
Detection of 15-bp Deletion Mutation within PLAG1 Gene and Its Effects on Growth Traits in Goats

 ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Sample Collection and DNA Extraction
2.2. Primer Design, InDel Genotyping and DNA Sequencing
2.3. Statistical Analysis
3. Results
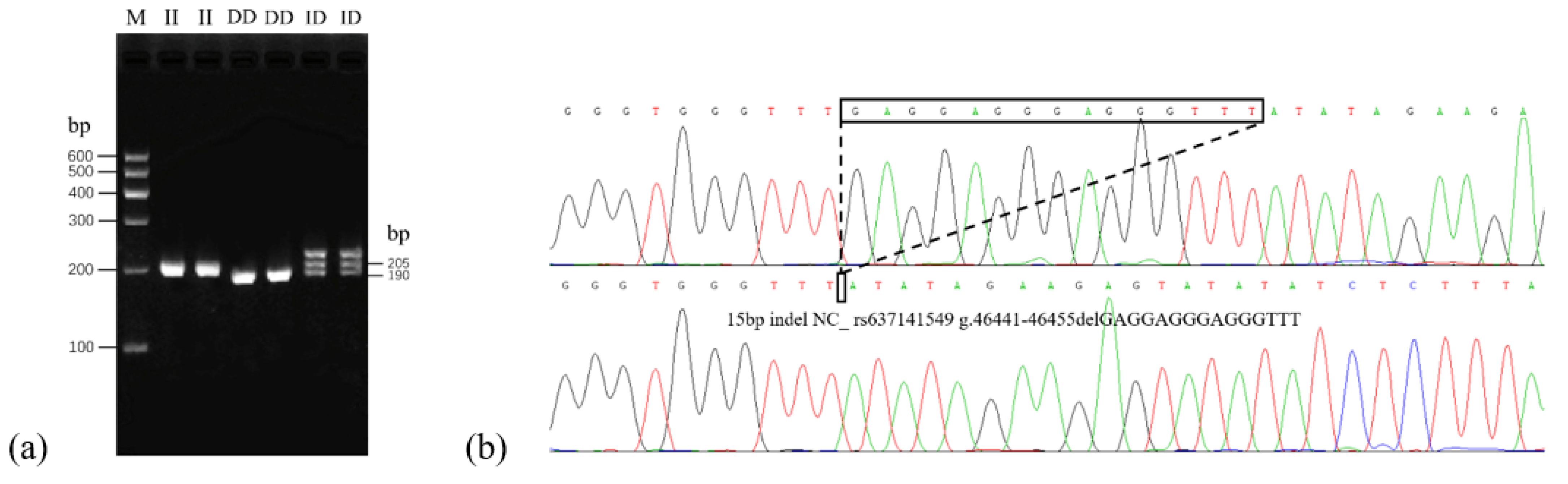
3.1. Detection and Genotyping of the PLAG1 Gene in Goat
3.2. Population Genetic Parameter Analysis
3.3. Relationship between PLAG1 InDel and Growth-Related Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guan, D.L.; Luo, N.J.; Tan, X.S.; Zhao, Z.Q.; Huang, Y.F.; Na, R.S.; Zhang, J.H.; Zhao, Y.J. Scanning of selection signature provides a glimpse into important economic traits in goats (Capra hircus). Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Wang, K.; Yan, H.L.; Xu, H.; Yang, Q.; Zhang, S.H.; Pan, C.Y.; Chen, H.; Zhu, H.J.; Liu, J.W.; Qu, L.; et al. A novel indel within goat casein alpha S1 gene is significantly associated with litter size. Gene 2018, 671, 161–169. [Google Scholar] [CrossRef]
- Dai, B.; Liang, H.; Guo, D.D.; Bi, Z.W.; Yuan, J.L.; Jin, Y.; Huan, L.; Guo, X.D.; Cang, M.; Liu, D.J. The Overexpression of Tβ4 in the Hair Follicle Tissue of Alpas Cashmere Goats Increases Cashmere Yield and Promotes Hair Follicle Development. Animals 2019, 10, 75. [Google Scholar] [CrossRef]
- Shaat, I.; Maki-Tanila, A. Variation in direct and maternal genetic effects for meat production traits in Egyptian Zaraibi goats. J. Anim. Breed. Genet. 2009, 126, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Collard, B.C.; Mackill, D.J. Marker-assisted selection: An approach for precision plant breeding in the twenty-first century. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2008, 363, 557–572. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, X.; Jiang, E.; Yan, H.; Zhu, H.; Chen, H.; Liu, J.; Qu, L.; Pan, C.; Lan, X. InDels within caprine IGF2BP1 intron 2 and the 3′-untranslated regions are associated with goat growth traits. Anim. Genet. 2020, 51, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Cui, Y.; Wang, Z.; Yan, H.L.; Meng, Z.; Zhu, H.J.; Qu, L.; Lan, X.Y.; Pan, C.Y. One 16 bp insertion/deletion (indel) within the KDM6A gene revealing strong associations with growth traits in goat. Gene 2019, 686, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Y.; Liu, D.L.; Tang, S.Q.; Li, D.H.; Han, R.L.; Tian, Y.D.; Li, H.; Li, G.X.; Li, W.T.; Liu, X.J.; et al. A multiallelic indel in the promoter region of the Cyclin-dependent kinase inhibitor 3 gene is significantly associated with body weight and carcass traits in chickens. Poult. Sci. 2019, 98, 556–565. [Google Scholar] [CrossRef]
- Ren, T.; Li, W.; Liu, D.; Liang, K.; Wang, X.; Li, H.; Jiang, R.; Tian, Y.; Kang, X.; Li, Z. Two insertion/deletion variants in the promoter region of the QPCTL gene are significantly associated with body weight and carcass traits in chickens. Anim. Genet. 2019, 50, 279–282. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, H.; Luo, Y.; Zhao, M.; Gong, H.; Hao, Z.; Hu, J.; Hickford, J.G.H. Variation in the Caprine KAP24-1 Gene Affects Cashmere Fibre Diameter. Animals 2019, 9, 15. [Google Scholar] [CrossRef]
- Kas, K.; Voz, M.L.; Röijer, E.; Aström, A.K.; Meyen, E.; Stenman, G.; Van de Ven, W.J. Promoter swapping between the genes for a novel zinc finger protein and beta-catenin in pleiomorphic adenomas with t(3;8)(p21;q12) translocations. Nat. Genet. 1997, 15, 170–174. [Google Scholar] [CrossRef]
- Van Dyck, F.; Declercq, J.; Braem, C.V.; Van de Ven, W.J. PLAG1, the prototype of the PLAG gene family: Versatility in tumour development (review). Int. J. Oncol. 2007, 30, 765–774. [Google Scholar] [CrossRef]
- Cho, Y.S.; Go, M.J.; Kim, Y.J.; Heo, J.Y.; Oh, J.H.; Ban, H.J.; Yoon, D.; Lee, M.H.; Kim, D.J.; Park, M.; et al. A large-scale genome-wide association study of Asian populations uncovers genetic factors influencing eight quantitative traits. Nat. Genet. 2009, 41, 527–534. [Google Scholar] [CrossRef]
- Kim, J.J.; Lee, H.I.; Park, T.; Kim, K.; Lee, J.E.; Cho, N.H.; Shin, C.; Cho, Y.S.; Lee, J.Y.; Han, B.G.; et al. Identification of 15 loci influencing height in a Korean population. J. Hum. Genet. 2010, 55, 27–31. [Google Scholar] [CrossRef]
- Okada, Y.; Kamatani, Y.; Takahashi, A.; Matsuda, K.; Hosono, N.; Ohmiya, H.; Daigo, Y.; Yamamoto, K.; Kubo, M.; Nakamura, Y.; et al. A genome-wide association study in 19 633 Japanese subjects identified LHX3-QSOX2 and IGF1 as adult height loci. Hum. Mol. Genet. 2010, 19, 2303–2312. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gudbjartsson, D.F.; Walters, G.B.; Thorleifsson, G.; Stefansson, H.; Halldorsson, B.V.; Zusmanovich, P.; Sulem, P.; Thorlacius, S.; Gylfason, A.; Steinberg, S.; et al. Many sequence variants affecting diversity of adult human height. Nat. Genet. 2008, 40, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Lettre, G.; Jackson, A.U.; Gieger, C.; Schumacher, F.R.; Berndt, S.I.; Sanna, S.; Eyheramendy, S.; Voight, B.F.; Butler, J.L.; Guiducci, C.; et al. Identification of ten loci associated with height highlights new biological pathways in human growth. Nat. Genet. 2008, 40, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Lango Allen, H.; Estrada, K.; Lettre, G.; Berndt, S.I.; Weedon, M.N.; Rivadeneira, F.; Willer, C.J.; Jackson, A.U.; Vedantam, S.; Raychaudhuri, S.; et al. Hundreds of variants clustered in genomic loci and biological pathways affect human height. Nature 2010, 467, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.R.; Esko, T.; Yang, J.; Vedantam, S.; Pers, T.H.; Gustafsson, S.; Chun, A.Y.; Estrada, K.; Luan, J.; Kutalik, Z.; et al. Defining the role of common variation in the genomic and biological architecture of adult human height. Nat. Genet. 2014, 46, 1173–1186. [Google Scholar] [CrossRef]
- Hensen, K.; Braem, C.; Declercq, J.; Van Dyck, F.; Dewerchin, M.; Fiette, L.; Denef, C.; Van de Ven, W.J. Targeted disruption of the murine PLAG1 proto-oncogene causes growth retardation and reduced fertility. Dev. Growth. Differ. 2004, 46, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Saatchi, M.; Schnabel, R.D.; Taylor, J.F.; Garrick, D.J. Large-effect pleiotropic or closely linked QTL segregate within and across ten US cattle breeds. BMC Genom. 2014, 15, 442. [Google Scholar] [CrossRef] [PubMed]
- Littlejohn, M.; Grala, T.; Sanders, K.; Walker, C.; Waghorn, G.; Macdonald, K.; Coppieters, W.; Georges, M.; Spelman, R.; Hillerton, E.; et al. Genetic variation in PLAG1 associates with early life body weight and peripubertal weight and growth in Bos taurus. Anim. Genet. 2012, 43, 591–594. [Google Scholar] [CrossRef] [PubMed]
- Fortes, M.R.; Kemper, K.; Sasazaki, S.; Reverter, A.; Pryce, J.E.; Barendse, W.; Bunch, R.; McCulloch, R.; Harrison, B.; Bolormaa, S.; et al. Evidence for pleiotropism and recent selection in the PLAG1 region in Australian Beef cattle. Anim. Genet. 2013, 44, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Bouwman, A.C.; Daetwyler, H.D.; Chamberlain, A.J.; Ponce, C.H.; Sargolzaei, M.; Schenkel, F.S.; Sahana, G.; Govignon-Gion, A.; Boitard, S.; Dolezal, M.; et al. Meta-analysis of genome-wide association studies for cattle stature identifies common genes that regulate body size in mammals. Nat. Genet. 2018, 50, 362. [Google Scholar] [CrossRef] [PubMed]
- Rubin, C.J.; Megens, H.J.; Martinez Barrio, A.; Maqbool, K.; Sayyab, S.; Schwochow, D.; Wang, C.; Carlborg, O.; Jern, P.; Jorgensen, C.B.; et al. Strong signatures of selection in the domestic pig genome. Proc. Natl. Acad. Sci. USA 2012, 109, 19529–19536. [Google Scholar] [CrossRef]
- Guo, Y.; Hou, L.; Zhang, X.; Huang, M.; Mao, H.; Chen, H.; Ma, J.; Chen, C.; Ai, H.; Ren, J.; et al. A meta analysis of genome-wide association studies for limb bone lengths in four pig populations. BMC Genet. 2015, 16, 95. [Google Scholar] [CrossRef][Green Version]
- Metzger, J.; Philipp, U.; Lopes, M.S.; Machado, A.D.; Felicetti, M.; Silvestrelli, M.; Distl, O. Analysis of copy number variants by three detection algorithms and their association with body size in horses. BMC Genom. 2013, 14. [Google Scholar] [CrossRef]
- Wu, H.; Pan, Y.; Zhang, Q.F.; Cao, Y.; Li, J.; Chen, H.; Cai, Y.; Sun, X.Z.; Lan, X.Y. Insertion/deletion (InDel) variations in sheep PLAG1 gene locating in growth-related major QTL are associated with adult body weight and morphometric traits. Small Rumin. Res. 2019, 178, 63–69. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, M.; Wu, H.; Akhatayeva, Z.; Lan, X.Y.; Fei, P.F.; Mao, C.; Jiang, F.G. Indel mutations of sheep PLAG1 gene and their associations with growth traits. Anim. Biotechnol. 2021. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, X.; Jia, W.; Pan, C.; Li, X.; Lei, C.; Chen, H.; Lan, X. Novel Nucleotide Variations, Haplotypes Structure and Associations with Growth Related Traits of Goat AT Motif-Binding Factor (ATBF1) Gene. Asian Australas. J. Anim. Sci. 2015, 28, 1394–1406. [Google Scholar] [CrossRef]
- Aljanabi, S.M.; Martinez, I. Universal and rapid salt-extraction of high quality genomic DNA for PCR-based techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wang, J.; Liu, N.; Cui, W.; Dong, W.; Xing, B.; Pan, C. Pig SOX9: Expression profiles of Sertoli cell (SCs) and a functional 18 bp indel affecting testis weight. Theriogenology 2019, 138, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, X.; Ma, L.; Xu, H.; Cao, X.; Luo, R.; Chen, H.; Sun, X.; Cai, Y.; Lan, X. Detection of a new 20-bp insertion/deletion (indel) within sheep PRND gene using mathematical expectation (ME) method. Prion 2017, 11, 143–150. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Shi, Y.Y.; He, L. SHEsis, a powerful software platform for analyses of linkage disequilibrium, haplotype construction, and genetic association at polymorphism loci. Cell Res. 2005, 15, 97–98. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Roychoudhury, A.K. Sampling variances of heterozygosity and genetic distance. Genetics 1974, 76, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, Q.; Wang, K.; Zhang, S.; Pan, C.; Chen, H.; Qu, L.; Yan, H.; Lan, X. A novel 12-bp indel polymorphism within the GDF9 gene is significantly associated with litter size and growth traits in goats. Anim. Genet. 2017, 48, 735–736. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Yan, H.; Li, J.; Xu, H.; Wang, K.; Zhu, H.; Chen, H.; Qu, L.; Lan, X. A novel 14-bp duplicated deletion within goat GHR gene is significantly associated with growth traits and litter size. Anim. Genet. 2017, 48, 499–500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Wang, K.; Liu, J.W.; Zhu, H.J.; Qu, L.; Chen, H.; Lan, X.Y.; Pan, C.Y.; Song, X.Y. An 11-bp Indel Polymorphism within the CSN1S1 Gene Is Associated with Milk Performance and Body Measurement Traits in Chinese Goats. Animals 2019, 9, 1114. [Google Scholar] [CrossRef]
- Bi, Y.; Feng, B.; Wang, Z.; Zhu, H.; Qu, L.; Lan, X.; Pan, C.; Song, X. Myostatin (MSTN) Gene Indel Variation and Its Associations with Body Traits in Shaanbei White Cashmere Goat. Animals 2020, 10, 168. [Google Scholar] [CrossRef]
- Gao, J.; Song, X.; Wu, H.; Tang, Q.; Wei, Z.; Wang, X.; Lan, X.; Zhang, B. Detection of rs665862918 (15-bp Indel) of the HIAT1 Gene and its Strong Genetic Effects on Growth Traits in Goats. Animals 2020, 10, 358. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Q.; Wang, K.; Yan, H.; Pan, C.; Chen, H.; Liu, J.; Zhu, H.; Qu, L.; Lan, X. Two strongly linked single nucleotide polymorphisms (Q320P and V397I) in GDF9 gene are associated with litter size in cashmere goats. Theriogenology 2019, 125, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Utsunomiya, Y.T.; Milanesi, M.; Utsunomiya, A.T.H.; Torrecilha, R.B.P.; Kim, E.S.; Costa, M.S.; Aguiar, T.S.; Schroeder, S.; do Carmo, A.S.; Carvalheiro, R.; et al. A PLAG1 mutation contributed to stature recovery in modern cattle. Sci. Rep. 2017, 7, 17140. [Google Scholar] [CrossRef]
- Minuto, F.; Palermo, C.; Arvigo, M.; Barreca, A.M. The IGF system and bone. J. Endocrinol. Investig. 2005, 28, 8–10. [Google Scholar]
- Wilson, E.M.; Rotwein, P. Control of MyoD function during initiation of muscle differentiation by an autocrine signaling pathway activated by insulin-like growth factor-II. J. Biol. Chem. 2006, 281, 29962–29971. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.K.; Westwood, M. Biology and significance of signalling pathways activated by IGF-II. Growth Factors 2012, 30, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Voz, M.L.; Agten, N.S.; Van de Ven, W.J.; Kas, K. PLAG1, the main translocation target in pleomorphic adenoma of the salivary glands, is a positive regulator of IGF-II. Cancer Res. 2000, 60, 106–113. [Google Scholar] [PubMed]
- Van Laere, A.S.; Nguyen, M.; Braunschweig, M.; Nezer, C.; Collette, C.; Moreau, L.; Archibald, A.L.; Haley, C.S.; Buys, N.; Tally, M.; et al. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature 2003, 425, 832–836. [Google Scholar] [CrossRef]
- Guan, B.; Gao, M.; Wu, C.H.; Wang, T.L.; Shih, I.M. Functional Analysis of In-frame Indel ARID1A Mutations Reveals New Regulatory Mechanisms of Its Tumor Suppressor Functions. Neoplasia 2012, 14, 986–993. [Google Scholar] [CrossRef]
- Minor, E.A.; Court, B.L.; Dubovy, S.; Wang, G.F. AMD-Associated Variants at the Chromosome 10q26 Locus and the Stability of ARMS2 Transcripts. Investig. Ophth. Vis. Sci. 2013, 54, 5913–5919. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; An, X.; Song, Y.; Gao, T.; Lei, Y.; Cao, B. Two Mutations in the Caprine MTHFR 3’UTR Regulated by MicroRNAs Are Associated with Milk Production Traits. PLoS ONE 2015, 10, e0133015. [Google Scholar] [CrossRef] [PubMed]
- Declercq, J.; Van Dyck, F.; Van Damme, B.; Van de Ven, W.J. Upregulation of Igf and Wnt signalling associated genes in pleomorphic adenomas of the salivary glands in PLAG1 transgenic mice. Int. J. Oncol. 2008, 32, 1041–1047. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xu, H.; Li, H.; Wang, Z.; Abudureyimu, A.; Yang, J.; Cao, X.; Lan, X.; Zang, R.; Cai, Y. A Deletion Downstream of the CHCHD7 Gene Is Associated with Growth Traits in Sheep. Animals 2020, 10, 1472. [Google Scholar] [CrossRef]
- Li, H.; Wang, X.; Chen, H.; Qu, L.; Lan, X. A 17-bp InDel (rs668420586) within goat CHCHD7 gene located in growth-related QTL affecting body measurement traits. 3 Biotech 2020, 10, 441. [Google Scholar] [CrossRef] [PubMed]
- Karim, L.; Takeda, H.; Lin, L.; Druet, T.; Arias, J.A.; Baurain, D.; Cambisano, N.; Davis, S.R.; Farnir, F.; Grisart, B.; et al. Variants modulating the expression of a chromosome domain encompassing PLAG1 influence bovine stature. Nat. Genet. 2011, 43, 405–413. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Breeds | Growth Traits | Effective Sample Size | Mean (cm) | Standard Error | Minimum (cm) | Maximum (cm) |
|---|---|---|---|---|---|---|
| HNBG | BH | 205 | 52.80 | 0.28 | 43.20 | 63.20 |
| BL | 205 | 56.15 | 0.31 | 46.00 | 68.30 | |
| CC | 205 | 72.38 | 0.46 | 54.00 | 91.00 | |
| CD | 205 | 26.56 | 0.16 | 19.20 | 33.00 | |
| CW | 205 | 14.91 | 0.13 | 10.80 | 21.60 | |
| HW | 205 | 13.71 | 0.94 | 10.20 | 17.20 | |
| CCB | 204 | 7.80 | 0.51 | 6.00 | 10.00 | |
| GZDG | BH | 80 | 73.53 | 0.48 | 63.00 | 83.00 |
| BL | 80 | 74.81 | 0.49 | 65.00 | 86.00 | |
| HAH | 80 | 74.08 | 0.47 | 64.00 | 84.00 | |
| CC | 26 | 87.50 | 0.48 | 83.00 | 92.00 | |
| CW | 26 | 16.69 | 0.09 | 16.00 | 17.00 | |
| CCB | 26 | 8.00 | 0.00 | 8.00 | 8.00 | |
| SWCG | BW | 649 | 48.91 | 0.61 | 15.00 | 79.00 |
| BH | 812 | 56.02 | 0.18 | 36.50 | 73.00 | |
| BL | 634 | 66.05 | 0.24 | 46.00 | 83.00 | |
| HAH | 811 | 58.94 | 0.17 | 23.20 | 73.00 | |
| CC | 814 | 84.61 | 0.30 | 62.00 | 114.00 | |
| CD | 813 | 29.00 | 0.11 | 14.70 | 42.00 | |
| CW | 814 | 19.91 | 0.13 | 11.50 | 34.00 | |
| HW | 472 | 15.40 | 0.09 | 10.00 | 21.00 | |
| CCB | 725 | 8.31 | 0.09 | 5.60 | 11.00 | |
| IMWC | BW | 452 | 24.13 | 0.36 | 11.00 | 54.80 |
| BH | 450 | 51.44 | 0.20 | 39.50 | 65.00 | |
| BL | 451 | 56.53 | 0.31 | 41.00 | 75.00 | |
| HAH | 450 | 53.29 | 0.23 | 43.50 | 68.00 | |
| CC | 450 | 69.61 | 0.37 | 55.50 | 97.00 | |
| CD | 451 | 24.27 | 0.16 | 16.00 | 38.50 | |
| CW | 451 | 14.74 | 0.13 | 9.00 | 24.00 | |
| HW | 450 | 12.97 | 0.12 | 8.50 | 53.50 | |
| CCB | 450 | 8.43 | 0.03 | 7.00 | 10.00 |
| Primer | Primer sequences (from 5’ to 3’) | Length | Region | Tm* |
|---|---|---|---|---|
| P1 | F: GGGGCGTCAACCTAAGAAACAT R: GAAATCCGCAGTGATAGCTGG | 210 | Intron 1 | TD |
| P2 | F: ATGAGTTACTGGTAAGGAGA R: AGAAGGCAATGGCAC | 209 | Intron 1 | TD |
| P3 | F: GAGATTCCTGTTAGAATTTATAGT R: CTGAAAAGCCCCGTACT | 132 | Intron 1 | TD |
| P4 | F: TGAGCAACAGGGAGGGTA R: TGGTGGCTACATCCAAGC | 205 | Downstream | 62 °C |
| Breeds | Size | Genotypic Frequencies | Allelic Frequencies | HWE | Population Parameters | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | II | ID | DD | I | D | p Values | Ho | He | Ne | PIC | |
| SWCG | 826 | 0.805 | 0.190 | 0.005 | 0.900 | 0.100 | p < 0.05 | 0.820 | 0.180 | 1.219 | 0.163 |
| IMWC | 452 | 0.695 | 0.281 | 0.024 | 0.835 | 0.165 | p < 0.05 | 0.725 | 0.275 | 1.380 | 0.237 |
| HNBG | 212 | 0.439 | 0.486 | 0.075 | 0.682 | 0.318 | p < 0.05 | 0.566 | 0.434 | 1.767 | 0.340 |
| GZDG | 91 | 0.944 | 0.056 | 0.000 | 0.973 | 0.027 | p < 0.05 | 0.947 | 0.053 | 1.056 | 0.052 |
| Breeds | Growth Traits | Genotypes | p Value | ||
|---|---|---|---|---|---|
| II | ID | DD | |||
| SWCG (n = 683) | BW (kg) | 53.23 ± 0.73 A (n = 436) | 46.79 ± 1.39 B (n = 120) | — | 0.0006 |
| BH (cm) | 57.01 ± 0.20 a (n = 544) | 55.96 ± 0.46 B (n = 136) | — | 0.022 | |
| BL (cm) | 67.10 ± 0.30 b (n = 392) | 66.54 ± 0.53 a (n = 111) | — | 0.369 | |
| HAH (cm) | 19.87 ± 0.17 (n = 545) | 20.32 ± 0.34 (n = 137) | — | 0.223 | |
| CC (cm) | 59.83 ± 0.18 a (n = 542) | 58.69 ± 0.49 b (n = 137) | — | 0.033 | |
| CD (cm) | 15.45 ± 0.12 (n = 285) | 15.45 ± 0.21 (n = 95) | — | 0.982 | |
| CW (cm) | 8.34 ± 0.12 (n = 545) | 8.31 ± 0.07 (n = 138) | — | 0.899 | |
| HW (cm) | 85.08 ± 0.36 (n = 544) | 83.87 ± 0.74 (n = 138) | — | 0.135 | |
| CCB (cm) | 29.20 ± 0.13 (n = 544) | 29.2 ± 0.23 (n = 137) | — | 0.862 | |
| HNBG (n = 205) | BH (cm) | 52.84 ± 0.43 (n = 92) | 52.87 ± 0.41 (n = 98) | 52.05 ± 1.11 (n = 15) | 0.749 |
| BL (cm) | 55.96 ± 0.44 (n = 92) | 56.51 ± 0.48 (n = 98) | 55.05 ± 1.10 (n = 15) | 0.383 | |
| CC (cm) | 72.88 ± 0.62 (n = 92) | 72.01 ± 0.69 (n = 98) | 71.80 ± 2.08 (n = 15) | 0.652 | |
| CD (cm) | 26.71 ± 0.21 (n = 92) | 26.37 ± 0.25 (n = 98) | 26.85 ± 0.16 (n = 15) | 0.567 | |
| CW (cm) | 15.09 ± 0.17 (n = 92) | 14.7 ± 0.21 (n = 98) | 14.9 ± 0.13 (n = 15) | 0.517 | |
| HW (cm) | 13.78 ± 0.14 (n = 92) | 13.69 ± 0.13 (n = 98) | 13.5 ± 0.46 (n = 15) | 0.670 | |
| CCB (cm) | 7.82 ± 0.74 (n = 92) | 7.76 ± 0.73 (n = 97) | 7.90 ± 0.24 (n = 15) | 0.722 | |
| BI | 130.68 ± 1.07 a (n = 92) | 127.52 ± 0.71 b (n = 98) | 130.36 ± 2.59 a (n = 15) | 0.044 | |
| BLI | 106.22 ± 0.84 (n = 92) | 107.09 ± 0.77 (n = 98) | 106.19 ± 0.55 (n = 15) | 0.687 | |
| ChCI | 138.19 ± 1.00 (n = 92) | 136.33 ± 0.98 (n = 98) | 138.15 ± 3.50 (n = 15) | 0.412 | |
| GZDG (n = 80) | BH (cm) | 73.35 ± 0.51 (n = 75) | 76.2 ± 1.07 (n = 5) | -- | 0.147 |
| BL (cm) | 74.63 ± 0.51 (n = 75) | 77.60 ± 1.40 (n = 5) | -- | 0.132 | |
| HAH (cm) | 73.92 ± 0.49 (n = 75) | 76.40 ± 1.36 (n = 5) | -- | 0.197 | |
| HW (cm) | 16.65 ± 0.10 B (n = 23) | 17.00 ± 0.00 A (n = 3) | -- | 0.002 | |
| CC (cm) | 87.35 ± 0.51 (n = 23) | 88.67 ± 1.67 (n = 3) | -- | 0.395 | |
| CCB (cm) | 8.00 ± 0.00 (n = 23) | 8.00 ± 0.00 (n = 3) | -- | 0.000 | |
| IMWC (n = 68) | BW (kg) | 34.30 ± 1.06 (n = 47) | 36.74 ± 1.66 (n = 21) | -- | 0.211 |
| BH (cm) | 56.86 ± 0.51 (n = 47) | 55.81 ± 0.76 (n = 21) | -- | 0.255 | |
| BL (cm) | 66.67 ± 0.62 (n = 47) | 66.40 ± 0.80 (n = 21) | -- | 0.805 | |
| HAH (cm) | 59.86 ± 0.56 (n = 47) | 60.35 ± 0.77 (n = 21) | -- | 0.615 | |
| CC (cm) | 78.48 ± 0.94 (n = 47) | 81.02 ± 1.48 (n = 21) | -- | 0.142 | |
| CD (cm) | 29.52 ± 0.33 (n = 47) | 20.40 ± 0.44 (n = 21) | -- | 0.841 | |
| CW (cm) | 18.62 ± 0.37 (n = 47) | 19.21 ± 0.52 (n = 21) | -- | 0.361 | |
| HW (cm) | 15.97 ± 0.83 (n = 47) | 15.36 ± 0.23 (n = 21) | -- | 0.611 | |
| CCB (cm) | 8.38 ± 0.68 (n = 47) | 8.57 ± 0.11 (n = 21) | -- | 0.140 | |
| Breeds | Growth Traits | Genotypes | p Value | ||
|---|---|---|---|---|---|
| II | ID | DD | |||
| SWCG (n = 89) | BW (kg) | 35.67 ± 0.90 (n = 73) | 32.91 ± 1.74 (n = 16) | — | 0.187 |
| BH (cm) | 49.99 ± 0.44 (n = 73) | 48.89 ± 1.10 (n = 16) | — | 0.305 | |
| BL (cm) | 62.90±0.62 (n=73) | 62.29 ± 1.47 (n = 16) | — | 0.681 | |
| HAH (cm) | 54.03 ± 0.50 a (n = 73) | 51.46 ± 1.26 b (n = 16) | — | 0.036 | |
| CC (cm) | 79.85 ± 0.84 (n = 73) | 76.81 ± 1.85 (n = 16) | — | 0.130 | |
| CD (cm) | 28.19 ± 0.29 (n = 73) | 27.45 ± 1.05 (n = 16) | — | 0.345 | |
| CW (cm) | 20.57 ± 0.32 (n = 73) | 20.17 ± 0.75 (n = 16) | — | 0.602 | |
| HW (cm) | 15.20 ± 0.20 (n = 73) | 14.88 ± 1.69 (n = 16) | — | 0.493 | |
| CCB (cm) | 8.01 ± 0.10 (n = 73) | 7.62 ± 0.24 (n = 16) | — | 0.122 | |
| IMWC (n = 373) | BW (kg) | 21.79 ± 0.34 (n = 267) | 22.94 ± 0.59 (n = 106) | — | 0.076 |
| BH (cm) | 50.69 ± 0.23 (n = 265) | 50.18 ± 0.40 (n = 106) | — | 0.245 | |
| BL (cm) | 54.56 ± 0.33 (n = 266) | 55.18 ± 0.49 (n = 106) | — | 0.304 | |
| HAH (cm) | 52.19 ± 0.24 (n = 265) | 52.01 ± 0.40 (n = 106) | — | 0.692 | |
| CC (cm) | 67.46 ± 0.39 (n = 265) | 68.71 ± 0.63 (n = 106) | — | 0.089 | |
| CD (cm) | 23.30 ± 0.17 (n = 266) | 23.37 ± 0.25 (n = 106) | — | 0.828 | |
| CW (cm) | 13.90 ± 0.13 (n = 266) | 14.30 ± 0.21 (n = 106) | — | 0.105 | |
| HW (cm) | 12.34 ± 0.10 B(n = 266) | 12.86 ± 0.15 A (n = 105) | — | 0.005 | |
| CCB (cm) | 8.43 ± 0.03 (n = 265) | 8.42 ± 0.06 (n = 106) | — | 0.977 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, Z.; Wang, K.; Wu, H.; Wang, Z.; Pan, C.; Chen, H.; Lan, X. Detection of 15-bp Deletion Mutation within PLAG1 Gene and Its Effects on Growth Traits in Goats. Animals 2021, 11, 2064. https://doi.org/10.3390/ani11072064
Wei Z, Wang K, Wu H, Wang Z, Pan C, Chen H, Lan X. Detection of 15-bp Deletion Mutation within PLAG1 Gene and Its Effects on Growth Traits in Goats. Animals. 2021; 11(7):2064. https://doi.org/10.3390/ani11072064
Chicago/Turabian StyleWei, Zhenyu, Ke Wang, Hui Wu, Zhen Wang, Chuanying Pan, Hong Chen, and Xianyong Lan. 2021. "Detection of 15-bp Deletion Mutation within PLAG1 Gene and Its Effects on Growth Traits in Goats" Animals 11, no. 7: 2064. https://doi.org/10.3390/ani11072064
APA StyleWei, Z., Wang, K., Wu, H., Wang, Z., Pan, C., Chen, H., & Lan, X. (2021). Detection of 15-bp Deletion Mutation within PLAG1 Gene and Its Effects on Growth Traits in Goats. Animals, 11(7), 2064. https://doi.org/10.3390/ani11072064