1. Introduction
Auricularia mushrooms, a member of the basidiomycetes, also known as wood ears, are a group of edible fungi that form a gelatinous fruiting body [
1]. It is well documented that
Auricularia are widely distributed in temperate, subtropical, and tropical regions, particularly in many northern temperate-zone countries, including China, Japan, Korea, and Thailand [
2,
3]. As one of the traditional Chinese edible and medicinal fungi,
Auricularia has been confirmed to possess various biological activities [
4].
It has been generally considered that
Auricularia polysaccharides (AP) are the major bioactive substance. Various bioactivities of AP have been reported, including immunomodulatory, antioxidant, antitumor activities, hepatoprotective [
5]. Wu et al. (2010) [
6] found that AP could modulate the immune functions of aged mice by increasing the weights of the thymus and spleen. In the opinion of Miao et al. (2020) [
5], AP could activate the innate immune system and effector cells, including T-lymphocytes, B-lymphocytes, macrophages, and natural killer cells to express cytokines, such as IL-1β, TNF-α, IL-6, and IL-10. Five water-soluble APs from different varieties were all observed to have scavenging activities against superoxide, 2,2-diphenyl-1-picrylhydrazyl (DPPH), and hydroxyl radicals [
7]. A soluble AP was observed to down-regulate the serum levels of blood lipids in dietary-induced hypercholesterolemic rats to normal [
8]. According to Miao et al. (2020) [
5], the AP could inhibit the absorption of exogenous lipids by binding with cholate or lipid molecules in gastrointestinal tracts and promote total cholesterol (TC) metabolism. The hypolipidemic activities were determined by molecular weight, monosaccharide compositions, uronic acid concentrations, and chain conformations of AP. Wang et al. (2018) [
9] measured the hepatoprotective effects of AP in the acute alcohol-induced alcoholic liver diseases (ALD), and AP showed increasing antioxidant activities, reducing lipid peroxidation, inhibiting the expression levels of inflammatory mediators, and preventing the alcohol-induced histopathological alterations. The biological activities may be attributed to the abundant glucose and xylose concentrations in AP.
Selenium is an essential nonmetallic micronutrient for all animals, plays vital roles in several physiological processes, and has potent anti-inflammatory, antioxidant, and antiviral effects [
10]. The bioavailability, as well as the toxicological effect of Se, was associated with chemical forms. Organic Se possessed a potent antioxidant capacity that provided greater protection against oxidative damage and less toxicity than inorganic Se [
11]. Several studies indicated that dietary Se supplementation could improve the meat quality of chicken [
12,
13]. Chen et al. (2019) [
14] showed that Se supplementation could enhance the antioxidant status and meat quality of growing-finishing pigs.
To produce Selenium
Auricularia cornea culture (SAC), a specific culture media containing sodium selenite was inoculated with the
Auricularia cornea (AC) strain and processed via solid fermentation under humidity–temperature controlled conditions for 15 days. Subsequently, the fermented culture media was dried at 60 °C. AS AC could utilize the carbohydrates and proteins present in media via enzymolysis, they result in a variety of metabolic products, such as organic selenium, peptides, amino acids, oligosaccharides, and polysaccharides. The analyzed nutrient content of SAC is shown in
Table 1.
Because of its abundant nutraceutical compounds, we hypothesized that SAC might be an effective feed additive that could improve the growth and antioxidant status of pigs. There was also a strong correlation between oxidative stress and meat quality [
15]. Therefore, the purpose of the current study was to determine the digestible energy (DE), metabolizable energy (ME) contents of SAC and to evaluate the effects of SAC supplementation on growth performance, short chain fatty acid (SCFA) contents in feces, serum biochemical parameters, carcass characteristics, meat quality, and oxidative stability of fresh meat in growing-finishing pigs.
3. Results
The DE and metabolizable energy (ME) values of SAC were 11.21 and 10.92 MJ/kg (as-fed basis), respectively (
Table 5).
As shown in
Table 6, pigs fed a 2.4% SAC diet had a higher final body weight (FBW) (
p < 0.05) than the other three diets. Compared with the control diet, pigs fed diets supplemented with SAC showed a decreased F/G (
p < 0.05) value during day 1 to 23. From day 24 to 45, 1.2% and 2.4% SAC supplemented in diets enhanced ADG and reduced F/G, compared with the control and 0.6% SAC diets (
p < 0.01). During the overall period, pigs in 1.2% and 2.4% SAC treatments had the higher ADG (
p < 0.05), and 0.6%, 1.2%, and 2.4% SAC treatments had the lower F/G than that in the control treatment (
p < 0.05).
Compared with the control diet, dietary SAC supplementation significantly increased butyrate concentration (
p < 0.05) on day 45 (
Table 7).
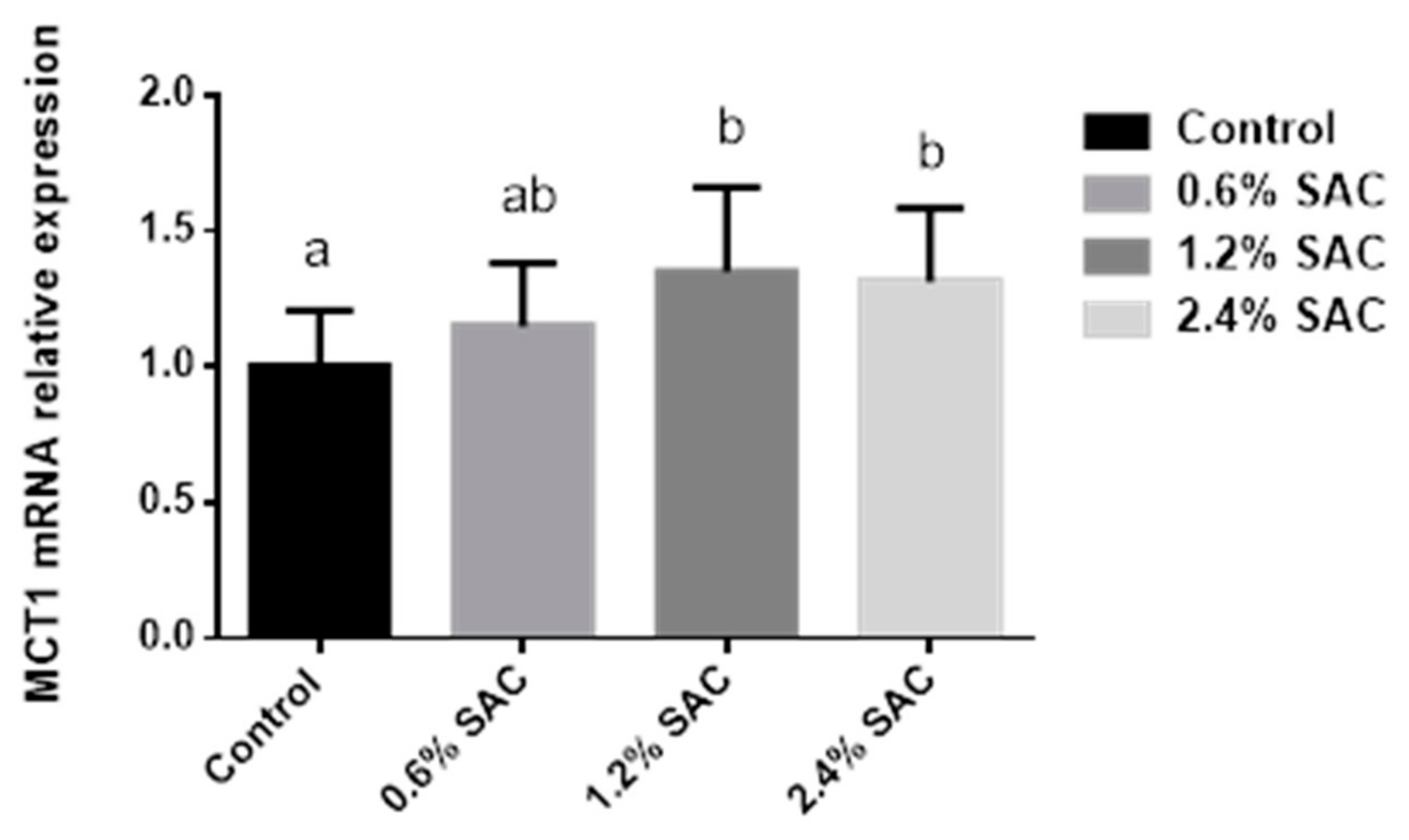
As shown in
Figure 1, pigs fed 1.2% and 2.4% SAC diets showed a higher MCT1 mRNA expression (
p = 0.04) in the colon than in the control diet.
As illustrated in
Table 8, the GSH-Px contents (
p < 0.05) in 1.2% and 2.4% SAC groups were remarkably higher than that in the control group on day 45. However, no differences in T-AOC, SOD, MDA, and the other serum indices were found in finishing pigs fed different levels of SAC.
As revealed in
Table 9, dietary supplementation of 2.4% SAC significantly improved GSH-Px content (
p < 0.05), and SAC supplemented at 1.2% and 2.4% markedly reduced MDA contents (
p < 0.05) in the liver. The GSH-Px activity (
p < 0.05) in 2.4% treatment was significantly higher in LDM than in the control treatment.
Finishing pigs fed 2.4% SAC showed a higher Se content (
p < 0.05) in LDM compared with the other three treatments (
Table 10). There was an increasing trend in liver Se content for pigs fed SAC diets (
p = 0.09).
As shown in
Table 11, no effects on carcass traits and meat quality were observed in pigs fed different levels of SAC.
As displayed in
Table 12, compared with the control diet, pigs fed a 2.4% SAC diet had lower MDA content (
p < 0.05) in fresh meat during the simulated retail display on day 6.
4. Discussion
The ME value is defined as the DE minus GE in urine and fermentation gases. The ME represents approximately 92–98% of the proportion of DE. Gas energy varies and is typically small for conventional diets fed to pigs (about 0.5% DE for growing-finishing pigs). It could be as high as 3% of DE in sows fed high-fiber diets. The GE value in urine is the primary factor defining the proportion of DE converted to ME [
19]. According to the procedure by Adeola [
23], we performed this study without evaluating gas losses on ME measurement. The ME content in SAC should be accurately determined via gas loss assessment in future study. The SAC’s NDF (24.09%) and ADF (8.98%) contents were comparable with those of wheat bran (NDF:32.28%, ADF:11.00%), rice bran (NDF:26.28%, ADF:11.87%), and corn bran (NDF:32.96%, ADF:9.23%). The DE (11.21 MJ/kg) and ME (10.92 MJ/kg) contents in SAC were analogous to wheat bran (DE:10.13 MJ/kg, ME:9.70 MJ/kg), rice bran (DE:12.97 MJ/kg, ME:12.54 MJ/kg), and corn bran (DE:11.08 MJ/kg, ME:10.81 MJ/kg), indicating that SAC could be a fibrous ingredient [
19].
In the current study, we observed that pigs fed SAC diets could improve FBW, ADG and reduce F/G. Many studies showed that dietary Se levels and sources did not affect growth performance [
14,
33,
34]. The most likely explanation was that the nutraceutical compounds in SAC, especially the polysaccharide (β-glucan), played a vital role in the growth improvement. The mushrooms’ β-glucan contents ranged from 0.21% to 0.53% (dry weight basis) [
35]. As Luo et al. (2019) [
36] indicated, β-glucan was a sort of functional polysaccharide that widely existed in the cell wall of fungi and possessed many biological activities. Previous studies showed that pigs fed β-glucan diets might enhance growth performance, which was probably due to the intestinal improvement, as β-glucan could modulate intestinal structure and morphology [
30,
37,
38].
The European Food and Safety Authority (EFSA) suggested that the supplementation level of Se-yeast should not exceed a maximum of 0.2 mg/kg in a complete diet [
39]. Therefore, the up-level of Se supplementation in the diet was 0.13 mg/kg in the current study. Most of the researches revealed that growth performance was unaffected by Se levels (0.1, 0.3, and 0.5 mg/kg) and sources (Se-yeast or sodium selenite) [
40,
41]. A study showed a decreased ADFI of growing-finishing pigs fed 0.5 mg/kg sodium selenite [
42]. The discrepancy could be attributed to an adverse effect because inorganic Se is more susceptible to selenosis compared to organic Se [
14].
In the present study, pigs fed SAC showed a higher butyrate content compared with the control diet. We speculated that the polysaccharide in SAC might play a central role in increasing SCFA. Polysaccharides could be an effective substrate for the SCFA-generated bacteria and modify the relative composition of microbiota in the intestine. Feed ingredients that stimulate a higher SCFA production, particularly butyrate, were generally considered beneficial [
43]. This study was in agreement with previous results, as Högberg et al. (2006) [
44] revealed that diets supplemented with high-level insoluble non-starch polysaccharides (NSP) (188–250 g/kg DM) could promote butyrate content in the hindgut. Metzler-Zebeli et al. (2011) [
45] also observed that β-glucan increased caecal and colonic butyrate concentrations. The content of NSP and β-glucan in SAC was not detected in this study, and the concentration of crude fiber, NDF, and ADF in SAC was 6.43%, 24.09%, and 8.98%, respectively. A further quantitative study is necessary to explore the mechanism of improving butyrate content by SAC addition.
Butyrate is the result of microbial fermentation in the hindgut of animals. It serves not only as the primary energy source for colonocytes but also as a cellular mediator for gut cells, such as gut tissue development, immune modulation, gene expression, diarrhea control, and oxidative stress reduction [
46]. The butyrate transportation was mediated by the monocarboxylate transporter 1 (MCT1). To ascertain the mechanisms, we analyzed the colonic MCT1 mRNA expression, as Cuff et al. (2020) [
47] indicated that the MCT1 pathway was positively relevant to the butyrate generation, and butyrate showed a time and concentration-dependent relationship of both MCT1 mRNA and protein. The result showed a higher MCT1 mRNA expression in pigs fed SAC diets, consistent with a previous study indicating that dietary β-glucan supplementation could upregulate MCT1 expression in the cecum [
30].
We observed a significant increase in GSH-Px content in serum, LDM, and liver of pigs fed SAC diets. The result might be attributed to the Se present in SAC. Pilarczyk et al. (2012) [
48] revealed that Se is an integral part of GSH-Px. It works through the reduction of hydrogen peroxide that protects subcellular and cellular membranes from oxidative damage. This result was in accordance with a previous study where pigs showed a significantly higher serum GSH-Px activity as dietary Se level increased, compared with the basal diet [
49]. Pigs fed higher SAC level diets revealed a decreased MDA in the liver. Malondialdehyde, one of the most studied lipid peroxidation products, was an important marker for oxidative stress. The MDA concentration is well correlated with the GSH-Px activity, and GSH-Px protects the tissue from peroxidation to keep MDA content in a low status [
50]. Zhan et al. (2007) [
50] showed that dietary Se supplementation significantly reduced the MDA content in the liver, which was consistent with our findings.
In the current study, the LDM Se contents were improved remarkably in response to the dietary 2.4% SAC supplementation. The reason might be because muscle Se content had a high correlation to dietary Se content. A previous study demonstrated that diets with a high Se level had a greater loin Se content [
51]. This result was consistent with previous findings, which reported a significant improving Se retention in both the muscles and glands of growing pigs fed Se diets [
52,
53]. Researchers confirmed that organic Se was more greatly retained by tissue (muscle and liver) than inorganic Se [
40,
49,
52]. We have not saturated the mechanism for Se deposition in the growing pigs fed SAC diets, and we observed a lower Se content in muscle than in the other tissue. The possible reason might be the lower rate of muscle accretion, which allowed more Se to be available to be distributed to other non-muscle tissues [
40,
53].
In this study, SAC-supplemented diets did not affect the meat quality of finishing pigs. The possible reason might be the relatively low Se content (upper limit 0.13 mg/kg) in experimental diets. In the study of Chen et al. (2019) [
14], a higher level of dietary Se supplementation (0.5 mg/kg) tended to reduce the drip loss of loins. Similarly, Calvo et al. (2016) [
54] reported that a dose of dietary 0.4 mg/kg Se (Se-enriched yeast) addition could increase water-holding capacity. Meat color is one of the leading factors affecting meat purchasing. In the current study, no effect was observed on meat color for pigs fed SAC diets. A relatively low Se level in SAC diets might lead to the result. A supplemented level of 0.5 mg/kg Se could improve a* value of meat in finishing pigs [
14]. Calvo et al. (2017) [
55] also reported that dietary 0.3 mg/kg Se addition increased the a* value of muscle in growing-finishing pigs. In these studies, the meat color improvement might be attributed to the enhanced antioxidant activities induced by dietary Se supplementation. As Mancini et al. (2005) [
56] indicated, Metmyoglobin reduction is a vital factor for meat color life, and remarkably lies on oxygen scavenging enzymes, reducing enzyme systems, and the nicotinamide adenine dinucleotide (NADH) pool.
Lipid peroxidation is a leading cause of meat quality deterioration during storage. In the current study, SAC supplementation reduced MDA content in fresh meat on day 6. Malondialdehyde was generated by lipid oxidation reaction induced by oxygen radicals in tissues and was one of the most crucial metabolites of lipid peroxides [
50,
57]. The MDA content in meat has been used as an important marker of lipid oxidation intensity [
58]. The decreased MDA concentration indicated that Se contained in SAC could enhance the ability to protect against oxidation, delay the onset of rancidity resulting from lipid oxidation, and extend the shelf life of meat to some extent, which could also avoid reducing consumer acceptability. The Se protective action against the deterioration of meat quality during storage was largely attributed to its effects of maintaining muscle membrane integrity and preventing lipid peroxidation [
50,
57]. Calvo et al. (2017) [
59] reported that pigs fed organic Se showed higher stability against oxidation in muscles, which was in accord with our result. Pappas et al. (2012) [
57] also found that chickens fed Se diets showed a linear decrease in lipid oxidation of breast muscle, which was probably due to the Se antioxidant properties.
One of the main challenges facing the meat industry is to add value to the final product. The novel Se source might benefit the meat industry because of health improvements and increasing meat oxidative stability. Meat is a major source of nutrition, such as protein, selenium, and vitamins. These nutrients are linked to the maintenance of health status. Research on selenium (particularly organic Se) is increasing rapidly concerning immune response and cancer prevention in humans. It is essential for the meat industry that meat products are delivered to the market not only in terms of consumer preference but also in terms of health benefits. The SAC is mainly composed of fungal polysaccharides, organic Se, and metabolites of AC. Currently, dietary SAC addition increased Se’s content in meat and benefited pigs’ health. In the future, quantification research of SAC is essential for achieving its desired potential benefits in the commercial production and meat industry.


 ,
, 

{kind=link}