
Hormonal and Neurological Aspects of Dog Walking for Dog Owners and Pet Dogs


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Participants
2.3. Experiments I and II
2.4. Measurements of Hormones
2.5. Measurements of Monoamines, Their Metabolites, and GABA
2.6. Statistics
3. Results
3.1. Effect on Dog Owners’ and Pet-Dogs’ Hormone Levels in Experiment I
3.2. Effects on Owners’ Hormone Levels and Neural Activity (MHPG and GABA Levels) in Experiment II
3.3. Results for Pet Dogs in Experiment II
4. Discussion
4.1. Dog Owners
4.2. Pet Dogs
4.3. A Relationship between a Pet Dog and the Owner
5. Conclusions
6. Limitations and Speculation
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, J.M. Stressful life events and use of physician services among the elderly: The moderating role of pet ownership. J. Pers. Soc. Psychol. 1990, 58, 1081–1086. [Google Scholar] [CrossRef]
- Serpell, J. Beneficial effects of pet ownership on some aspects of human health and behaviour. J. R. Soc. Med. 1991, 84, 717–720. [Google Scholar] [CrossRef] [PubMed]
- Friedmann, E.; Locker, B.Z.; Lockwood, R. Perception of animals and cardiovascular responses during verbalization with an animal present. Anthrozoös 1993, 6, 115–134. [Google Scholar] [CrossRef]
- Serpell, J.; Barrett, P. The Domestic Dog, 2nd ed.; Cambridge University Press: New York, NY, USA, 2017; pp. 5–66. [Google Scholar]
- Odendaal, J.S.J. Animal-assisted therapy—magic or medicine? J. Psychosom. Res. 2000, 49, 275–280. [Google Scholar] [CrossRef]
- Bowlby, J. The Bowlby-Ainsworth attachment theory. Behav. Brain Sci. 1979, 2, 637–638. [Google Scholar] [CrossRef]
- Odendaal, J.S.; Meintjes, R. Neurophysiological correlates of affiliative behaviour between humans and dogs. Vet. J. 2003, 165, 296–301. [Google Scholar] [CrossRef]
- Nagasawa, M.; Kikusui, T.; Onaka, T.; Ohta, M. Dog’s gaze at its owner increases owner’s urinary oxytocin during social interaction. Horm. Behav. 2009, 55, 434–441. [Google Scholar] [CrossRef]
- Nagasawa, M.; Mitsui, S.; En, S.; Ohtani, N.; Ohta, M.; Sakuma, Y.; Onaka, T.; Mogi, K.; Kikusui, T. Oxytocin-gaze positive loop and the coevolution of human-dog bonds. Science 2015, 348, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.A.; Beck, A.M.; McCune, S. The Health Benefits of Dog Walking for Pets and People: Evidence and Case Studies; Purdue University Press: West Lafayette, IN, USA, 2011. [Google Scholar]
- Christian, H.; Bauman, A.; Epping, J.N.; Levine, G.N.; McCormack, G.; Rhodes, R.E.; Richards, E.; Rock, M.; Westgarth, C. Encouraging Dog walking for health promotion and disease prevention. Am. J. Lifestyle Med. 2018, 12, 233–243. [Google Scholar] [CrossRef]
- Powell, L.; Edwards, K.M.; Bauman, A.; Guastella, A.J.; Drayton, B.; Stamatakis, E.; McGreevy, P. Canine endogenous oxytocin responses to dog-walking and affiliative human–dog interactions. Animals 2019, 9, 51. [Google Scholar] [CrossRef]
- Powell, L.; Edwards, K.M.; Michael, S.; McGreevy, P.; Bauman, A.; Guastella, A.J.; Drayton, B.; Stamatakis, E. Effects of human–dog interactions on salivary oxytocin concentrations and heart rate variability: A four-condition cross-over trial. Anthrozoös 2020, 33, 37–52. [Google Scholar] [CrossRef]
- Motooka, M.; Koike, H.; Yokoyama, T.; Kennedy, N.L. Effect of dog-walking on autonomic nervous activity in senior citizens. Med. J. Aust. 2006, 184, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Krause-Parello, C.A.; Friedmann, E.; Blanchard, K.; Payton, M.; Gee, N.R. Veterans and shelter dogs: Examining the impact of a dog-walking intervention on physiological and post-traumatic stress symptoms. Anthrozoös 2020, 33, 225–241. [Google Scholar] [CrossRef]
- Fujimaru, C.; Okamura, H.; Kawasaki, M.; Kakuma, T.; Yoshii, C.; Matsuishi, T. Self-perceived work-related stress and its relation to salivary IgA, cortisol and 3-methoxy-4-hydroxyphenyl glycol levels among neonatal intensive care nurses. Stress Health 2012, 28, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Apfel, B.A.; Otte, C.; Inslicht, S.S.; McCaslin, S.E.; Clare Henn-Haase, P.D.; Metzler, T.J.; Makotkine, I.; Yehuda, R.; Neylan, T.C.; Marmar, C.R. Pretraumatic prolonged elevation of salivary MHPG predicts peritraumatic distress and symptoms of post-traumatic stress disorder. J. Psychiatr. Res. 2011, 45, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Post, R.M.; Gordon, E.K.; Goodwin, F.K.; Bunney, W.E. Central norepinephrine metabolism in affective Illness: MHPG in the cerebrospinal fluid. Science 1973, 179, 1002–1003. [Google Scholar] [CrossRef]
- Bowden, C.; Cheetham, S.C.; Lowther, S.; Katona, C.L.; Crompton, M.R.; Horton, R.W. Reduced dopamine turnover in the basal ganglia of depressed suicides. Brain Res. 1997, 769, 135–140. [Google Scholar] [CrossRef]
- Krnjevic, K. Role of GABA in cerebral cortex. Can. J. Physiol. Pharmacol. 1997, 75, 439–451. [Google Scholar] [CrossRef] [PubMed]
- Harvie, H.; Rodrigo, A.; Briggs, C.; Thiessen, S.; Kelly, D.M. Does stress run through the leash? An examination of stress transmission between owners and dogs during a walk. Anim. Cogn. 2021, 24, 239–250. [Google Scholar] [CrossRef]
- Arai, K.; Watanabe, G.; Fujimoto, M.; Nagata, S.; Takemura, Y.; Taya, K.; Sasamoto, S. A sensitive radioimmunoassay for cortisol using 125I-labeled radioligand. J. Reprod. Dev. 1995, 41, 15–20. [Google Scholar] [CrossRef]
- Daughters, K.; Manstead, A.S.R.; Hubble, K.; Rees, A.; Thapar, A.; van Goozen, S.H.M. Salivary oxytocin concentrations in males following intranasal administration of oxytocin: A double-blind, cross-over study. PLoS ONE 2015, 10, e0145104. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.E.; Sergiel, A.; Selva, N.; Swenson, J.E.; Zedrosser, A.; Stenhouse, G.; Janz, D.M. Correcting for enzyme immunoassay changes in long term monitoring studies. MethodsX 2021, 8, 101212. [Google Scholar] [CrossRef]
- Beetz, A.; Uvnäs-Moberg, K.; Julius, H.; Kotrschal, K. Psychosocial and psychophysiological effects of human-animal interactions: The possible role of oxytocin. Front Psychol. 2012, 3, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Li, G.Y.; Watanabe, I.; Kunitake, Y.; Sugataka, K.; Muraoka, T.; Kojima, N.; Kawashima, T.; Yamada, S. Relationship between saliva level of 3-methoxy-4-hydroxyphenylglycol and mental health in the elderly general population. Psychiatry Clin. Neurosci. 2008, 62, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Sheline, Y.I.; Miller, K.; Bardgett, M.E.; Csernansky, J.G. Higher cerebrospinal fluid MHPG in subjects with dementia of the Alzheimer type: Relationship with cognitive dysfunction. Am. J. Geriatr. Psychiatry 1998, 6, 155–161. [Google Scholar] [CrossRef]
- Sugaya, N.; Izawa, S.; Yamamoto, R.; Ogawa, N.; Yajima, J.; Okamura, H.; Horiuchi, S.; Tsuda, A.; Nomura, S. Salivary 3-methoxy-4-hydroxyphenylglycol increases after awakening in healthy young adults. Psychophysiology 2015, 52, 425–428. [Google Scholar] [CrossRef]
- Reuster, T.; Buechler, J.; Winiecki, P.; Oehler, J. Influence of reboxetine on salivary MHPG concentration and cognitive symptoms among patients with alcohol-related Korsakoff’s syndrome. Neuropsychopharmacology 2003, 28, 974–978. [Google Scholar] [CrossRef][Green Version]
- Giacomello, G.; Scholten, A.; Parr, M.K. Current methods for stress marker detection in saliva. J. Pharm. Biomed. Anal. 2020, 191, 113604. [Google Scholar] [CrossRef]
- Buchman, A.S.; Yu, L.; Wilson, R.S.; Boyle, P.A.; Schneider, J.A.; Bennett, D.A. Brain pathology contributes to simultaneous change in physical frailty and cognition in old age. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2014, 69, 1536–1544. [Google Scholar] [CrossRef]
- Ma, L.; Chan, P. Understanding the physiological links between physical frailty and cognitive decline. Aging Dis. 2019, 11, 405–418. [Google Scholar] [CrossRef]
- Steenbergen, L.; Sellaro, R.; Stock, A.K.; Beste, C.; Colzato, L.S. γ-Aminobutyric Acid (GABA) administration improves action selection processes: A randomised controlled trial. Sci. Rep. 2015, 5, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Esch, T.; Stefano, G.B. The neurobiology of love. Neuroendocrinol. Lett. 2005, 26, 175–192. [Google Scholar] [PubMed]
- Ogi, A.; Mariti, C.; Baragli, P.; Sergi, V.; Gazzano, A. Effects of stroking on salivary oxytocin and cortisol in guide dogs: Preliminary results. Animals 2020, 10, 708. [Google Scholar] [CrossRef] [PubMed]
- López-Arjona, M.; Mateo, S.V.; Cerón, J.J.; Martínez-Subiela, S. Changes in salivary oxytocin after stroking in dogs: Validation of two assays for its assessment. Res. Vet. Sci. 2021, 136, 527–534. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Owner | Dog | Owner | Dog | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Sex | Age | Breeds | Age | Sex | Age | Breeds | Age | ||
| 1 * | M | 71 | Labrador Retriever | 11 | 18 * | F | 66 | Miniature Dachshund | 4 |
| 2 | F | 65 | Golden Retriever | 2 | 19 | F | 70 | Golden Retriever | 2 |
| 3 | M | 71 | Shiba | 12 | 20 | M | 55 | White Shepherd | 4 |
| 4 | F | 58 | Golden Retriever | 10 | 21 | F | 50 | White Shepherd | 4 |
| 5 | F | 59 | Shiba | 7 | 22 | F | 45 | Shiba | 5 |
| 6 | M | 75 | Golden Retriever | 3 | 23 | M | 50 | Shiba | 5 |
| 7 * | F | 70 | Miniature Dachshund | 5 | 24 | M | 50 | White Shepherd | 4 |
| 8 * | F | 66 | Mix | 10 | 25 | F | 50 | Toy Poodle | 7 |
| 9 | M | 63 | Labrador Retriever | 3 | 26 * | F | 50 | Miniature Dachshund | 7 |
| 10 | F | 61 | Labrador Retriever | 12 | 27 | F | 50 | Mix | 5 |
| 11 | F | 60 | Labrador Retriever | 9 | 28 | M | 55 | Mix | 5 |
| 12 | F | 65 | Golden Retriever | 1 | 29 | F | 65 | Labrador Retriever | 8 |
| 13 * | F | 60 | Toy Poodle | 2 | 30 | F | 67 | Labrador Retriever | 9 |
| 14 * | F | 53 | Miniature Dachshund | 2 | 31 | F | 68 | Miniature Schnauzer | 7 |
| 15 | F | 65 | Shiba | 1 | 32 | F | 50 | Shiba | 5 |
| 16 * | F | 60 | Toy Poodle | 8 | 33 * | F | 53 | Toy Poodle | 7 |
| 17 | F | 65 | Shiba | 7 | 34 * | M | 75 | Miniature Schnauzer | 2 |
| Oxytocin | Cortisol | |||||
|---|---|---|---|---|---|---|
| Stages | N | Means | S.E. | Means | S.E. | |
| Walk without dog | Pre(A) | 10 | 59.24 | 23.54 | 0.38 | 0.06 |
| Walk1(A) | 10 | 82.38 | 20.13 | 0.33 | 0.05 | |
| Walk2(A) | 10 | 92.44 | 27.68 | 0.28 | 0.04 | |
| Post(A) | 10 | 92.27 | 34.20 | 0.25 | 0.03 | |
| Walk with dog | Pre(D) | 10 | 57.96 | 11.45 | 0.50 | 0.11 |
| Walk1(D) | 10 | 117.11 | 28.54 | 0.38 | 0.08 | |
| Walk2(D) | 10 | 85.99 | 20.10 | 0.41 | 0.10 | |
| Post(D) | 10 | 77.22 | 17.88 | 0.34 | 0.06 | |
| Dog | Pre(D) | 10 | 275.0 | 25.1 | 0.16 | 0.01 |
| Walk1(D) | 10 | 329.3 | 26.9 | 0.20 | 0.02 | |
| Walk2(D) | 10 | 312.9 | 21.8 | 0.20 | 0.03 | |
| Post(D) | 10 | 325.3 | 27.8 | 0.17 | 0.02 | |
| A. Walking without Dog | |||||
| Substances | Units | N | Means | S.E. | |
| Pre(A) | Oxytocin | pg/mL | 14 | 62.30 | 9.14 |
| Cortisol | μg/dL | 14 | 0.38 | 0.06 | |
| MHPG | ng/mL | 14 | 264.10 | 134.31 | |
| GABA | μM | 14 | 1.15 | 0.13 | |
| Walk1(A) | Oxytocin | pg/mL | 14 | 93.40 | 19.47 |
| Cortisol | μg/dL | 14 | 0.33 | 0.05 | |
| MHPG | ng/mL | 14 | 274.59 | 142.76 | |
| GABA | μM | 14 | 0.98 | 0.10 | |
| Walk2(A) | Oxytocin | pg/mL | 14 | 83.18 | 16.56 |
| Cortisol | μg/dL | 14 | 0.28 | 0.04 | |
| MHPG | ng/mL | 14 | 294.59 | 162.75 | |
| GABA | μM | 14 | 0.84 | 0.14 | |
| Post(A) | Oxytocin | pg/mL | 14 | 60.96 | 15.39 |
| Cortisol | μg/dL | 14 | 0.25 | 0.03 | |
| MHPG | ng/mL | 14 | 285.11 | 144.83 | |
| GABA | μM | 14 | 0.62 | 0.12 | |
| B. Walking with Dog | |||||
| Substances | Units | N | Means | S.E. | |
| Pre(D) | Oxytocin | pg/mL | 14 | 60.96 | 14.23 |
| Cortisol | μg/dL | 14 | 0.50 | 0.11 | |
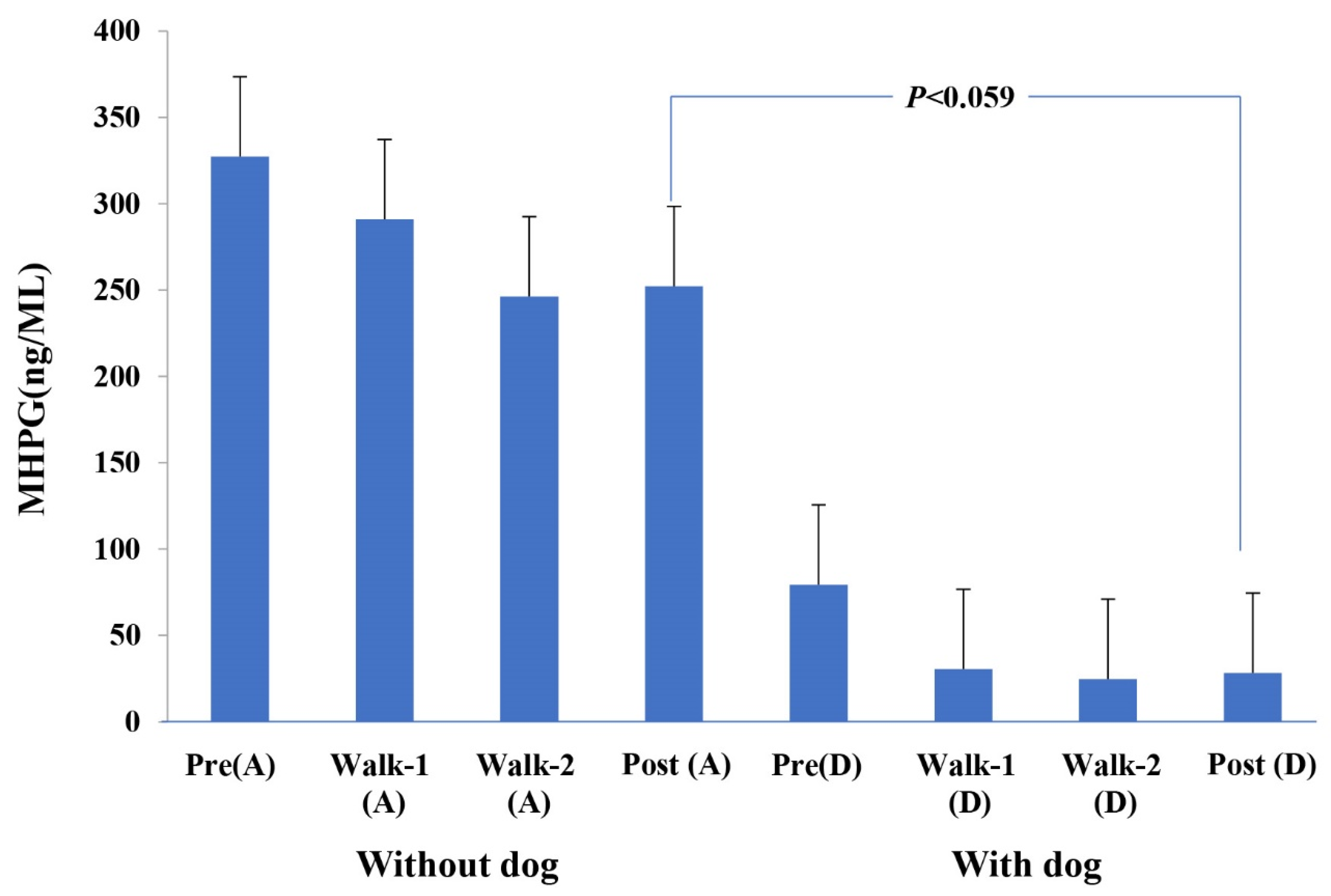
| MHPG | ng/mL | 14 | 79.36 | 44.37 | |
| GABA | μM | 14 | 0.91 | 0.13 | |
| Walk1(D) | Oxytocin | pg/mL | 14 | 64.39 | 12.41 |
| Cortisol | μg/dL | 14 | 0.38 | 0.08 | |
| MHPG | ng/mL | 14 | 30.55 | 8.12 | |
| GABA | μM | 14 | 1.21 | 0.10 | |
| Walk2(D) | Oxytocin | pg/mL | 14 | 68.35 | 12.50 |
| Cortisol | μg/dL | 14 | 0.41 | 0.10 | |
| MHPG | ng/mL | 14 | 28.51 | 6.70 | |
| GABA | μM | 14 | 1.36 | 0.13 | |
| Post(D) | Oxytocin | pg/mL | 14 | 67.06 | 15.39 |
| Cortisol | μg/dL | 14 | 0.34 | 0.06 | |
| MHPG | ng/mL | 14 | 28.35 | 7.63 | |
| GABA | μM | 14 | 1.27 | 0.14 | |
| Oxytocin | Cortisol | |||||
|---|---|---|---|---|---|---|
| Stages | N | Means | S.E. | Means | S.E. | |
| Dog | Pre(D) | 14 | 284.5 | 19.0 | 0.17 | 0.03 |
| Walk1(D) | 14 | 332.7 | 20.9 | 0.16 | 0.02 | |
| Walk2(D) | 14 | 345.0 | 25.2 | 0.14 | 0.01 | |
| Post(D) | 14 | 312.2 | 28.9 | 0.21 | 0.05 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akiyama, J.; Ohta, M. Hormonal and Neurological Aspects of Dog Walking for Dog Owners and Pet Dogs. Animals 2021, 11, 2732. https://doi.org/10.3390/ani11092732
Akiyama J, Ohta M. Hormonal and Neurological Aspects of Dog Walking for Dog Owners and Pet Dogs. Animals. 2021; 11(9):2732. https://doi.org/10.3390/ani11092732
Chicago/Turabian StyleAkiyama, Junko, and Mitsuaki Ohta. 2021. "Hormonal and Neurological Aspects of Dog Walking for Dog Owners and Pet Dogs" Animals 11, no. 9: 2732. https://doi.org/10.3390/ani11092732
APA StyleAkiyama, J., & Ohta, M. (2021). Hormonal and Neurological Aspects of Dog Walking for Dog Owners and Pet Dogs. Animals, 11(9), 2732. https://doi.org/10.3390/ani11092732