
Is the Prevalence of Leishmania infantum Linked to Breeds in Dogs? Characterization of Seropositive Dogs in Ibiza




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Data and Collection of Samples
2.3. Serologic Tests
2.4. Statistical Analysis
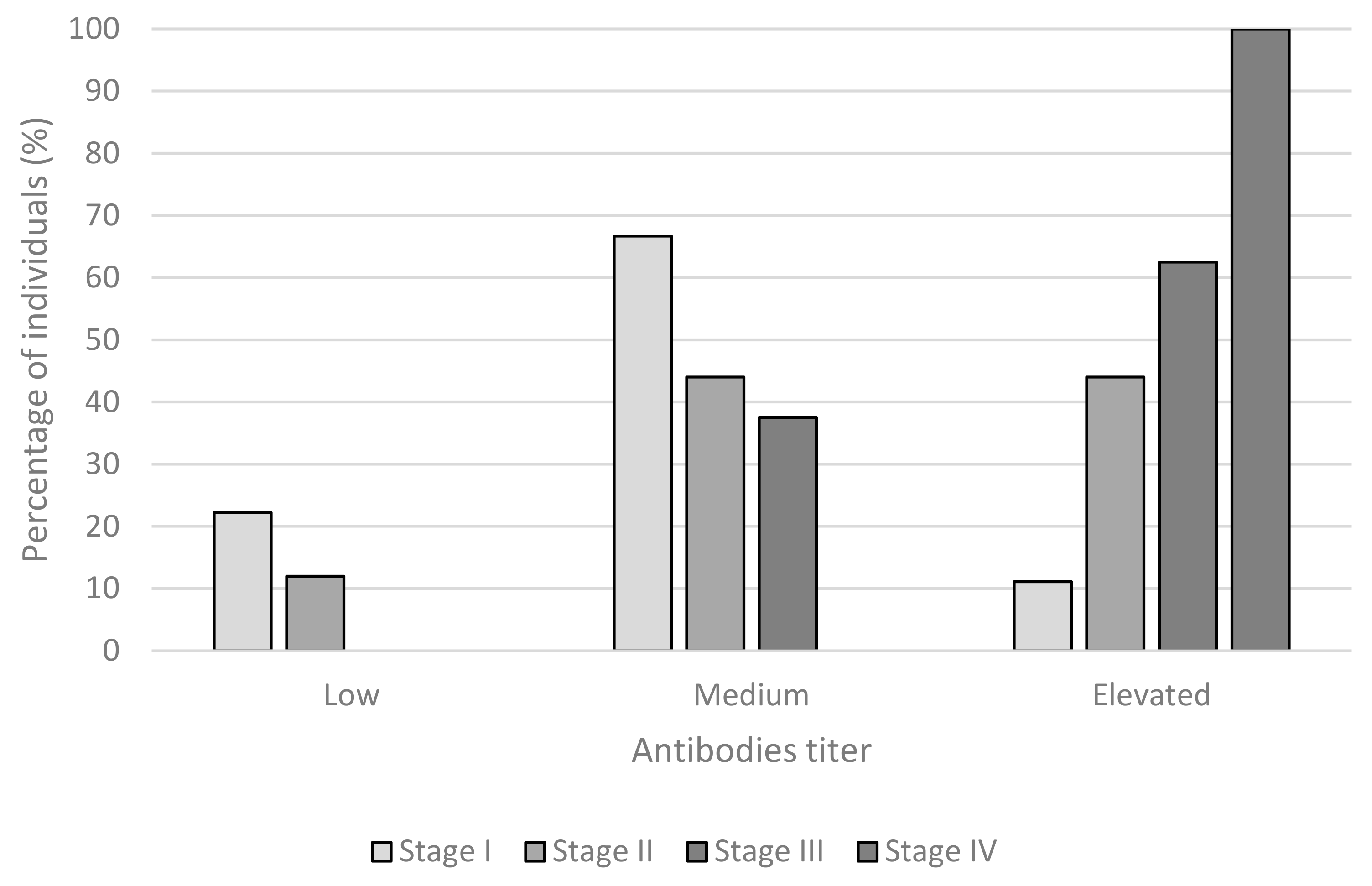
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohebali, M.; Malmasi, A.; Khodabakhsh, M.; Zarei, Z.; Akhoundi, B.; Hajjaran, H.; Azarm, A. Feline leishmaniosis due to Leishmania infantum in Northwest Iran: The role of cats in endemic areas of visceral leishmaniosis. Vet. Parasitol. Reg. Stud. Rep. 2017, 9, 13–16. [Google Scholar] [CrossRef]
- Solano-Gallego, L.; Miró, G.; Koutinas, A.; Cardoso, L.; Pennisi, M.G.; Ferrer, L.; Bourdeau, P.; Oliva, G.; Baneth, G. LeishVet Guidelines for the practical management of canine leishmaniosis. Parasites Vectors 2011, 4, 86. [Google Scholar] [CrossRef] [Green Version]
- Solano-Gallego, L.; Morell, P.; Arboix, M.; Alberola, J.; Ferrer, L. Prevalence of leishmania infantum infection in dogs living in an area of canine leishmaniasis endemicity using PCR on several tissues and serology. J. Clin. Microbiol. 2001, 39, 560–563. [Google Scholar] [CrossRef] [Green Version]
- Leontides, L.S.; Saridomichelakis, M.N.; Billinis, C.; Kontos, V.; Koutinas, A.F.; Galatos, A.D.; Mylonakis, M.E. A Cross-sectional study of Leishmania Spp. infection in clinically healthy dogs with polymerase chain reaction and serology in Greece. Vet. Parasitol. 2002, 109, 19–27. [Google Scholar] [CrossRef]
- Berrahal, F.; Mary, C.; Roze, M.; Berenger, A.; Escoffier, K.; Lamouroux, D.; Dunan, S. Canine leishmaniasis: Identification of asymptomatic carriers by polymerase chain reaction and immunoblotting. Am. J. Trop. Med. Hyg. 1996, 55, 273–277. [Google Scholar] [CrossRef]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votýpka, J.; Marty, P.; Delaunay, P.; Sereno, D. A Historical overview of the classification, evolution, and dispersion of leishmania parasites and sandflies. PLoS Negl. Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef] [PubMed]
- Mhadhbi, M.; Sassi, A. Infection of the Equine population by leishmania parasites. Equine Vet. J. 2020, 52, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Asfaram, S.; Fakhar, M.; Teshnizi, S.H. Is the cat an important reservoir host for visceral leishmaniasis? A systematic review with meta-analysis. J. Venom. Anim. Toxins Incl. Trop. Dis. 2019, 25, e20190012. [Google Scholar] [CrossRef]
- González, E.; Molina, R.; Iriso, A.; Ruiz, S.; Aldea, I.; Tello, A.; Fernández, D.; Jiménez, M. Opportunistic feeding behaviour and leishmania infantum detection in phlebotomus perniciosus females collected in the human leishmaniasis focus of Madrid, Spain (2012–2018). PLoS Negl. Trop. Dis. 2021, 15, e0009240. [Google Scholar] [CrossRef] [PubMed]
- Moraes-Silva, E.; Antunes, F.R.; Rodrigues, M.S.; da Silva Julião, F.; Dias-Lima, A.G.; Lemos-de-Sousa, V.; de Alcantara, A.C.; Reis, E.A.G.; Nakatani, M.; Badaró, R.; et al. Domestic swine in a visceral leishmaniasis endemic area produce antibodies against multiple leishmania infantum antigens but apparently resist to L. infantum infection. Acta Trop. 2006, 98, 176–182. [Google Scholar] [CrossRef] [Green Version]
- Navea-Pérez, H.M.; Díaz-Sáez, V.; Corpas-López, V.; Merino-Espinosa, G.; Morillas-Márquez, F.; Martín-Sánchez, J. Leishmania infantum in wild rodents: Reservoirs or just irrelevant incidental hosts? Parasitol. Res. 2015, 114, 2363–2370. [Google Scholar] [CrossRef]
- Tsokana, C.N.; Sokos, C.; Giannakopoulos, A.; Birtsas, P.; Athanasiou, L.V.; Valiakos, G.; Sofia, M.; Chatzopoulos, D.C.; Kantere, M.; Spyrou, V.; et al. Serological and molecular investigation of selected parasitic pathogens in European brown hare (Lepus Europaeus) in Greece: Inferring the ecological niche of toxoplasma gondii and leishmania infantum in hares. Parasitol. Res. 2019, 118, 2715–2721. [Google Scholar] [CrossRef] [PubMed]
- Abbate, J.M.; Arfuso, F.; Napoli, E.; Gaglio, G.; Giannetto, S.; Latrofa, M.S.; Otranto, D.; Brianti, E. Leishmania infantum in wild animals in endemic areas of southern Italy. Comp. Immunol. Microbiol. Infect. Dis. 2019, 67, 101374. [Google Scholar] [CrossRef]
- El Hamouchi, A.; El Kacem, S.; Ejghal, R.; Lemrani, M. Genetic polymorphism in leishmania infantum isolates from human and animals determined by nagt PCR-RFLP. Infect. Dis. Poverty 2018, 7, 54. [Google Scholar] [CrossRef]
- Solano-Gallego, L.; Koutinas, A.; Miró, G.; Cardoso, L.; Pennisi, M.G.; Ferrer, L.; Bourdeau, P.; Oliva, G.; Baneth, G. Directions for the diagnosis, clinical staging, treatment and prevention of canine leishmaniosis. Vet. Parasitol. 2009, 165, 1–18. [Google Scholar] [CrossRef]
- Morillas, F.; Sanchez Rabasco, F.; Ocaña, J.; Martin-Sanchez, J.; Ocaña-Wihelmi, J.; Acedo, C.; Sanchiz-Marin, M.C. Leishmaniosis in the focus of the Axarquía region, Malaga province, southern Spain: A survey of the human, dog, and vector. Parasitol. Res. 1996, 82, 569–570. [Google Scholar] [CrossRef] [PubMed]
- Amusategui, I.; Sainz, A.; Aguirre, E.; Tesouro, M.A. Seroprevalence of leishmania infantum in northwestern Spain, an area traditionally considered free of leishmaniasis. Ann. N. Y. Acad. Sci. 2004, 1026, 154–157. [Google Scholar] [CrossRef] [PubMed]
- Otranto, D.; Napoli, E.; Latrofa, M.S.; Annoscia, G.; Tarallo, V.D.; Greco, G.; Lorusso, E.; Gulotta, L.; Falsone, L.; Basano, F.S.; et al. Feline and canine leishmaniosis and other vector-borne diseases in the Aeolian islands: Pathogen and vector circulation in a confined environment. Vet. Parasitol. 2017, 236, 144–151. [Google Scholar] [CrossRef]
- Tamponi, C.; Scarpa, F.; Carta, S.; Knoll, S.; Sanna, D.; Gai, C.; Pipia, A.P.; Dessì, G.; Casu, M.; Varcasia, A.; et al. Seroprevalence and risk factors associated with leishmania infantum in dogs in Sardinia (Italy), an endemic island for leishmaniasis. Parasitol. Res. 2021, 120, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Burnham, A.C.; Ordeix, L.; Alcover, M.M.; Martínez-Orellana, P.; Montserrat-Sangrà, S.; Willen, L.; Spitzova, T.; Volf, P.; Solano-Gallego, L. Exploring the relationship between susceptibility to canine leishmaniosis and anti-phlebotomus perniciosus saliva antibodies in Ibizan hounds and dogs of other breeds in Mallorca, Spain. Parasites Vectors 2020, 13, 129. [Google Scholar] [CrossRef]
- Gálvez, R.; Montoya, A.; Cruz, I.; Fernández, C.; Martín, O.; Checa, R.; Chicharro, C.; Migueláñez, S.; Marino, V.; Miró, G. Latest trends in leishmania infantum infection in dogs in Spain, part I: Mapped seroprevalence and sand fly distributions. Parasites Vectors 2020, 13, 204. [Google Scholar] [CrossRef] [Green Version]
- De Araujo, F.J.; da Silva, L.D.O.; Mesquita, T.G.; Pinheiro, S.K.; Wonei de Seixas, V.; Chrusciak-Talhari, A.; de Oliveira Guerra, J.A.; Talhari, S.; Ramasawmy, R. Polymorphisms in the TOLLIP gene influence susceptibility to cutaneous leishmaniasis caused by leishmania guyanensis in the Amazonas State of Brazil. PLoS Negl. Trop. Dis. 2015, 9, e0003875. [Google Scholar] [CrossRef]
- De Vasconcelos, T.C.B.; Doyen, N.; Cavaillon, J.-M.; Bruno, S.F.; de Campos, M.P.; de Miranda, L.H.M.; de Fátima Madeira, M.; Belo, V.S.; Figueiredo, F.B. Cytokine and INOS profiles in lymph nodes of dogs naturally infected with leishmania infantum and their association with the parasitic DNA load and clinical and histopathological features. Vet. Parasitol. 2016, 227, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Rivera, M.P.; Ramírez-Ramírez, A.; Chiñas-Pérez, A.; Monroy-Ostria, A.; Cancino-Díaz, M.E.; Hernández-Montes, O. NRAMP1 Polymorphisms like susceptibility marker in Mexican focus of cutaneous leishmaniasis. Biomed. Res. Int. 2016, 2016, 7951285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.; Tiwary, P.; Chakravarty, J.; Sundar, S. Association of Interleukin-18 gene polymorphism with susceptibility to visceral leishmaniasis in endemic area of Bihar, an Indian population. Sci. World J. 2014, 2014, 852104. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Flores, A.; De la Rosa-López, G.; Zavaleta-Villa, B.; Chávez-López, S.; Pastor-Santiago, J.; Guzmán-Bracho, C.; Romero-Valdovinos, M.; Martínez-Hernández, F.; Olivo-Díaz, A. Association of leishmaniasis with TNF alpha promoter and SLC11A1 gene polymorphisms in patients of two endemic areas in Mexico. Microbes Infect. 2015, 17, 387–394. [Google Scholar] [CrossRef]
- Ready, P.D. Epidemiology of visceral leishmaniasis. Clin. Epidemiol. 2014, 6, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sophie, M.; Hameed, A.; Muneer, A.; Samdani, A.J.; Saleem, S.; Azhar, A. SLC11A1 Polymorphisms and host susceptibility to cutaneous leishmaniasis in Pakistan. Parasites Vectors 2017, 10, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soutter, F.; Solano-Gallego, L.; Attipa, C.; Gradoni, L.; Fiorentino, E.; Foglia Manzillo, V.; Oliva, G.; Tasker, S.; Helps, C.; Catchpole, B. An investigation of polymorphisms in innate and adaptive immune response genes in canine leishmaniosis. Vet. Parasitol. 2019, 269, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Solano-Gallego, L.; Llull, J.; Ramos, G.; Riera, C.; Arboix, M.; Alberola, J.; Ferrer, L. The Ibizian hound presents a predominantly cellular immune response against natural leishmania infection. Vet. Parasitol. 2000, 90, 37–45. [Google Scholar] [CrossRef]
- De Vasconcelos, T.C.B.; Furtado, M.C.; Belo, V.S.; Morgado, F.N.; Figueiredo, F.B. Canine susceptibility to visceral leishmaniasis: A systematic review upon genetic aspects, considering breed factors and immunological concepts. Infect. Genet. Evol. 2019, 74, 103293. [Google Scholar] [CrossRef]
- Barbiéri, C.L. Immunology of canine leishmaniasis. Parasite Immunol. 2006, 28, 329–337. [Google Scholar] [CrossRef]
- Gradoni, L. Canine leishmania vaccines: Still a long way to go. Vet. Parasitol. 2015, 208, 94–100. [Google Scholar] [CrossRef]
- Balaña-Fouce, R.; Calvo-Álvarez, E.; Álvarez-Velilla, R.; Prada, C.F.; Pérez-Pertejo, Y.; Reguera, R.M. Role of Trypanosomatid’s arginase in polyamine biosynthesis and pathogenesis. Mol. Biochem. Parasitol. 2012, 181, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Guerra, J.M.; Fernandes, N.C.C.A.; Réssio, R.A.; Kimura, L.M.; Barbosa, J.E.R.; Taniguchi, H.H.; Hiramoto, R.M.; Motoie, G.; Tolezano, J.E.; Cogliati, B. Cytokine profile and parasite load in lymph nodes of dogs naturally infected with leishmania infantum from distinct epidemiological scenarios in São Paulo State, Brazil. Vet. Immunol. Immunopathol. 2021, 233, 110198. [Google Scholar] [CrossRef]
- Verçosa, B.L.A.; Muniz-Junqueira, M.I.; Menezes-Souza, D.; Fujiwara, R.T.; Melo, M.N.; Vasconcelos, A.C. The balance between IL-12/IL4 in renal tissue switches the inflammatory response arm and shows relationship with the clinical signs in leishmania-infected dogs. Vet. Immunol. Immunopathol. 2021, 234, 110196. [Google Scholar] [CrossRef] [PubMed]
- Batista, L.F.S.; Utsunomiya, Y.T.; Silva, T.B.F.; Dias, R.A.; Tomokane, T.Y.; Pacheco, A.D.; da Matta, V.L.R.; Silveira, F.T.; Marcondes, M.; Nunes, C.M.; et al. Genome-wide association study of cell-mediated response in dogs naturally infected by leishmania infantum. Infect. Immun. 2016, 84, 3629–3637. [Google Scholar] [CrossRef] [Green Version]
- De Oliveira Cardoso, J.M.; de Brito, R.C.F.; Costa, A.F.P.; Siqueira Mathias, F.A.; Soares Reis, L.E.; Vieira, J.F.P.; de Oliveira Aguiar Soares, R.D.; Reis, A.B.; Roatt, B.M. IL-10 receptor blockade controls the In Vitro infectivity of leishmania infantum and promotes a Th1 activation in PBMC of dogs with visceral leishmaniasis. Mol. Immunol. 2021, 137, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.d.O.; Costa, S.F.; Souza, F.S.; Mendes, J.M.F.; de Pinheiro, C.G.M.; Moreira, D.R.d.M.; Silva, L.K.; de Lima, V.M.F.; de Sá Oliveira, G.G. Blocking IL-10 signaling with soluble IL-10 receptor restores In Vitro specific lymphoproliferative response in dogs with leishmaniasis caused by leishmania infantum. PLoS ONE 2021, 16, e0239171. [Google Scholar] [CrossRef]
- Utsunomiya, Y.T.; Ribeiro, É.S.; Quintal, A.P.N.; Sangalli, J.R.; Gazola, V.R.; Paula, H.B.; Trinconi, C.M.; Lima, V.M.F.; Perri, S.H.V.; Taylor, J.F.; et al. Genome-wide scan for visceral leishmaniasis in mixed-breed dogs identifies candidate genes involved in T helper cells and macrophage signaling. PLoS ONE 2015, 10, e0136749. [Google Scholar] [CrossRef]
- Quilez, J.; Martínez, V.; Woolliams, J.A.; Sanchez, A.; Pong-Wong, R.; Kennedy, L.J.; Quinnell, R.J.; Ollier, W.E.R.; Roura, X.; Ferrer, L.; et al. Genetic control of canine leishmaniasis: Genome-wide association study and genomic selection analysis. PLoS ONE 2012, 7, e35349. [Google Scholar] [CrossRef] [Green Version]
- Batista, L.F.S.; Torrecilha, R.B.P.; Silva, R.B.; Utsunomiya, Y.T.; Silva, T.B.F.; Tomokane, T.Y.; Pacheco, A.D.; Bosco, A.M.; Paulan, S.C.; Rossi, C.N.; et al. Chromosomal segments may explain the antibody response cooperation for canine leishmaniasis pathogenesis. Vet. Parasitol. 2020, 288, 109276. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Robert, E.; Altet, L.; Utzet-Sadurni, M.; Giger, U.; Sanchez, A.; Francino, O. Slc11a1 (formerly Nramp1) and susceptibility to canine visceral leishmaniasis. Vet. Res. 2008, 39, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Searle, S.; Blackwell, J.M. Evidence for a functional repeat polymorphism in the promoter of the human NRAMP1 gene that correlates with autoimmune versus infectious disease susceptibility. J. Med. Genet. 1999, 36, 295–299. [Google Scholar] [PubMed]
- Miró, G.; Checa, R.; Montoya, A.; Hernández, L.; Dado, D.; Gálvez, R. Current situation of leishmania infantum infection in shelter dogs in northern Spain. Parasites Vectors 2012, 5, 60. [Google Scholar] [CrossRef] [Green Version]
- Olías-Molero, A.I.; Corral, M.J.; Jiménez-Antón, M.D.; Alunda, J.M. Early antibody response and clinical outcome in experimental canine leishmaniasis. Sci. Rep. 2019, 9, 18606. [Google Scholar] [CrossRef]
- Puntas, M.G.S. Estudio Epidemiologico de la Leishmaniosis en la Isla de Menorca. Ph.D. Thesis, Universitat de Barcelona, Barcelona, Spain, 1991. [Google Scholar]
- Chalghaf, B.; Chemkhi, J.; Mayala, B.; Harrabi, M.; Benie, G.B.; Michael, E.; Ben Salah, A. Ecological niche modeling predicting the potential distribution of leishmania vectors in the Mediterranean basin: Impact of climate change. Parasites Vectors 2018, 11, 461. [Google Scholar] [CrossRef]
- Martínez-Orellana, P.; Marí-Martorell, D.; Montserrat-Sangrà, S.; Ordeix, L.; Baneth, G.; Solano-Gallego, L. Leishmania infantum-specific IFN-γ production in stimulated blood from dogs with clinical leishmaniosis at diagnosis and during treatment. Vet. Parasitol. 2017, 248, 39–47. [Google Scholar] [CrossRef]
- Sanchez-Robert, E.; Altet, L.; Sanchez, A.; Francino, O. Polymorphism of Slc11a1 (Nramp1) gene and canine leishmaniasis in a case-control study. J. Hered. 2005, 96, 755–758. [Google Scholar] [CrossRef]
- Abranches, P.; Silva-Pereira, M.C.; Conceição-Silva, F.M.; Santos-Gomes, G.M.; Janz, J.G. Canine leishmaniasis: Pathological and ecological factors influencing transmission of infection. J. Parasitol. 1991, 77, 557–561. [Google Scholar] [CrossRef]
- Parker, H.G.; Dreger, D.L.; Rimbault, M.; Davis, B.W.; Mullen, A.B.; Carpintero-Ramirez, G.; Ostrander, E.A. Genomic analyses reveal the influence of geographic origin, migration, and hybridization on modern dog breed development. Cell Rep. 2017, 19, 697–708. [Google Scholar] [CrossRef] [PubMed]
- Vonholdt, B.M.; Pollinger, J.P.; Lohmueller, K.E.; Han, E.; Parker, H.G.; Quignon, P.; Degenhardt, J.D.; Boyko, A.R.; Earl, D.A.; Auton, A.; et al. Genome-wide SNP and haplotype analyses reveal a rich history underlying dog domestication. Nature 2010, 464, 898–902. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, C.S.; Ratzlaff, F.R.; Pötter, L.; Romão, P.R.T.; de Avila Botton, S.; Vogel, F.S.F.; Sangioni, L.A. Clinical and pathological aspects of canine cutaneous leishmaniasis: A meta-analysis. Acta Parasitol. 2019, 64, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Viol, M.A.; Lima, V.M.F.; Aquino, M.C.C.; Gallo, G.; Alves, I.P.; Generoso, D.; Perri, S.H.V.; Lucheis, S.B.; Langoni, H.; Nunes, C.M.; et al. Detection of cross infections by Leishmania Spp. and Trypanosoma Spp. in dogs using indirect immunoenzyme assay, indirect fluorescent antibody test and polymerase chain reaction. Parasitol. Res. 2012, 111, 1607–1613. [Google Scholar] [CrossRef]
- Zaffaroni, E.; Rubaudo, L.; Lanfranchi, P.; Mignone, W. Epidemiological patterns of canine leishmaniasis in Western Liguria (Italy). Vet. Parasitol. 1999, 81, 11–19. [Google Scholar] [CrossRef]
- Dantas-Torres, F.; de Brito, M.E.F.; Brandão-Filho, S.P. Seroepidemiological survey on canine leishmaniasis among dogs from an urban area of Brazil. Vet. Parasitol. 2006, 140, 54–60. [Google Scholar] [CrossRef]
- Gálvez, R.; Miró, G.; Descalzo, M.A.; Nieto, J.; Dado, D.; Martín, O.; Cubero, E.; Molina, R. Emerging trends in the seroprevalence of canine leishmaniasis in the Madrid region (Central Spain). Vet. Parasitol. 2010, 169, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Mhadhbi, M.; Chaabouni, A.; Bouabid, C.; Sassi, A. Relationships between specific antibody responses and clinical signs of dogs living in Tunisian endemic areas of canine leishmaniasis caused by leishmania infantum. Acta Trop. 2021, 218, 105906. [Google Scholar] [CrossRef]
- Fernandes, M.A.; Leonel, J.A.F.; Isaac, J.A.; Benassi, J.C.; Silva, D.T.; Spada, J.C.P.; Pereira, N.W.B.; Ferreira, H.L.; Keid, L.B.; Soares, R.M.; et al. Molecular detection of leishmania infantum DNA according to clinical stages of leishmaniasis in dog. Rev. Bras. Parasitol. Vet. 2019, 28, 194–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amusategui, I.; Sainz, A.; Rodríguez, F.; Tesouro, M.A. Distribution and relationships between clinical and biopathological parameters in canine leishmaniasis. Eur. J. Epidemiol. 2003, 18, 147–156. [Google Scholar] [CrossRef]

{kind=link}
| Clinical Stage | Clinical Signs |
|---|---|
| Stage I: mild disease | Mild clinical signs, peripheral lymphadenopathy, or popular dermatitis. |
| Stage II: moderate disease | Clinical signs of stage I include diffuse or symmetrical cutaneous lesions (exfoliative dermatitis, ulcerations), anorexia, weight loss, fever, and epistaxis. |
| Stage III: severe disease | Clinical signs of stages I and II, include vasculitis, arthritis, uveitis, and glomerulonephritis. |
| Stage IV: very severe disease | Clinical signs of stages I, II and III, include pulmonary thromboembolism, or nephrotic syndrome, and end-stage renal disease. |
| Breed | Studied Dogs | Seropositive Dogs | Seroprevalence (%) |
|---|---|---|---|
| Beagle | 16 | 1 | 6.25 ab |
| Maltese Bichon | 68 | 1 | 1.47 a |
| Boxer | 23 | 9 | 39.13 c |
| Bull Terrier | 16 | 2 | 12.50 bc |
| French Bulldog | 426 | 6 | 1.41 a |
| Pug | 138 | 1 | 0.72 a |
| Chihuahua | 172 | 4 | 2.33 a |
| Cocker Spaniel | 41 | 11 | 26.83 c |
| Doberman Pinscher | 7 | 3 | 42.86 c |
| Dogue de Bordeaux | 3 | 1 | 33.33 c |
| Golden Retriever | 23 | 2 | 8.70 bc |
| Great Dane | 6 | 2 | 33.33 c |
| Jack Russell | 34 | 3 | 8.82 bc |
| Labrador Retriever | 103 | 14 | 13.59 bc |
| Mastiff | 22 | 2 | 9.09 bc |
| German Shepherd | 271 | 20 | 7.38 b |
| Majorcan Shepherd | 3 | 1 | 33.33 c |
| Pinscher | 55 | 2 | 3.64 ab |
| Pitbull | 18 | 1 | 5.56 ab |
| Pointer | 49 | 12 | 24.49 c |
| Pomeranian | 14 | 2 | 14.29 bc |
| Schnauzer | 8 | 1 | 12.50 bc |
| Spitz | 6 | 1 | 16.67 bc |
| Weimaraner | 3 | 1 | 33.33 c |
| Yorkshire | 47 | 3 | 6.38 ab |
| Crossbreed | 1569 | 43 | 2.74 a |
| Overall | 3141 | 149 | 4.74 |
| p-value | <0.05 |
| Variable Analyzed | Categorical Factors | Distribution of Seropositive (%) |
|---|---|---|
| Sex | Male | 60.26 |
| Female | 39.74 | |
| Age | Young (<4 years) | 54.97 |
| Adult (4 ≤ years < 10) | 38.41 | |
| Older (≥10 years) | 6.62 | |
| Presentation of disease | Cutaneous | 62.91 |
| Visceral | 35.10 | |
| Both | 1.99 | |
| Clinical stage | I | 15.89 |
| II | 58.28 | |
| III | 24.50 | |
| IV | 1.32 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Edo, M.; Marín-García, P.J.; Llobat, L. Is the Prevalence of Leishmania infantum Linked to Breeds in Dogs? Characterization of Seropositive Dogs in Ibiza. Animals 2021, 11, 2579. https://doi.org/10.3390/ani11092579
Edo M, Marín-García PJ, Llobat L. Is the Prevalence of Leishmania infantum Linked to Breeds in Dogs? Characterization of Seropositive Dogs in Ibiza. Animals. 2021; 11(9):2579. https://doi.org/10.3390/ani11092579
Chicago/Turabian StyleEdo, Maria, Pablo Jesús Marín-García, and Lola Llobat. 2021. "Is the Prevalence of Leishmania infantum Linked to Breeds in Dogs? Characterization of Seropositive Dogs in Ibiza" Animals 11, no. 9: 2579. https://doi.org/10.3390/ani11092579
APA StyleEdo, M., Marín-García, P. J., & Llobat, L. (2021). Is the Prevalence of Leishmania infantum Linked to Breeds in Dogs? Characterization of Seropositive Dogs in Ibiza. Animals, 11(9), 2579. https://doi.org/10.3390/ani11092579