Nitrogen Balance of Dairy Cows Divergent for Milk Urea Nitrogen Breeding Values Consuming Either Plantain or Perennial Ryegrass

 ,
,  ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Herbage Measurements
2.2. Animal Measurements
2.3. Statistical Analysis
3. Results
3.1. Herbage Measurements
3.2. Herbage Intake and Digestibility
3.3. Milk Production
3.4. Rumen
3.5. Purine Excretion and Microbial Crude Protein Flow and Efficiency
3.6. Excretory Pattern
3.7. Water Balance
3.8. Nitrogen Balance and Partitioning
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pacheco, D.; Waghorn, G. Dietary nitrogen—Definitions, digestion, excretion and consequences of excess for grazing ruminants. Proc. N. Z. Grassl. Assoc. 2008, 70, 107–116. [Google Scholar] [CrossRef]
- Castillo, A.R.; Kebreab, E.; Beever, D.E.; Barbi, J.H.; Sutton, J.D.; Kirby, H.C.; France, J. The effect of protein supplementation on nitrogen utilization in lactating dairy cows fed grass silage diets. J. Anim. Sci. 2001, 79, 247–253. [Google Scholar] [CrossRef]
- Kebreab, E.; France, J.; Beever, D.E.; Castillo, A.R. Nitrogen pollution by dairy cows and its mitigation by dietary manipulation. Nutr. Cycl. Agroecosyst. 2001, 60, 275–285. [Google Scholar] [CrossRef]
- Gregorini, P.; Beukes, P.C.; Dalley, D.; Romera, A.J. Screening for diets that reduce urinary nitrogen excretion and methane emissions while maintaining or increasing production by dairy cows. Sci. Total Environ. 2016, 551–552, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Oudshoorn, F.W.; Kristensen, T.; Nadimi, E. Dairy cow defecation and urination frequency and spatial distribution in relation to time-limited grazing. Livest. Sci. 2008, 113, 62–73. [Google Scholar] [CrossRef]
- Clark, C.; Waghorn, G.; Gregorini, P.; Woodward, S.; Clark, D. Diurnal pattern of urinary and faecal nitrogen excretion by dairy cows fed ryegrass pasture twice daily indoors. Adv. Anim. Biosci. 2010, 2, 269. [Google Scholar]
- Selbie, D.R.; Buckthought, L.E.; Shepherd, M.A. The challenge of the urine patch for managing nitrogen in grazed pasture systems. Adv. Agron. 2015, 129, 229–292. [Google Scholar]
- Eggleston, H.S.; Buendia, L.; Miwa, K.; Ngara, T.; Tanabe, K. IPCC Guidelines for National Greenhouse Gase Inventories; IPCC: Kanagawa, Japan, 2006. [Google Scholar]
- Del Grosso, S.J.; Wirth, T.; Ogle, S.; Parton, W.J. Estimating agricultural nitrous oxide emissions. Eos 2008, 89, 529. [Google Scholar] [CrossRef]
- Schallenberg, M.; de Winton, M.D.; Verburg, P.; Kelly, D.J.; Hamill, K.D.; Hamilton, D.P. Ecosystem services of lakes. In Ecosystem Services in New Zealand—Conditions and Trends; Dymond, J.R., Ed.; Manaaki Whenua Press: Lincoln, New Zealand, 2013. [Google Scholar]
- Gregorini, P.; Maxwell, T. Grazing in future multiscapes—From thoughtscapes to ethical and sustainable foodscapes. Journal 2020, 24, 23–25. [Google Scholar]
- United Nations. Sustainable Development Goals. 2015. Available online: https://sustainabledevelopment.un.org/sdgs (accessed on 17 March 2021).
- Schullehner, J.; Hansen, B.; Thygesen, M.; Pedersen, C.B.; Sigsgaard, T. Nitrate in drinking water and colorectal cancer risk: A nationwide population-based cohort study. Int. J. Cancer 2018, 143, 73–79. [Google Scholar] [CrossRef]
- Johnson, S.F. Methemoglobinemia: Infants at risk. Curr. Probl. Pediatr. Adolesc. Health Care 2019, 49, 57–67. [Google Scholar] [CrossRef]
- Brender, J.D.; Olive, J.M.; Felkner, M.; Suarez, L.; Marckwardt, W.; Hendricks, K.A. Dietary nitrites and nitrates, nitrosatable drugs, and neural tube defects. Epidemiology 2004, 15, 330–336. [Google Scholar] [CrossRef]
- Thomson, B.C.; Ward, K.; Smith, N.; Gibbs, J.; Muir, P.D. Effect of feeding time on urinary and faecal nitrogen excretion patterns in sheep. N. Z. J. Agric. Res. 2021, 64, 314–319. [Google Scholar] [CrossRef]
- Beukes, P.C.; Gregorini, P.; Romera, A.J.; Woodward, S.L.; Khaembah, E.; Chapman, D.F.; Nobilly, F.; Bryant, R.H.; Edwards, G.; Clark, D.A. The potential of diverse pastures to reduce nitrogen leaching on New Zealand dairy farms. Anim. Prod. Sci. 2014, 54, 1971–1979. [Google Scholar] [CrossRef] [Green Version]
- Garrett, K.; Beck, M.R.; Marshall, C.J.; Fleming, A.; Logan, C.M.; Maxwell, T.M.R.; Greer, A.W.; Gregorini, P. Functional diversity vs. monotony: The effect of a multiforage diet as opposed to a single forage diet on animal intake, performance, welfare, and urinary nitrogen excretion. J. Anim. Sci. 2021, 99, skab058. [Google Scholar] [CrossRef]
- Carmona-Flores, L.; Bionaz, M.; Downing, T.; Sahin, M.; Cheng, L.; Ates, S. Milk production, N partitioning, and Methane emissions in dairy cows grazing mixed or spatially separated simple and diverse pastures. Animals 2020, 10, 1301. [Google Scholar] [CrossRef] [PubMed]
- Box, L.A.; Edwards, G.; Bryant, R.H. Milk production and urinary nitrogen excretion of dairy cows grazing plantain in early and late lactation. N. Z. J. Agric. Res. 2017, 60, 470–482. [Google Scholar] [CrossRef]
- Bryant, R.H.; Snow, V.O.; Shorten, P.R.; Welten, B.G. Can alternative forages substantially reduce N leaching? Findings from a review and associated modelling. N. Z. J. Agric. Res. 2019, 63, 3–28. [Google Scholar] [CrossRef]
- Stewart, A. Plantain (Plantago lanceolata)–A potential pasture species. Proc. N. Z. Grassl. Assoc. 1996, 86, 77–86. [Google Scholar] [CrossRef]
- Navarrete, S.; Kemp, P.D.; Pain, S.J.; Back, P.J. Bioactive compounds, aucubin and acteoside, in plantain (Plantago lanceolata L.) and their effect on in vitro rumen fermentation. Anim. Feed Sci. Technol. 2016, 222, 158–167. [Google Scholar] [CrossRef]
- O’Connell, C.; Judson, H.; Barrell, G.K. Sustained diuretic effect of plantain when ingested by sheep. N. Z. Soc. Anim. Prod. 2016, 76, 14–17. [Google Scholar]
- Butler, W.; Calaman, J.J.; Beam, S.W. Plasma and milk urea nitrogen in relation to pregnancy rate in lactating dairy cattle. J. Anim. Sci. 1996, 74, 858–865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kauffman, A.; St-Pierre, N. The relationship of milk urea nitrogen to urine nitrogen excretion in holstein and jersey cows. J. Dairy Sci. 2001, 84, 2284–2294. [Google Scholar] [CrossRef]
- Kohn, R.; Kalscheur, K.; Russek-Cohen, E. Evaluation of models to estimate urinary nitrogen and expected milk urea Nitrogen. J. Dairy Sci. 2002, 85, 227–233. [Google Scholar] [CrossRef]
- Kohn, R.A.; Dinneen, M.M.; Russek-Cohen, E. Using blood urea nitrogen to predict nitrogen excretion and efficiency of nitrogen utilization in cattle, sheep, goats, horses, pigs, and rats. J. Anim. Sci. 2005, 83, 879–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoop, W.; Bovenhuis, H.; van Arendonk, J. Genetic parameters for milk urea Nitrogen in relation to milk production traits. J. Dairy Sci. 2007, 90, 1981–1986. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Villalobos, N.; Correa-Luna, M.; Burke, J.L.; Sneddon, N.W.; Schutz, M.; Donaghy, D.; Kemp, P. Genetic parameters for milk urea concentration and milk traits in New Zealand grazing dairy cattle. N. Z. J. Anim. Sci. Prod. 2018, 78, 56–61. [Google Scholar]
- Beatson, P.R.; Meier, S.; Cullen, N.G.; Eding, H. Genetic variation in milk urea nitrogen concentration of dairy cattle and its implications for reducing urinary nitrogen excretion. Animal 2019, 13, 2164–2171. [Google Scholar] [CrossRef] [Green Version]
- Marshall, C.; Beck, M.; Garrett, K.; Barrell, G.; Al-Marashdeh, O.; Gregorini, P. Grazing dairy cows with low milk urea nitrogen breeding values excrete less urinary urea nitrogen. Sci. Total Environ. 2020, 739, 139994. [Google Scholar] [CrossRef] [PubMed]
- Primary Industries Standing Committee. Nutrient Requirements of Domesticated Ruminants; CSIRO Publishing: Clayton, Australia, 2007. [Google Scholar] [CrossRef]
- Iowerth, D.; Jones, H.; Hayward, M.V. The effect of pepsin pretreatment of herbage on the prediction of dry matter digestibility from solubility in fungal cellulase solutions. J. Sci. Food Agric. 1975, 26, 711–718. [Google Scholar] [CrossRef]
- MAFF—Ministry of Agriculture, Fisheries and Food. Carbohydrates, soluble, in herbage. In The Analysis of Agricultural Materials: A Manual of the Analytical Methods Used by the Agricultural Development and Advisory Service; MAFF: London, UK, 1986. [Google Scholar]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemist: Washington, DC, USA, 1990. [Google Scholar]
- Van Soest, P.; Robertson, J.; Lewis, B. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Haynes, R.J.; Williams, P.H. Nutrient cycling and soil fertility in grazed pature ecosystem. Adv. Agron. 1993, 49, 119–199. [Google Scholar]
- Romera, A.J.; Levy, G.; Beukes, P.; Clark, D.A.; Glassey, C.B. A urine patch framework to simulate nitrogen leaching on New Zealand dairy farms. Nutr. Cycl. Agroecosyst. 2012, 92, 329–346. [Google Scholar] [CrossRef]
- George, S.; Dipu, M.; Mehra, U.; Singh, P.; Verma, A.; Ramgaokar, J. Improved HPLC method for the simultaneous determination of allantoin, uric acid and creatinine in cattle urine. J. Chromatogr. B 2006, 832, 134–137. [Google Scholar] [CrossRef] [PubMed]
- International Atomic Energy Agency. Estimation of Rumen Microbial Protein Production From Purine Derivatives In Urine—A Laboratory Manual for FAO/IAEA Co-Ordinated Research Program; IAEA: Vienna, Austria, 1997. [Google Scholar]
- Crawford, G.I.; Macdonald, J.C.; Watson, A.K.; Erickson, G.; Klopfenstein, T.J. Diurnal and dietary impacts on estimating microbial protein flow from urinary purine derivative excretion in beef cattle. Transl. Anim. Sci. 2020, 4, txaa140. [Google Scholar] [CrossRef]
- Martín-Orúe, S.M.; Balcells, J.; Guada, J.; Fondevila, M. Microbial nitrogen production in growing heifers: Direct measurement of duodenal flow of purine bases versus urinary excretion of purine derivatives as estimation procedures. Anim. Feed. Sci. Technol. 2000, 88, 171–188. [Google Scholar] [CrossRef]
- Clark, J.; Klusmeyer, T.; Cameron, M. Microbial protein synthesis and flows of Nitrogen fractions to the duodenum of dairy cows. J. Dairy Sci. 1992, 75, 2304–2323. [Google Scholar] [CrossRef]
- Talke, H.; Schubert, G.E. Enzymatische harnstoffbestimmung in blut und serum im optischen test nachwarburg. Klin Wochschr 1965, 43, 174–175. [Google Scholar] [CrossRef]
- Playne, M.J. Determination of ethanol, volatile fatty acids, lactic and succinic acids in fermentation liquids by gas chromatography. J. Sci. Food Agric. 1985, 36, 638–644. [Google Scholar] [CrossRef]
- Neeley, W.E.; Phillipson, J. Automated enzymatic method for determining ammonia in plasma, with 14-day reagent stability. Clin. Chem. 1988, 34, 1868–1869. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Müller, C.B.M.; Görs, S.; Derno, M.; Tuchscherer, A.; Wimmers, K.; Zeyner, A.; Kuhla, B. Differences between Holstein dairy cows in renal clearance rate of urea affect milk urea concentration and the relationship between milk urea and urinary Nitrogen excretion. Sci. Total Environ. 2020, 755, 143198. [Google Scholar] [CrossRef]
- Pacheco, D.; Lowe, K.; Burke, J.L.; Cosgrove, G.P. Urinary nitrogen excretion from cows at different stage of lactation grazing different ryegrass cultivars during spring or autumn. N. Z. Soc. Anim. Sci. 2009, 69, 196–200. [Google Scholar]
- Minnée, E.; Leach, C.; Dalley, D. Substituting a pasture-based diet with plantain (Plantago lanceolata) reduces nitrogen excreted in urine from dairy cows in late lactation. Livest. Sci. 2020, 239, 104093. [Google Scholar] [CrossRef]
- Beck, M.R.; Garrett, K.; Thompson, B.R.; Stevens, D.R.; Barrell, G.K.; Gregorini, P. Plantain (Plantago lanceolata) reduces the environmental impact of farmed red deer (Cervus elaphus). Transl. Anim. Sci. 2020, 4, txaa160. [Google Scholar] [CrossRef]
- Stewart, E.K.; Beauchemin, K.; Dai, X.; Macadam, J.W.; Christensen, R.G.; Villalba, J.J. Effect of tannin-containing hays on enteric methane emissions and nitrogen partitioning in beef cattle. J. Anim. Sci. 2019, 97, 3286–3299. [Google Scholar] [CrossRef] [PubMed]
- Waghorn, G. Beneficial and detrimental effects of dietary condensed tannins for sustainable sheep and goat production—Progress and challenges. Anim. Feed. Sci. Technol. 2008, 147, 116–139. [Google Scholar] [CrossRef]
- Powell, J.; Broderick, G.; Grabber, J.; Hymes-Fecht, U. Technical note: Effects of forage protein-binding polyphenols on chemistry of dairy excreta. J. Dairy Sci. 2009, 92, 1765–1769. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.B. Excretion of Purine Derivatives by Sheep and Cattle and Its Use for the Estimation of Absorbed Microbial Protein. Ph.D. Thesis, University of Aberdeen, Aberdeen, UK, 1989. [Google Scholar]
- Løvendahl, P.; Weisbjerg, M.R. Lactose in milk—How can lactose concentration data be beneficial in management and breeding? ICAR Tech. Ser. 2018, 22, 99–104. [Google Scholar]
- Wong, S.Y.; Hartel, R.W. Crystallization in lactose refining—A review. J. Food Sci. 2014, 79, R257–R272. [Google Scholar] [CrossRef] [PubMed]
- Di, H.J.; Cameron, K. Nitrate leaching losses and pasture yields as affected by different rates of animal urine nitrogen returns and application of a nitrification inhibitor—A lysimeter study. Nutr. Cycl. Agroecosyst. 2007, 79, 281–290. [Google Scholar] [CrossRef]
- Smolders, A.J.P.; Lucassen, E.C.H.E.T.; Bobbink, R.; Roelofs, J.G.M.; Lamers, L.P.M. How nitrate leaching from agricultural lands provokes phosphate eutrophication in groundwater fed wetlands: The sulphur bridge. Biogeochemistry 2009, 98, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Chadwick, D.; Cardenas, L.; Dhanoa, M.; Donovan, N.; Misselbrook, T.; Williams, J.; Thorman, R.; McGeough, K.; Watson, C.; Bell, M.; et al. The contribution of cattle urine and dung to nitrous oxide emissions: Quantification of country specific emission factors and implications for national inventories. Sci. Total Environ. 2018, 635, 607–617. [Google Scholar] [CrossRef]
- Hackmann, T.J.; Firkins, J.L. Maximizing efficiency of rumen microbial protein production. Front. Microbiol. 2015, 6, 465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, V.; Aguilar, M.; Van Amburgh, M.; Nayananjalie, W.; Hanigan, M. Milk urea nitrogen variation explained by differences in urea transport into the gastrointestinal tract in lactating dairy cows. J. Dairy Sci. 2021, 104, 6715–6726. [Google Scholar] [CrossRef]
- Rogers, J.; Marks, B.; Davis, C.; Clark, J. Alteration of rumen fermentation in steers by increasing rumen fluid dilution rate with mineral salts. J. Dairy Sci. 1979, 62, 1599–1605. [Google Scholar] [CrossRef]
- Castillo-González, A.; Burrola-Barraza, M.; Domínguez-Viveros, J.; Chávez-Martínez, A. Rumen microorganisms and fermentation. Arch. Med. Veter. 2014, 46, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Gregorini, P.; Waghorn, G.; Kuhn-Sherlock, B.; Romera, A.; Macdonald, K. Short communication: Grazing pattern of dairy cows that were selected for divergent residual feed intake as calves. J. Dairy Sci. 2015, 98, 6486–6491. [Google Scholar] [CrossRef]
- Beauchemin, K. Invited review: Current perspectives on eating and rumination activity in dairy cows. J. Dairy Sci. 2018, 101, 4762–4784. [Google Scholar] [CrossRef] [Green Version]
- Gregorini, P.; Provenza, F.D.; Villalba, J.J.; Beukes, P.; Forbes, M.J. Diurnal patterns of urination and drinking by grazing ruminants: A development in a mechanistic model of a grazing ruminant, MINDY. J. Agric. Sci. 2018, 156, 71–81. [Google Scholar] [CrossRef] [Green Version]
- Henley, S.S.; Golden, R.M.; Kashner, T.M. Statistical modeling methods: Challenges and strategies. Biostat. Epidemiol. 2019, 4, 105–139. [Google Scholar] [CrossRef] [Green Version]
- Ledgard, S.F.; Wei, S.; Wang, X.; Falconer, S.; Zhang, N.; Zhang, X.; Ma, L. Nitrogen and carbon footprints of dairy farm systems in China and New Zealand, as influenced by productivity, feed sources and mitigations. Agric. Water Manag. 2018, 213, 155–163. [Google Scholar] [CrossRef]
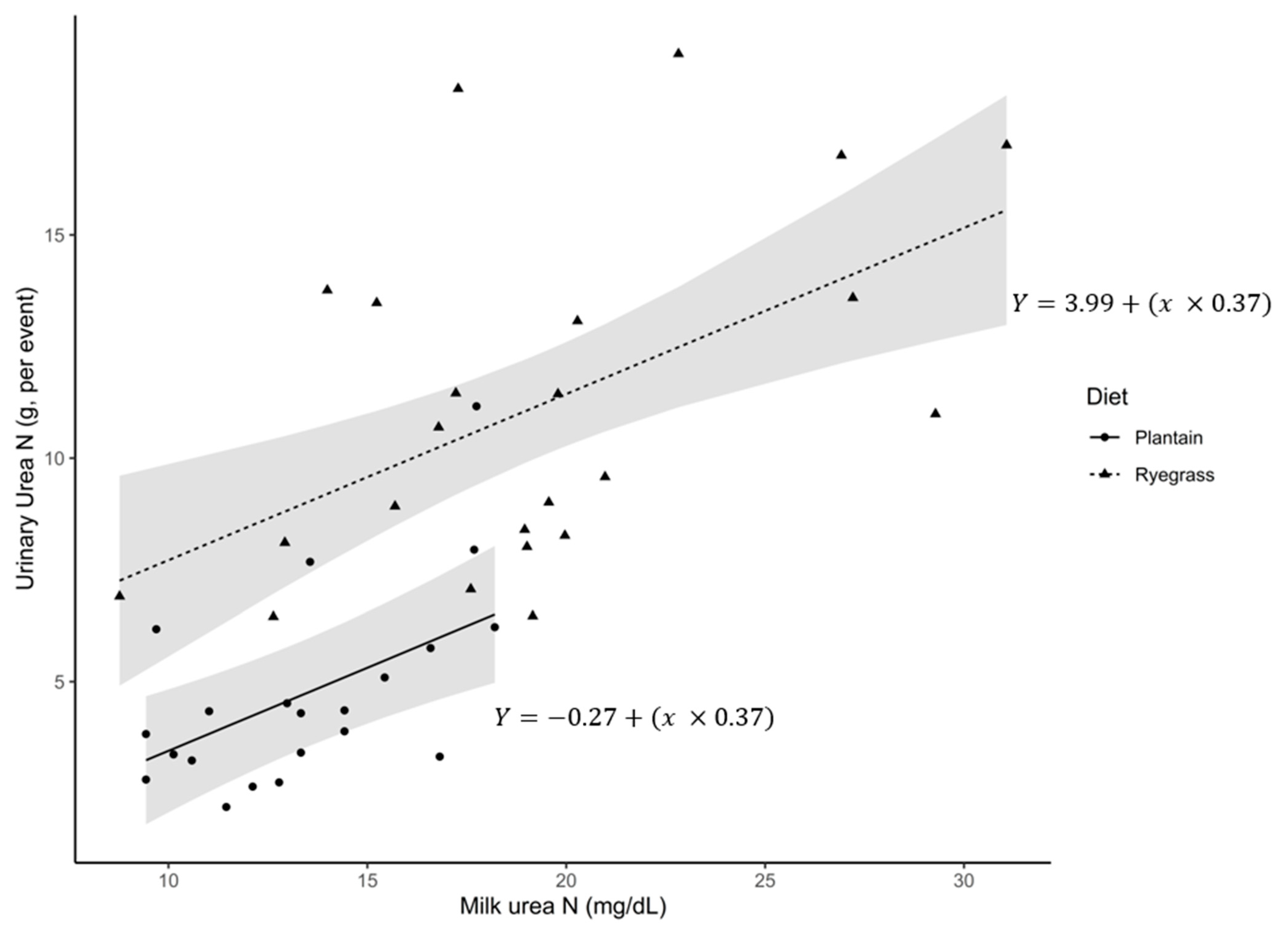

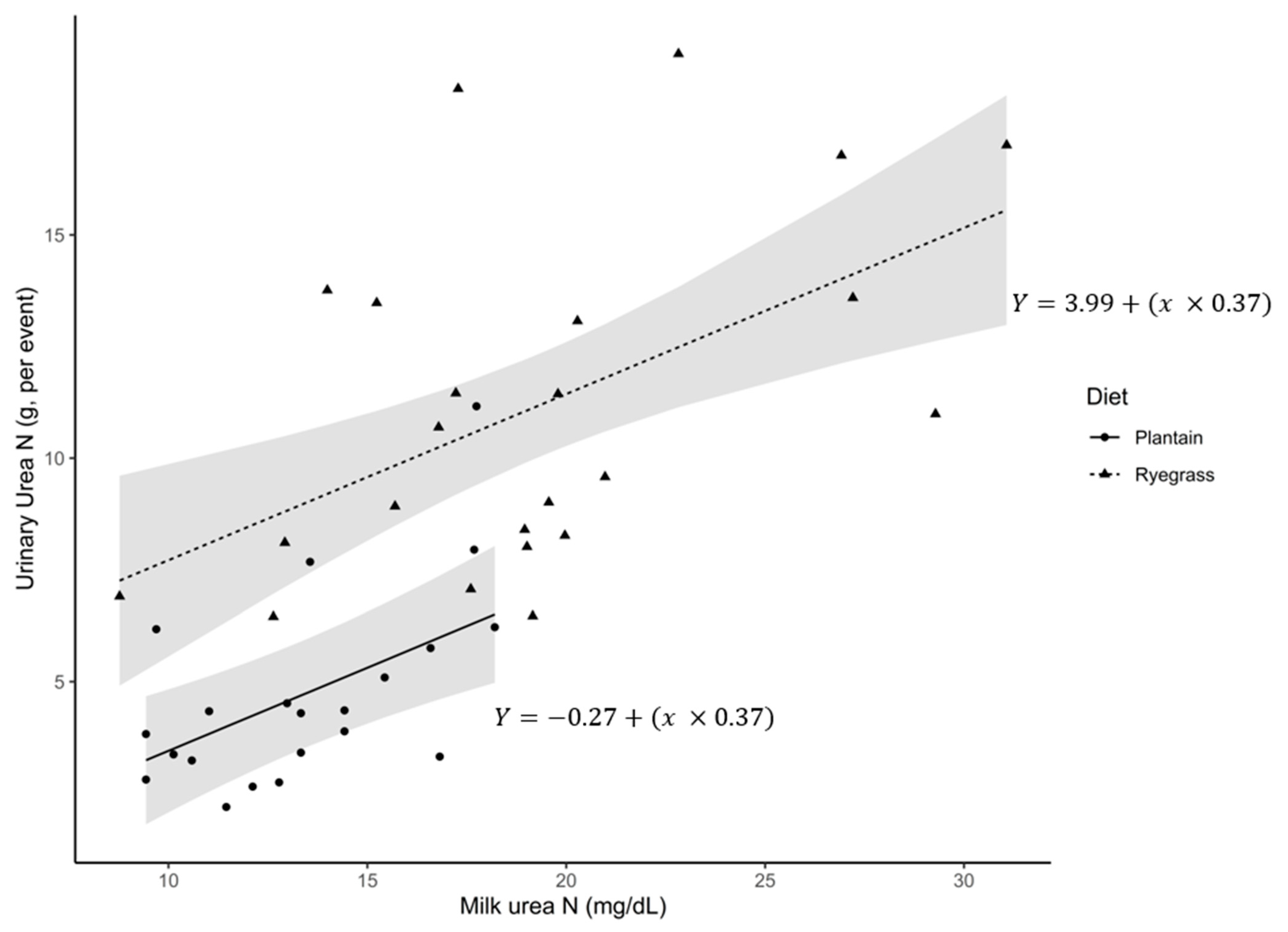{kind=link}
| Diet | p-Value | |||
|---|---|---|---|---|
| Item 1 | Plantain | Ryegrass | RSD 2 | Diet |
| DM, % | 12.8 | 18.5 | 0.01 | <0.01 |
| OM, % | 87.7 | 90.9 | 0.86 | <0.01 |
| CP, % | 13.4 | 14.9 | 2.00 | 0.01 |
| WSC, % | 19.1 | 16.0 | 4.39 | 0.02 |
| NDF, % | 27.3 | 46.5 | 2.45 | <0.01 |
| ADF, % | 24.1 | 26.2 | 1.32 | <0.01 |
| DMD, % | 74.0 | 75.1 | 2.52 | 0.15 |
| OMD, % | 77.6 | 79.5 | 3.13 | 0.04 |
| ME, MJ/kg DM | 11.1 | 11.5 | 0.43 | <0.01 |
| High MUNBV | Low MUNBV | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| Item 1 | Plantain | Ryegrass | Plantain | Ryegrass | RSD 2 | MUNBV | Diet | MUNBV × Diet |
| Dry Matter | 70 | 74.5 | 69.7 | 74.1 | 4.45 | 0.8 | <0.01 | 0.49 |
| Organic Matter | 70 | 76.8 | 69.5 | 76.4 | 3.99 | 0.72 | <0.01 | 0.5 |
| Nitrogen | 64 | 72 | 64.2 | 72.1 | 4.79 | 0.92 | <0.01 | 0.39 |
| ADF | 54.9 | 74.3 | 53.6 | 72 | 0.1 | 0.32 | <0.01 | 0.33 |
| NDF | 46 | 78.4 | 44.7 | 74.8 | 0.13 | 0.36 | <0.01 | 0.22 |
| High MUNBV | Low MUNBV | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| Item 1 | Plantain | Ryegrass | Plantain | Ryegrass | RSD 2 | MUNBV | Diet | MUNBV × Diet |
| Milk yield, kg | 18.6 | 18.3 | 19.3 | 19 | 1.73 | 0.15 | 0.58 | 0.74 |
| Fat, % | 4.54 | 4.78 | 4.51 | 4.75 | 0.01 | 0.86 | 0.19 | 0.3 |
| Fat yield, kg | 0.88 | 0.84 | 0.84 | 0.86 | 0.01 | 0.29 | 0.16 | 0.27 |
| Protein, % | 3.37 | 3.51 | 3.41 | 3.56 | 0.26 | 0.45 | 0.07 | 0.06 |
| Protein yield, kg | 0.62 | 0.64 | 0.65 | 0.67 | 0.05 | 0.06 | 0.37 | 0.44 |
| Lactose yield, kg | 0.93 | 0.92 | 1.01 | 1 | 0.09 | <0.01 | 0.68 | 0.76 |
| Milk solids, kg | 1.43 | 1.48 | 1.49 | 1.53 | 0.11 | 0.09 | 0.17 | 0.2 |
| Total Solids, kg | 2.49 | 2.49 | 2.66 | 2.66 | 0.19 | <0.01 | 0.89 | 0.38 |
| MUN, mg/dl | 13.2 | 19.4 | 11.8 | 15.9 | 0.17 | <0.01 | <0.01 | 0.7 |
| Day 1 | Day 12 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| High MUNBV | Low MUNBV | High MUNBV | Low MUNBV | p-Value | |||||||||
| Item 1 | PL | RG | PL | RG | PL | RG | PL | RG | RSD 2 | Day | MUNBV | Diet | MUNBV × Diet |
| Glucogenic | 21.3 | 21.1 | 20.9 | 20.8 | 17.5 | 17.4 | 17.2 | 17.1 | 0.08 | <0.01 | 0.53 | 0.79 | 0.41 |
| Non-Glucogenic | 76.3 | 76.3 | 76.6 | 76.6 | 80 | 80 | 80.4 | 80.4 | 0.01 | <0.01 | 0.47 | 0.96 | 0.45 |
| A:P ratio | 3.31 | 3.34 | 3.43 | 3.47 | 4.13 | 4.18 | 4.32 | 4.38 | 0.08 | <0.01 | 0.18 | 0.69 | 0.35 |
| NH3 | 7.29 | 6.79 | 7.16 | 6.68 | 5.53 | 5.24 | 5.45 | 5.17 | 0.44 | 0.19 | 0.94 | 0.75 | 0.89 |
| Total VFA | 73.8 | 72.6 | 69.5 | 68.2 | 72.3 | 71.1 | 67.9 | 66.7 | 20.79 | 0.83 | 0.56 | 0.87 | 0.35 |
| Item 1 | High MUNBV | Low MUNBV | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Plantain | Ryegrass | Plantain | Ryegrass | RSD 2 | MUNBV | Diet | MUNBV × Diet | |
| Purine derivatives (mmol/L) | ||||||||
| Creatinine | 1.95 | 2.81 | 1.89 | 2.69 | 0.22 | 0.27 | <0.01 | 0.95 |
| Hippuric | 22.59 | 41.08 | 19.95 | 33.37 | 0.35 | 0.08 | <0.01 | 0.62 |
| Allantoin | 4.99 | 7.81 | 4.8 | 7.37 | 0.24 | 0.42 | <0.01 | 0.65 |
| Uric | 0.39 | 0.43 | 0.43 | 0.48 | 0.03 | 0.26 | 0.34 | 0.28 |
| Total PD | 5.34 | 8.27 | 5.26 | 8.07 | 0.24 | 0.52 | <0.01 | 0.72 |
| Microbial N dynamics | ||||||||
| MCP flow, g/d | 1058 b | 1065 b | 1041 b | 1240 a | 149.23 | 0.09 | 0.03 | 0.04 |
| Microbial efficiency. g of MCP/kg TDOM | 96.5 | 90.3 | 107 | 100.7 | 21.76 | 0.13 | 0.36 | 0.2 |
| Item 1 | High MUNBV | Low MUNBV | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Plantain | Ryegrass | Plantain | Ryegrass | RSD 2 | MUNBV | Diet | MUNBV × Diet | |
| Urination characteristics | ||||||||
| UUN, g/L | 1.69 c | 4.31 a | 1.86 c | 3.29 b | 0.38 | <0.01 | <0.01 | <0.01 |
| UN, g/L | 2.50 c | 5.79 a | 2.70 c | 4.49 b | 0.32 | <0.01 | <0.01 | <0.01 |
| UUN, g/event | 4.53 c | 11.90 a | 4.44 c | 8.62 b | 0.47 | <0.01 | <0.01 | <0.01 |
| UN, g/event | 6.71 c | 16.20 a | 6.50 c | 11.66 b | 0.04 | <0.01 | <0.01 | <0.01 |
| Volume/event, L | 2.63 | 2.83 | 2.47 | 2.65 | 0.34 | 0.01 | <0.01 | 0.16 |
| Events/day | 15.1 a | 9.1 c | 15.2 a | 11.7 b | 0.22 | 0.06 | <0.01 | <0.01 |
| Volume/day, L | 40.4 a | 26.4 c | 39.3 a | 31.0 b | 0.23 | 0.38 | <0.01 | 0.05 |
| UUN, g/d | 67.5 | 109.12 | 68.82 | 102.04 | 0.23 | 0.17 | <0.01 | 0.26 |
| UN, g/d | 108.1 | 147.83 | 103.76 | 139.85 | 0.21 | 0.23 | <0.01 | 0.35 |
| UUN, % UN | 65.7 c | 75.3 a | 66.5 c | 73.5 b | 4.42 | 0.17 | <0.01 | <0.01 |
| Urine patch area, m2 | 0.53 | 0.57 | 0.5 | 0.53 | 0.34 | 0.02 | 0.01 | 0.15 |
| UUN loading rate, kg UUN ha−1 | 90.0 c | 241.1 a | 92.3 c | 173.7 b | 44.45 | <0.01 | <0.01 | <0.01 |
| Faecal characteristics | ||||||||
| N, g/event | 6.65 b | 5.51 c | 6.10 b | 7.75 a | 0.42 | <0.01 | 0.19 | <0.01 |
| OM, g/event | 238.65 b | 183.10 d | 216.28 c | 265.22 a | 0.39 | <0.01 | 0.05 | <0.01 |
| ADF, g/event | 105.47 a | 60.14 d | 97.72 b | 85.66 c | 0.39 | <0.01 | <0.01 | <0.01 |
| NDF, g/event | 144.07 a | 92.23 c | 133.44 b | 133.63 b | 0.4 | <0.01 | <0.01 | <0.01 |
| DM, g/event | 272.11 b | 221.35 d | 243.75 c | 320.07 a | 0.4 | <0.01 | 0.12 | <0.01 |
| Events/day | 17.65 a | 17.81 a | 17.92 a | 12.71 b | 0.23 | 0.04 | 0.04 | 0.03 |
| Total Faecal N/d | 117.66 | 102.5 | 114.69 | 100.24 | 0.18 | 0.57 | 0.01 | 0.47 |
| Total daily, kg/d | 4.7 | 4.12 | 4.54 | 3.99 | 0.01 | 0.37 | <0.01 | 0.12 |
| High MUNBV | Low MUNBV | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| Item | Plantain | Ryegrass | Plantain | Ryegrass | RSD 1 | MUNBV | Diet | MUNBV × Diet |
| Water consumed, L/d | ||||||||
| Trough | 0.8 | 30.3 | 9.4 | 35.5 | 9.83 | 0.03 | <0.01 | 0.21 |
| Forage | 100.5 | 62.8 | 94.9 | 60.6 | 0.12 | 0.12 | <0.01 | 0.87 |
| Total | 101.3 | 93.1 | 104.3 | 96.1 | 13.46 | 0.47 | 0.06 | 0.33 |
| Water excreted, L/d | ||||||||
| Urine | 40.4 a | 26.4 c | 39.3 a | 31.0 b | 0.23 | 0.38 | <0.01 | 0.05 |
| Faeces | 29.8 | 34.4 | 27.5 | 32.2 | 4.28 | 0.09 | <0.01 | 0.88 |
| Milk | 16.1 | 15.7 | 16.7 | 16.3 | 1.56 | 0.3 | 0.45 | 0.76 |
| Total | 87.4 | 77.6 | 85.2 | 79.2 | 0.1 | 0.83 | <0.01 | 0.28 |
| Water balance | 13.9 | 15.5 | 19.1 | 16.9 | 8.19 | 0.34 | 0.36 | 0.63 |
| Water absorbed, % | ||||||||
| Prop. of intake | 70.1 | 62.9 | 73.5 | 66.3 | 4.5 | 0.02 | <0.01 | 0.29 |
| High MUNBV | Low MUNBV | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|
| Item 1 | Plantain | Ryegrass | Plantain | Ryegrass | RSD 2 | MUNBV | Diet | MUNBV × Diet |
| Intake | ||||||||
| DMI, kg/d | 15.5 | 15.8 | 14.9 | 15.3 | 0.14 | 0.41 | 0.57 | 0.96 |
| ME, MJ/d | 179 | 190 | 172 | 183 | 1.92 | 0.41 | 0.19 | 0.95 |
| N, g/d | 338.5 | 390.1 | 327.6 | 379.2 | 56.34 | 0.25 | <0.01 | 0.59 |
| Nitrogen excreted | ||||||||
| Milk, % of intake | 30.1 | 26.8 | 32.9 | 29 | 0.2 | 0.15 | 0.04 | 0.69 |
| Urine, % of intake | 33.1 | 38.4 | 31.5 | 36.4 | 0.26 | 0.4 | 0.02 | 0.32 |
| Faeces, % of intake | 35.4 | 27.3 | 35.8 | 27.5 | 0.2 | 0.85 | <0.01 | 0.55 |
| Total N excreted, g/d | 341.4 | 374.2 | 329.5 | 362.8 | 36.05 | 0.47 | <0.01 | 0.32 |
| N Balance, g/d | −2.9 | 15.9 | −1.9 | 16.4 | 53.5 | 0.71 | 0.63 | 0.96 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marshall, C.J.; Beck, M.R.; Garrett, K.; Barrell, G.K.; Al-Marashdeh, O.; Gregorini, P. Nitrogen Balance of Dairy Cows Divergent for Milk Urea Nitrogen Breeding Values Consuming Either Plantain or Perennial Ryegrass. Animals 2021, 11, 2464. https://doi.org/10.3390/ani11082464
Marshall CJ, Beck MR, Garrett K, Barrell GK, Al-Marashdeh O, Gregorini P. Nitrogen Balance of Dairy Cows Divergent for Milk Urea Nitrogen Breeding Values Consuming Either Plantain or Perennial Ryegrass. Animals. 2021; 11(8):2464. https://doi.org/10.3390/ani11082464
Chicago/Turabian StyleMarshall, Cameron J., Matthew R. Beck, Konagh Garrett, Graham K. Barrell, Omar Al-Marashdeh, and Pablo Gregorini. 2021. "Nitrogen Balance of Dairy Cows Divergent for Milk Urea Nitrogen Breeding Values Consuming Either Plantain or Perennial Ryegrass" Animals 11, no. 8: 2464. https://doi.org/10.3390/ani11082464
APA StyleMarshall, C. J., Beck, M. R., Garrett, K., Barrell, G. K., Al-Marashdeh, O., & Gregorini, P. (2021). Nitrogen Balance of Dairy Cows Divergent for Milk Urea Nitrogen Breeding Values Consuming Either Plantain or Perennial Ryegrass. Animals, 11(8), 2464. https://doi.org/10.3390/ani11082464