortality in lactation (%) 4 | 5.13 ab | 2.83 b | 3.43 ab | 6.32 a | 6.77 | 0.0325 |
| Maternal Feed Restriction | Days of Pregnancy | p Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pregnancy | Control | R07 | R721 | R021 | 0 | 14 | 21 | 28 | ||||
| n = 5 | n = 5 | n = 5 | n = 5 | n = 10 | n = 20 | n = 20 | n = 20 | RMSE | pMFR | pTime | pMFR × Time | |
| T3 (pg/mL) | 3.76 a | 3.77 a | 3.19 ab | 3.06 b | 4.16 a | 3.07 b | 3.02 b | 3.54 ab | 0.79 | 0.0196 | 0.0026 | 0.0353 |
| T4 (pg/mL) | 4.74 | 4.24 | 5.00 | 5.16 | 5.89 ab | 3.66 b | 2.58 b | 7.00 a | 3.11 | 0.8525 | 0.0001 | 0.5731 |
| T3 to T4 ratio | 1.45 | 1.05 | 1.01 | 1.12 | 0.91 ab | 1.00 a | 2.06 a | 0.66 b | 1.24 | 0.8079 | 0.0022 | 0.5237 |
| Insulin (mU/L) | 15.73 a | 15.71 a | 9.47 b | 12.45 ab | 16.26 | 10.10 | 12.80 | 14.20 | 6.57 | 0.0225 | 0.0871 | 0.0055 |
| Glucose (mg/dL) | 79.94 | 78.00 | 74.49 | 84.13 | 63.82 b | 61.72 b | 76.64 b | 114.38 a | 22.34 | 0.6207 | 0.0001 | 0.7501 |
| Glucose-to-insulin ratio | 6.79 b | 7.12 b | 12.93 a | 9.17 ab | 4.59 | 9.56 | 10.26 | 11.70 | 6.79 | 0.0482 | 0.0802 | 0.2518 |
| HOMA-IR | 2.90 | 3.11 | 1.75 | 2.87 | 2.63 ab | 1.44 b | 2.40 b | 4.15 a | 1.79 | 0.1140 | 0.0001 | 0.0049 |
| NEFAS (mmol/L) | 0.42 ab | 0.33 b | 0.62 a | 0.52 ab | 0.54 | 0.54 | 0.46 | 0.35 | 0.27 | 0.0158 | 0.0781 | 0.0100 |
| Corticosterone (ng/mL) | 257.45 | 274.56 | 256.02 | 251.15 | 293.30 a | 363.82 b | 192.20 c | 195.85 c | 12.13 | 0.7344 | 0.0001 | 0.2519 |
| Maternal Feed Restriction | Days of Lactation | p Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lactation | Control | R07 | R721 | R021 | 7 | 14 1 | 30 2 | |||||
| n = 5 | n = 5 | n = 5 | n = 5 | n = 20 | n = 20 | (P) n = 20 | (NP) n = 20 | RMSE | pMFR | pTime | pMFR × Time | |
| T3 (pg/mL) | 3.47 | 3.41 | 3.48 | 3.40 | 3.70 a | 3.56 a | 3.54 a | 2.95 b | 0.66 | 0.9830 | 0.0056 | 0.0878 |
| T4 (pg/mL) | 10.10 | 12.83 | 11.26 | 11.38 | 7.84 | 12.45 | 13.18 | 12.10 | 5.76 | 0.6012 | 0.0753 | 0.5611 |
| T3 to T4 ratio | 0.57 | 0.36 | 0.40 | 0.72 | 0.74 | 0.67 | 0.33 | 0.31 | 0.58 | 0.2853 | 0.0847 | 0.2714 |
| Insulin (mU/L) | 16.36 | 12.09 | 13.36 | 14.23 | 21.48 a | 12.83 b | 10.05 b | 11.68 b | 7.41 | 0.4223 | 0.0006 | 0.2539 |
| Glucose (mg/dL) | 114.33 | 109.93 | 109.31 | 109.51 | 116.51 a | 111.28 ab | 112.09 ab | 103.20 b | 12.40 | 0.6352 | 0.0199 | 0.2822 |
| Glucose to Insulin ratio | 10.31 | 21.19 | 11.05 | 17.95 | 6.73 b | 15.13 b | 23.34 a | 15.30 b | 15.83 | 0.1555 | 0.0469 | 0.0802 |
| HOMA IR | 4.73 | 3.32 | 3.70 | 3.95 | 6.26 a | 3.58 b | 2.82 b | 3.05 b | 2.17 | 0.3178 | 0.0003 | 0.2801 |
| NEFAS (mmol/L) | 0.26 | 0.28 | 0.25 | 0.29 | 0.24 | 0.26 | 0.27 | 0.30 | 0.12 | 0.8113 | 0.4605 | 0.2305 |
| Corticosterone (ng/mL) | 239.11 | 234.36 | 215.89 | 246.54 | 151.11 a | 217.52 b | 272.84 c | 294.44 c | 11.51 | 0.4158 | 0.0001 | 0.2334 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández-Pacheco, C.; Millán, P.; Rodríguez, M.; Formoso-Rafferty, N.; Sánchez-Rodríguez, A.; Lorenzo, P.L.; Arias-Álvarez, M.; García-García, R.M.; Rebollar, P.G. Influence of Different Regimes of Moderate Maternal Feed Restriction during Pregnancy of Primiparous Rabbit Does on Long-Term Metabolic Energy Homeostasis, Productive Performance and Welfare. Animals 2021, 11, 2736. https://doi.org/10.3390/ani11092736
Fernández-Pacheco C, Millán P, Rodríguez M, Formoso-Rafferty N, Sánchez-Rodríguez A, Lorenzo PL, Arias-Álvarez M, García-García RM, Rebollar PG. Influence of Different Regimes of Moderate Maternal Feed Restriction during Pregnancy of Primiparous Rabbit Does on Long-Term Metabolic Energy Homeostasis, Productive Performance and Welfare. Animals. 2021; 11(9):2736. https://doi.org/10.3390/ani11092736
Chicago/Turabian StyleFernández-Pacheco, Carlota, Pilar Millán, María Rodríguez, Nora Formoso-Rafferty, Ana Sánchez-Rodríguez, Pedro L. Lorenzo, María Arias-Álvarez, Rosa M. García-García, and Pilar G. Rebollar. 2021. "Influence of Different Regimes of Moderate Maternal Feed Restriction during Pregnancy of Primiparous Rabbit Does on Long-Term Metabolic Energy Homeostasis, Productive Performance and Welfare" Animals 11, no. 9: 2736. https://doi.org/10.3390/ani11092736
APA StyleFernández-Pacheco, C., Millán, P., Rodríguez, M., Formoso-Rafferty, N., Sánchez-Rodríguez, A., Lorenzo, P. L., Arias-Álvarez, M., García-García, R. M., & Rebollar, P. G. (2021). Influence of Different Regimes of Moderate Maternal Feed Restriction during Pregnancy of Primiparous Rabbit Does on Long-Term Metabolic Energy Homeostasis, Productive Performance and Welfare. Animals, 11(9), 2736. https://doi.org/10.3390/ani11092736