
Effect of Lipopolysaccharides (LPS) and Lipoteichoic Acid (LTA) on the Inflammatory Response in Rumen Epithelial Cells (REC) and the Impact of LPS on Claw Explants

Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Characterization of Primary Rumen Epithelial Cells (REC)
2.1.1. Isolation and Cultivation of Rumen Epithelial Cells
2.1.2. Characterization of REC with Immunocytochemistry
2.2. Evaluation of the Effects of LPS and LTA on Gene Expression
2.2.1. Stimulation of REC with LPS and LTA
2.2.2. RNA Extraction and Gene Expression Analysis of REC
2.3. Effect of LPS on Separation Force and Gene Expression of Bovine Claw Explants
2.3.1. Preparation of Bovine Claw Explants with LPS
2.3.2. Metabolic Activity and Separation Force of Bovine Claw Explants
2.3.3. RNA Extraction and Gene Expression Analysis
2.4. Statistics
3. Results
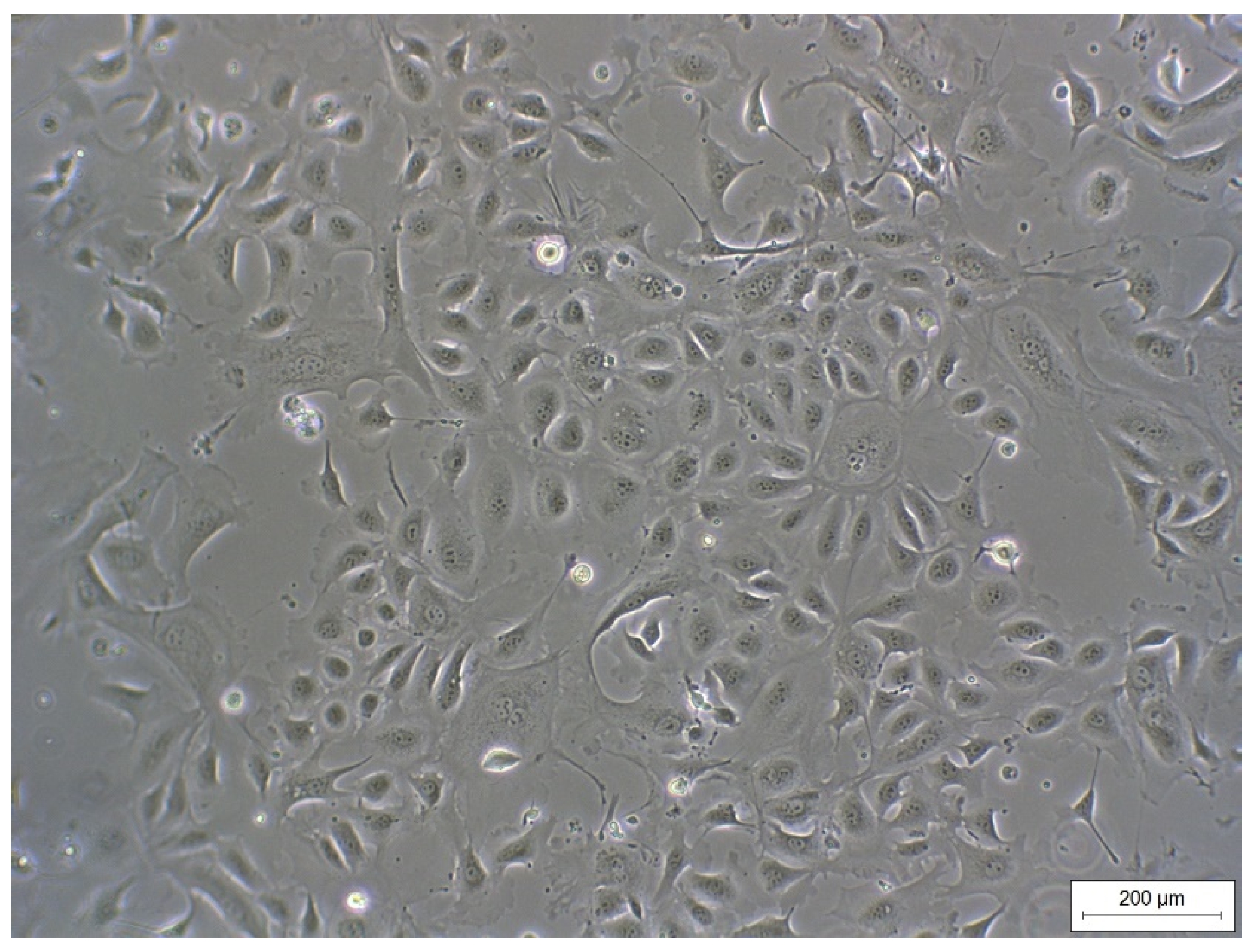
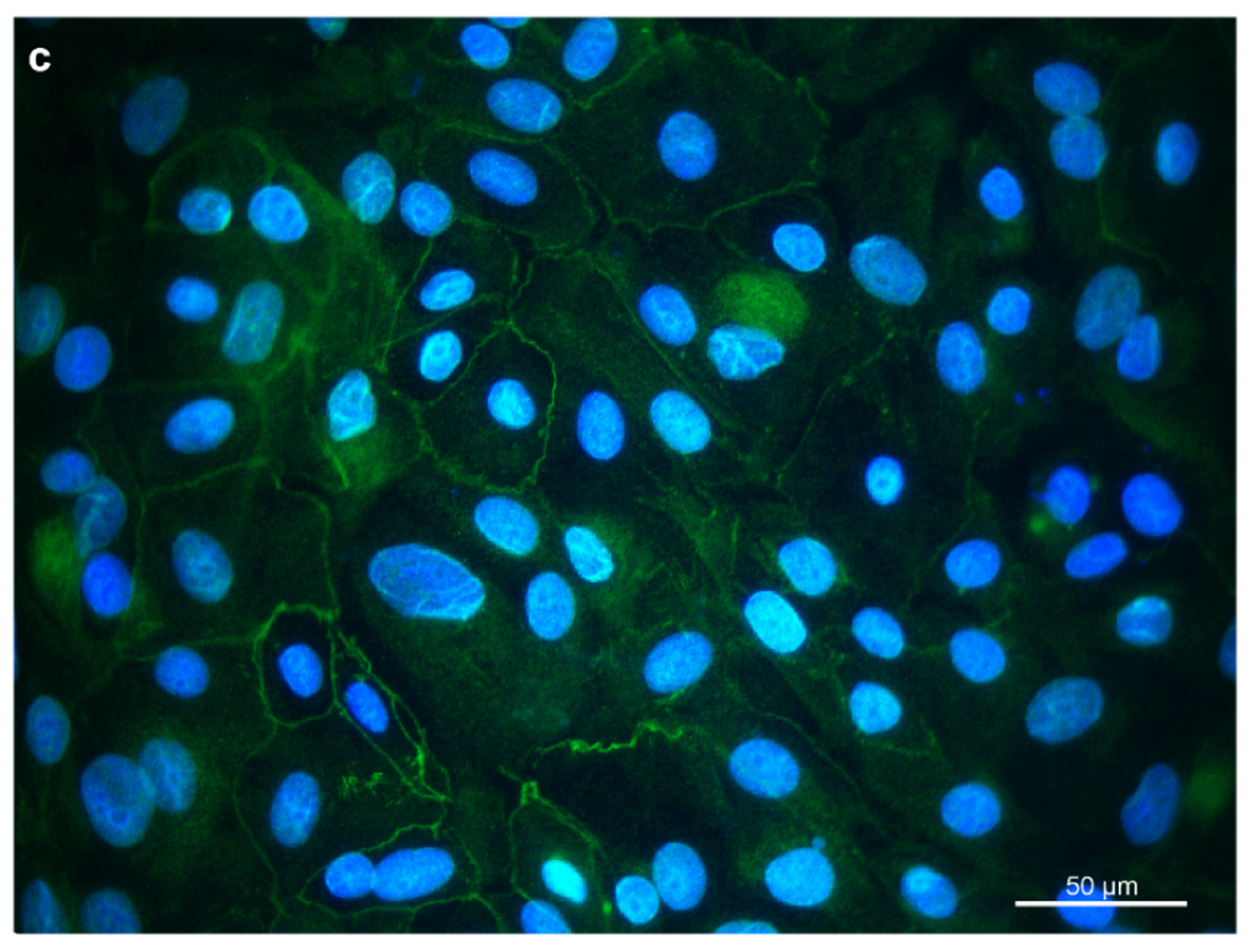
3.1. Isolation and Characterization of Primary Rumen Epithelial Cells
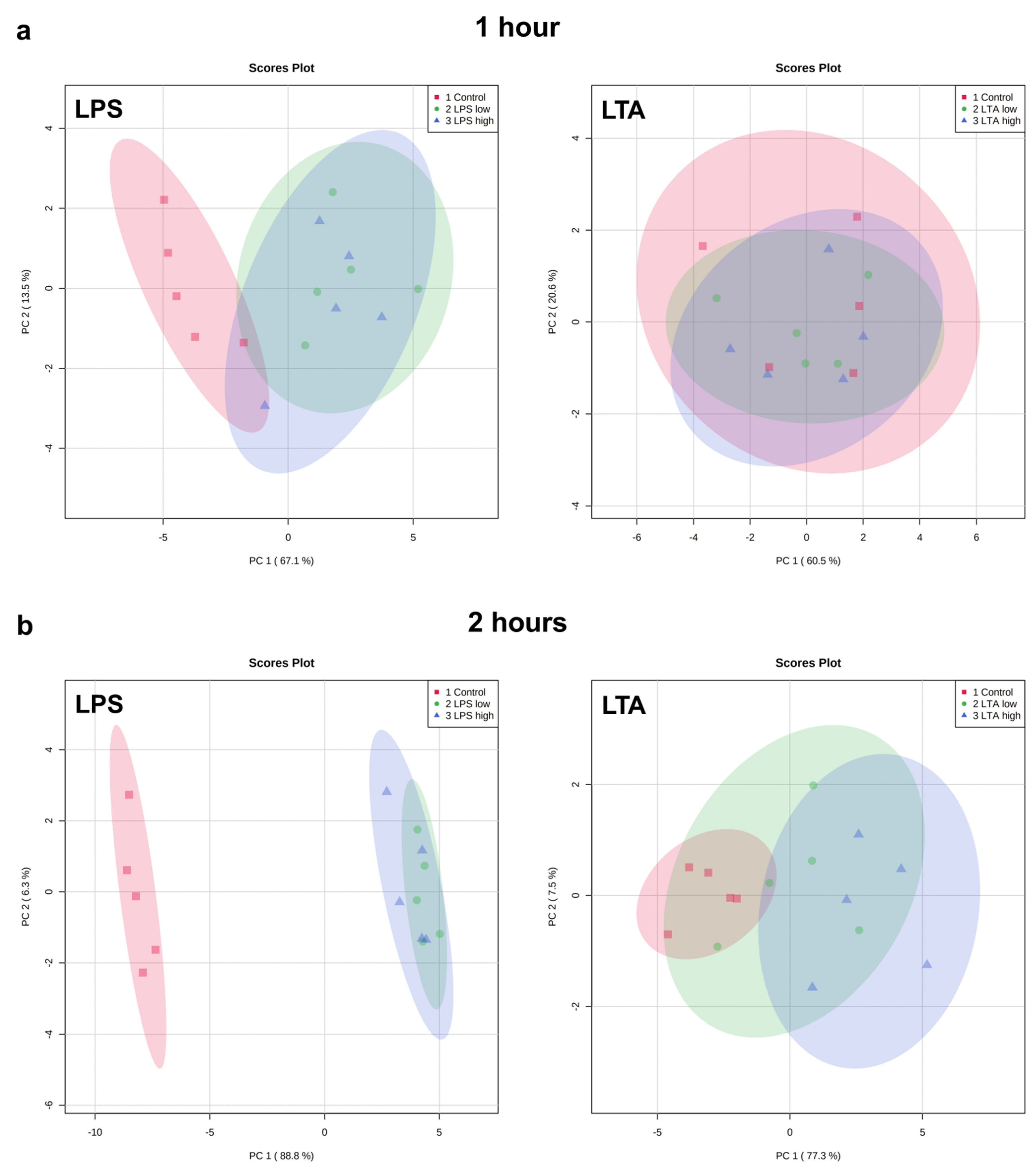
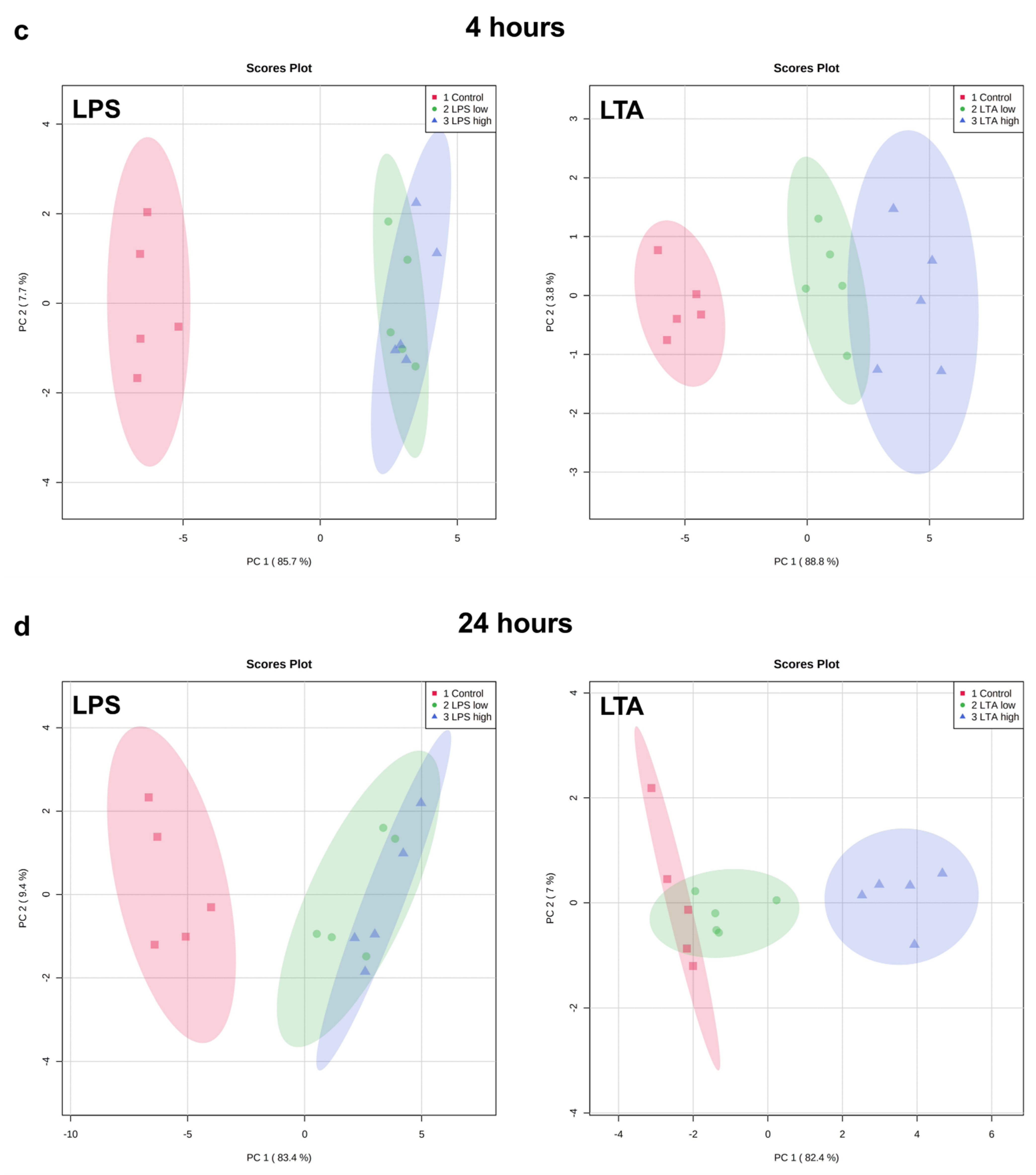
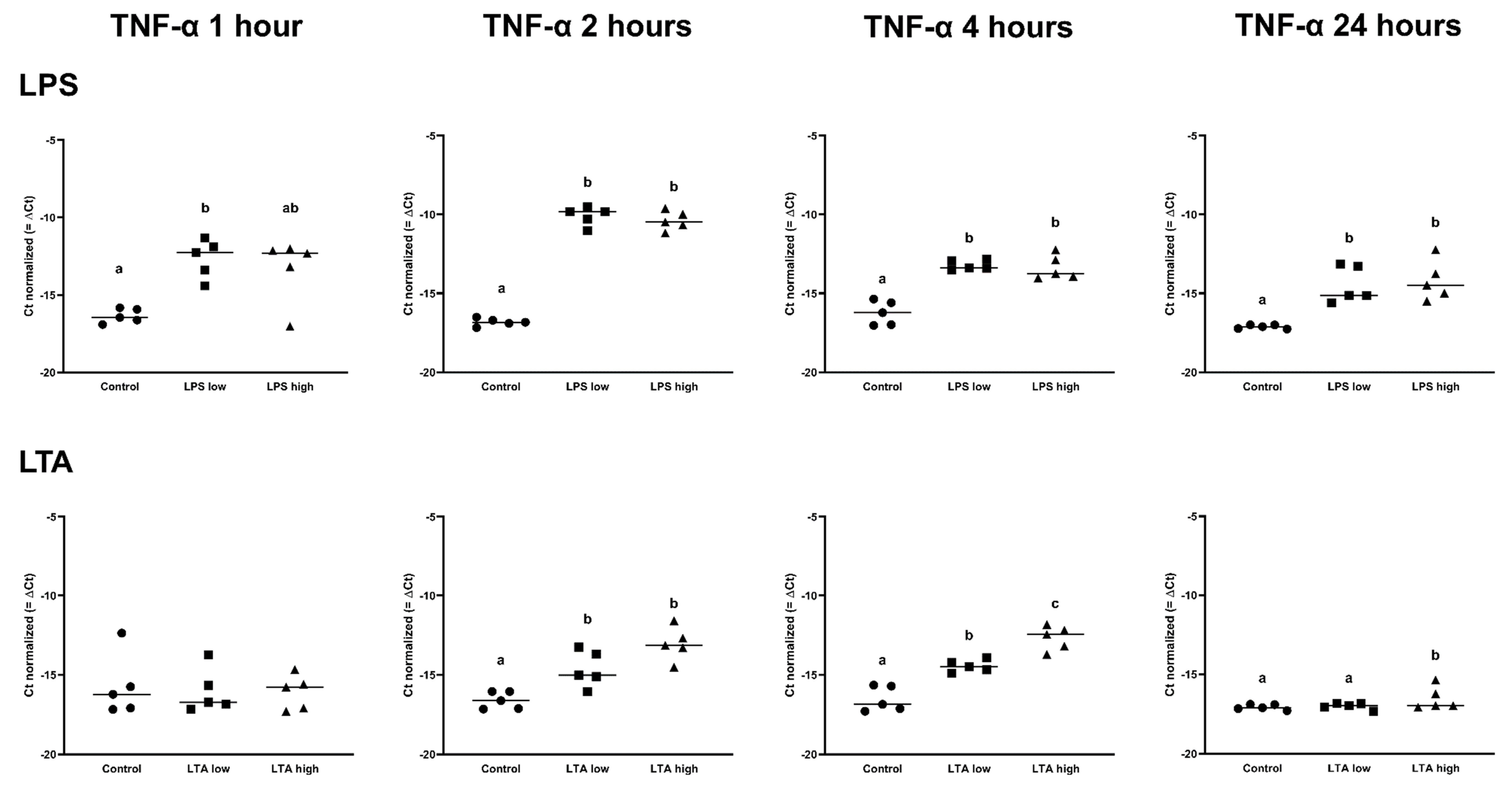
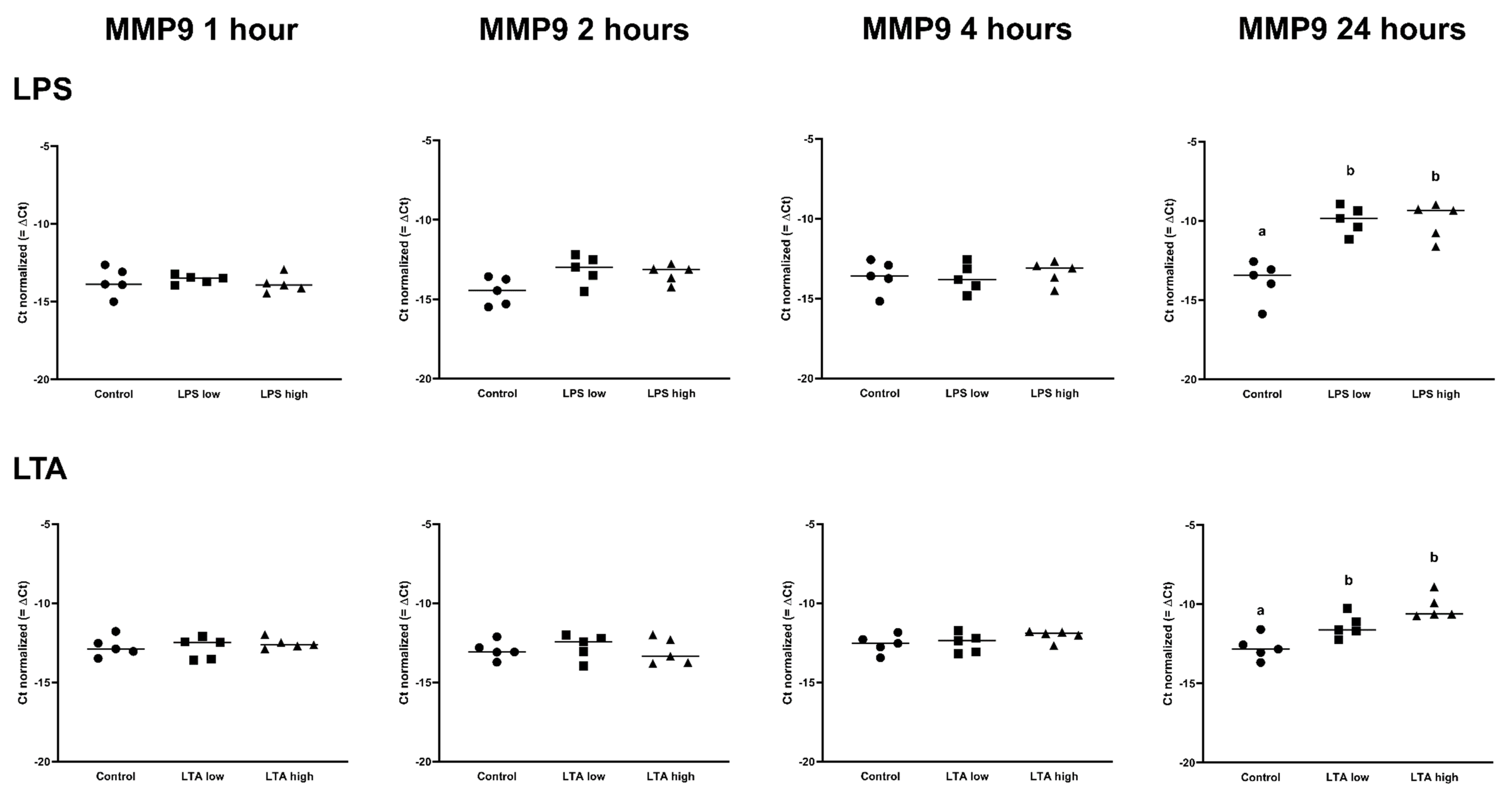
3.2. Effects of LPS and LTA on Gene Expression in Primary Rumen Epithelial Cells
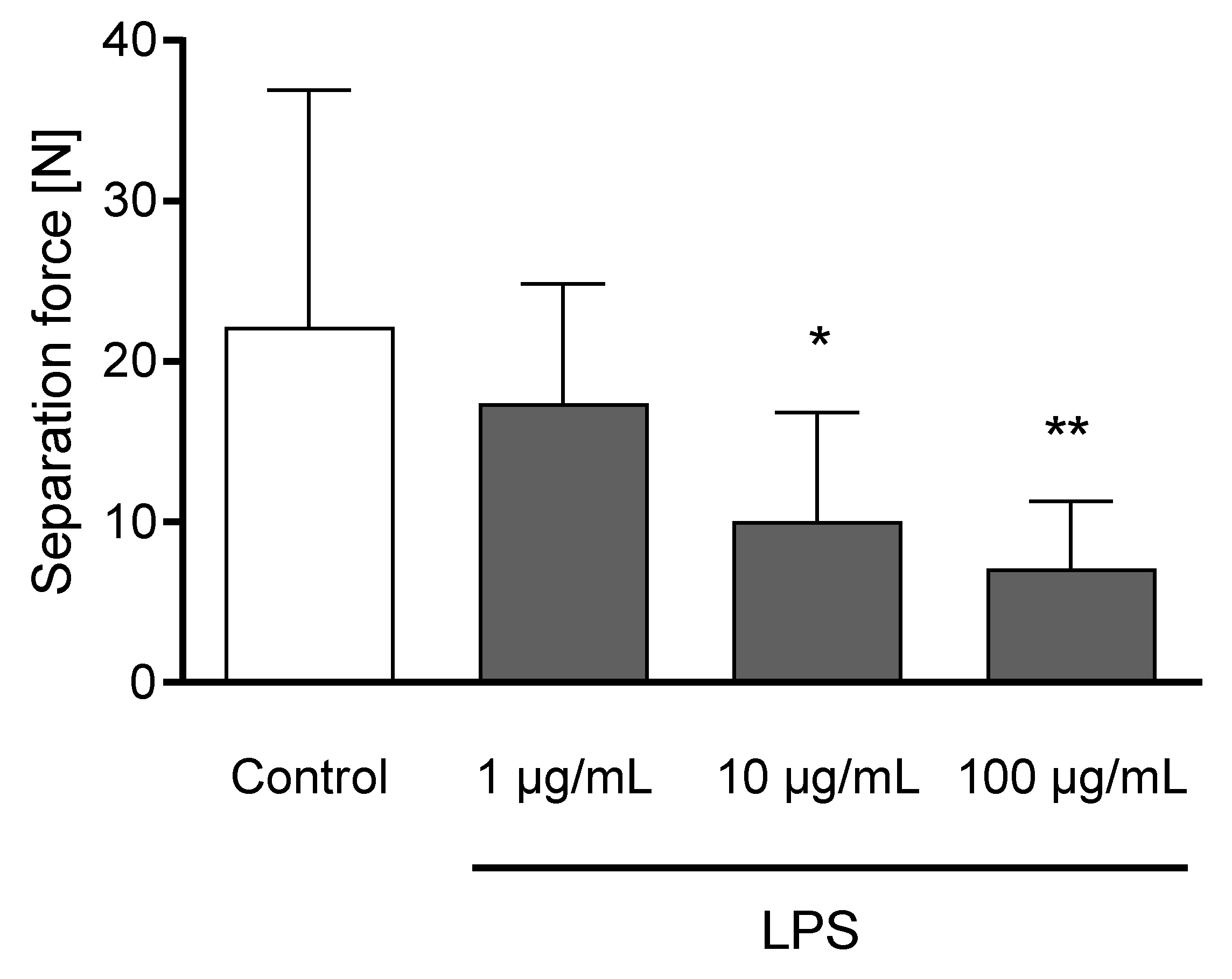
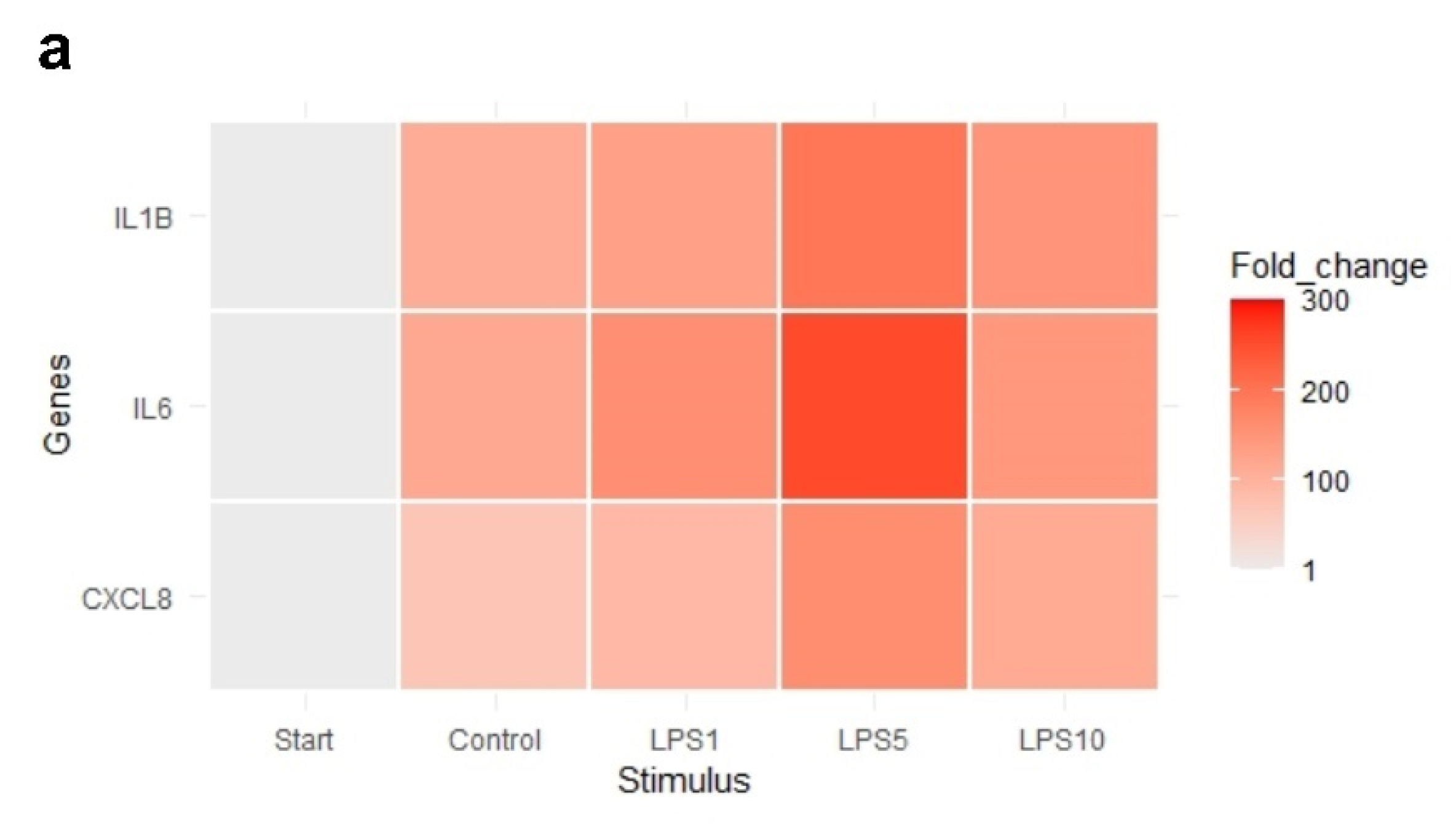
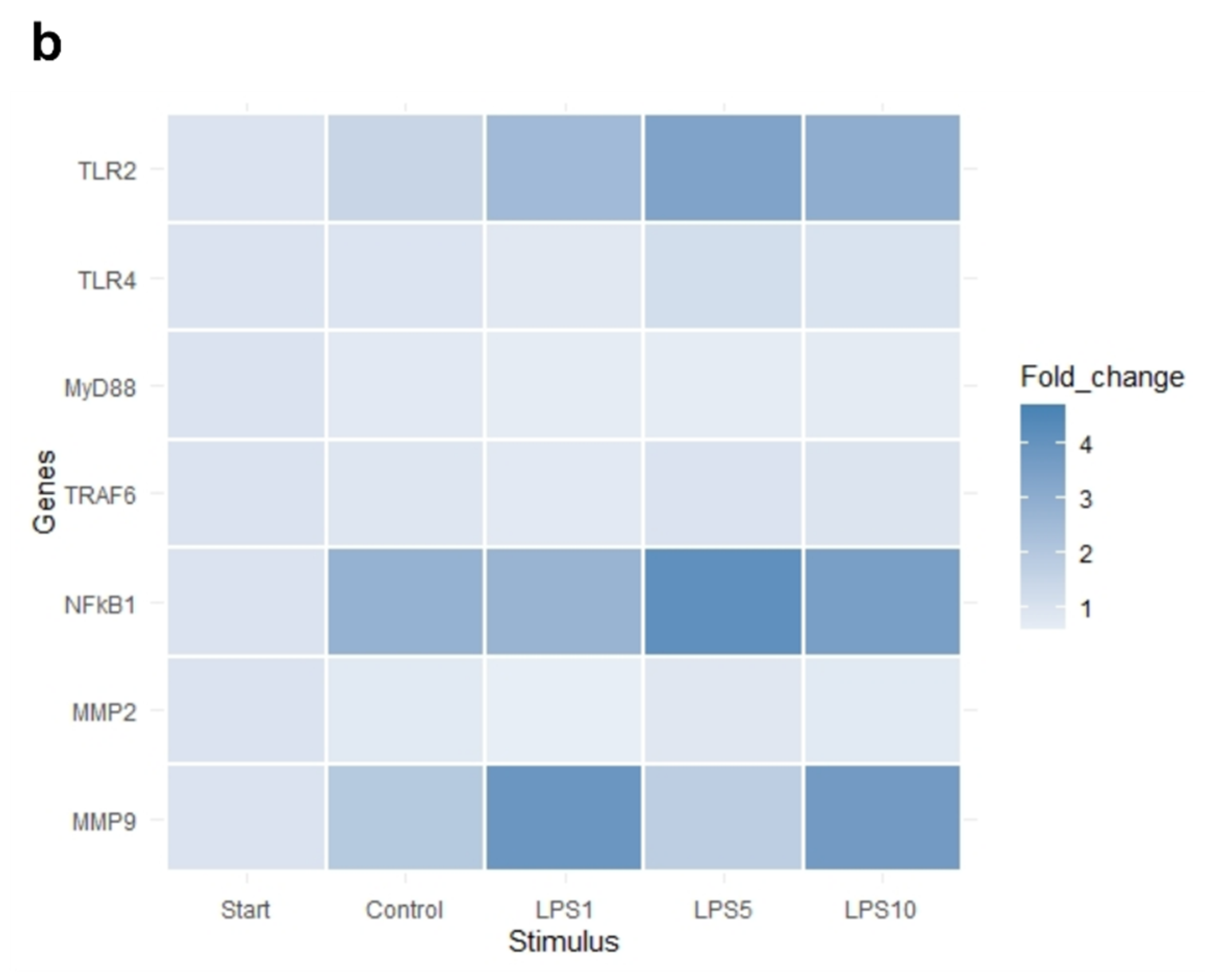
3.3. Effects of LPS on Tissue Integrity and Gene Expression of Bovine Claw Explants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Monteiro, H.F.; Faciola, A.P. Ruminal acidosis, bacterial changes, and lipopolysaccharides. J. Anim. Sci. 2020, 98, skaa248. [Google Scholar] [CrossRef]
- Eckel, E.F.; Ametaj, B.N. Invited review: Role of bacterial endotoxins in the etiopathogenesis of periparturient diseases of transition dairy cows. J. Dairy Sci. 2016, 99, 5967–5990. [Google Scholar] [CrossRef] [PubMed]
- Ametaj, B.N.; Zebeli, Q.; Iqbal, S. Nutrition, microbiota, and endotoxin-related diseases in dairy cows. Rev. Bras. Zootec. 2010, 39, 433–444. [Google Scholar] [CrossRef]
- Khiaosa-Ard, R.; Zebeli, Q. Diet-induced inflammation: From gut to metabolic organs and the consequences for the health and longevity of ruminants. Res. Vet. Sci. 2018, 120, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Kvidera, S.K.; Horst, E.A.; Abuajamieh, M.; Mayorga, E.J.; Fernandez, M.V.S.; Baumgard, L.H. Glucose requirements of an activated immune system in lactating Holstein cows. J. Dairy Sci. 2017, 100, 2360–2374. [Google Scholar] [CrossRef]
- Humer, E.; Kröger, I.; Neubauer, V.; Schedle, K.; Reisinger, N.; Zebeli, Q. Supplementing phytogenic compounds or autolyzed yeast modulates ruminal biogenic amines and plasma metabolome in dry cows experiencing subacute ruminal acidosis. J. Dairy Sci. 2018, 101, 9559–9574. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Zebeli, Q.; Mansmann, D.A.; Dunn, S.M.; Ametaj, B. Oral administration of LPS and lipoteichoic acid prepartum modulated reactants of innate and humoral immunity in periparturient dairy cows. Innate Immun. 2014, 20, 390–400. [Google Scholar] [CrossRef]
- Ametaj, B.; Sivaraman, S.; Dunn, S.; Zebeli, Q. Repeated oral administration of lipopolysaccharide from Escherichia coli 0111:B4 modulated humoral immune responses in periparturient dairy cows. Innate Immun. 2012, 18, 638–647. [Google Scholar] [CrossRef]
- Samarasinghe, M.B.; Larsen, T. Oral administration of lipopolysaccharides from Escherichia coli (serotype O111/: B4) does not induce an effective systemic immune response in milk-fed Holstein calves. J. dairy Sci. 2020, 103, 5525–5531. [Google Scholar] [CrossRef]
- Li, S.; Khafipour, E.; Krause, D.O.; Kroeker, A.; Gozho, G.N.; Plaizier, J.C. Effects of subacute ruminal acidosis challenges on fermentation and endotoxins in the rumen and hindgut of dairy cows. J. Dairy Sci. 2012, 95, 294–303. [Google Scholar] [CrossRef]
- Reisinger, N.; Schürer-Waldheim, S.; Mayer, E.; Debevere, S.; Antonissen, G.; Sulyok, M.; Nagl, V. Mycotoxin Occurrence in Maize Silage—A Neglected Risk for Bovine Gut Health? Toxins 2019, 11, 577. [Google Scholar] [CrossRef] [PubMed]
- Katwal, P.; Uprety, T.; Okda, F.; Antony, L.; Thomas, M.; Chase, C.; Diel, D.G.; Nelson, E.; Young, A.; Li, F. Characterization of bovine ileal epithelial cell line for lectin binding, susceptibility to enteric pathogens, and TLR mediated immune responses. Comp. Immunol. Microbiol. Infect. Dis. 2021, 74, 101581. [Google Scholar] [CrossRef] [PubMed]
- Kent-Dennis, C.; Aschenbach, J.R.; Griebel, P.J.; Penner, G.B. Effects of lipopolysaccharide exposure in primary bovine ruminal epithelial cells. J. Dairy Sci. 2020, 103, 9587–9603. [Google Scholar] [CrossRef]
- Kent-Dennis, C.; Penner, G.B. Effects of a proinflammatory response on metabolic function of cultured, primary ruminal epithelial cells. J. Dairy Sci. 2021, 104, 1002–1017. [Google Scholar] [CrossRef]
- Klotz, J.L.; Baldwin, R.L.; Gillis, R.C.; Heitmann, R.N. Refinements in Primary Rumen Epithelial Cell Incubation Techniques 1. J. Dairy Sci. 2001, 84, 183–193. [Google Scholar] [CrossRef]
- Andersen, P.H.; Jarlov, N.; Hesselholt, M.; Baek, L. Studies on in vivo Endotoxin Plasma Disappearance Times in Cattle. J. Vet. Med. A 1996, 43, 93–101. [Google Scholar] [CrossRef]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef] [PubMed]
- Abdela, N. Sub-acute Ruminal Acidosis (SARA) and its Consequence in Dairy Cattle: A Review of Past and Recent Research at Global Prospective. Achiev. Life Sci. 2016, 10, 187–196. [Google Scholar] [CrossRef]
- Tian, M.; Fan, J.; Zhuang, Z.; Dai, F.; Wang, C.; Hou, H.; Ma, Y. Effects of silymarin on p65 NF-κB, p38 MAPK and CYP450 in LPS-induced hoof dermal inflammatory cells of dairy cows. BMC Vet. Res. 2019, 15, 127. [Google Scholar] [CrossRef] [PubMed]
- Pollitt, C.C. Basement membrane pathology: A feature of acute equine laminitis. Equine Vet. J. 1996, 28, 38–46. [Google Scholar] [CrossRef]
- Reisinger, N.; Schaumberger, S.; Nagl, V.; Hessenberger, S.; Schatzmayr, G. Concentration Dependent Influence of Lipopolysaccharides on Separation of Hoof Explants and Supernatant Lactic Acid Concentration in an Ex Vivo/In Vitro Laminitis Model. PLoS ONE 2015, 10, e0143754. [Google Scholar] [CrossRef]
- Reisinger, N.; Schaumberger, S.; Nagl, V.; Hessenberger, S. Milk Thistle Extract and Silymarin Inhibit Lipopolysaccharide Induced Lamellar Separation of Hoof Explants In Vitro. Toxins 2014, 6, 2962–2974. [Google Scholar] [CrossRef] [PubMed]
- Reisinger, N.; Dohnal, I.; Nagl, V.; Schaumberger, S.; Schatzmayr, G.; Mayer, E. Fumonisin B₁ (FB₁) Induces Lamellar Separation and Alters Sphingolipid Metabolism of In Vitro Cultured Hoof Explants. Toxins 2016, 8, 89. [Google Scholar] [CrossRef]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]
- Baldwin, R.L.; Jesse, B. Technical note: Isolation and characterization of sheep ruminal epithelial cells. J. Anim. Sci. 1991, 69, 3603–3609. [Google Scholar] [CrossRef]
- Lu, Z.; Gui, H.; Yao, L.; Yan, L.; Martens, H.; Aschenbach, J.R.; Shen, Z. Short-chain fatty acids and acidic pH upregulate UT-B, GPR41, and GPR4 in rumen epithelial cells of goats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Stumpff, F.; Georgi, M.; Mundhenk, L.; Rabbani, I.; Fromm, M.; Martens, H.; Günzel, D. Sheep rumen and omasum primary cultures and source epithelia: Barrier function aligns with expression of tight junction proteins. J. Exp. Biol. 2011, 214, 2871–2882. [Google Scholar] [CrossRef]
- Loret, S.; Rusu, D.; El, B.; Taminiau, B.; Heinen, E.; Dandrifosse, G.; Mainil, J. Preliminary characterization of jejunocyte and colonocyte cell lines isolated by enzymatic digestion from adult and young cattle. Res. Vet. Sci. 2009, 87, 123–132. [Google Scholar] [CrossRef]
- Reisinger, N.; Emsenhuber, C.; Doupovec, B.; Mayer, E.; Schatzmayr, G.; Nagl, V.; Grenier, B. Endotoxin Translocation and Gut Inflammation Are Increased in Broiler Chickens Receiving an Oral Lipopolysaccharide (LPS) Bolus during Heat Stress. Toxins 2020, 12, 622. [Google Scholar] [CrossRef] [PubMed]
- Lichte, P.; Grigoleit, J.S.; Steiner, E.M.; Kullmann, J.S.; Schedlowski, M.; Oberbeck, R.; Kobbe, P. Low dose LPS does not increase TLR4 expression on monocytes in a human in vivo model. Cytokine 2013, 63, 74–80. [Google Scholar] [CrossRef]
- McCann, J.C.; Luan, S.; Cardoso, F.C.; Derakhshani, H.; Khafipour, E.; Loor, J.J.; Petri, R.M. Induction of Subacute Ruminal Acidosis Affects the Ruminal Microbiome and Epithelium. Front. Microbiol. 2016, 7, 701. [Google Scholar] [CrossRef]
- Liu, J.; Bian, G.; Zhu, W.; Mao, S. High-grain feeding causes strong shifts in ruminal epithelial bacterial community and expression of Toll-like receptor genes in goats. Front. Microbiol. 2015, 6, 167. [Google Scholar] [CrossRef] [PubMed]
- Minuti, A.; Palladino, A.; Khan, M.J.; Alqarni, S.; Agrawal, A.; Piccioli-Capelli, F.; Hidalgo, F.; Cardoso, F.C.; Trevisi, E.; Loor, J.J. Abundance of ruminal bacteria, epithelial gene expression, and systemic biomarkers of metabolism and inflammation are altered during the peripartal period in dairy cows. J. Dairy Sci. 2015, 98, 8940–8951. [Google Scholar] [CrossRef]
- Bach, A.; Guasch, I.; Elcoso, G.; Castex, M.; Fàbregas, F. Changes in gene expression in the rumen and colon epithelia during the dry period through lactation of dairy cows and effects of live yeast supplementation. J. Dairy Sci. 2018, 101, 2631–2640. [Google Scholar] [CrossRef]
- Chen, Y.; Oba, M.; Guan, L.L. Variation of bacterial communities and expression of Toll-like receptor genes in the rumen of steers differing in susceptibility to subacute ruminal acidosis. Vet. Microbiol. 2012, 159, 451–459. [Google Scholar] [CrossRef]
- Yang, C.; Lan, W.; Ye, S.; Zhu, B.; Fu, Z. Transcriptomic Analyses Reveal the Protective Immune Regulation of Conjugated Linoleic Acids in Sheep Ruminal Epithelial Cells. Front. Physiol. 2020, 11, 588082. [Google Scholar] [CrossRef]
- Fingleton, B. Matrix metalloproteinases as regulators of inflammatory processes. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2036–2042. [Google Scholar] [CrossRef] [PubMed]
- Medina, C.; Radomski, M.W. Role of Matrix Metalloproteinases in Intestinal Inflammation. J. Pharmacol. Exp. Ther. 2014, 318, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Nighot, P.; Al-Sadi, R.; Rawat, M.; Guo, S.; Watterson, D.M.; Ma, T. Matrix metalloproteinase 9-induced increase in intestinal epithelial tight junction permeability contributes to the severity of experimental DSS colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 309, 988–997. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Youssef, M.; Rawat, M.; Guo, S.; Dokladny, K.; Haque, M.; Watterson, M.D.; Ma, T.Y.; Ty, M. MMP-9-induced increase in intestinal epithelial tight permeability is mediated by p38 kinase signaling pathway activation of MLCK gene. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, 278–290. [Google Scholar] [CrossRef]
- Li, H.; Zheng, H.; Li, L.; Shen, X.; Zang, W.; Sun, Y. The Effects of Matrix Metalloproteinase-9 on Dairy Goat Mastitis and Cell Survival of Goat Mammary Epithelial Cells. PLoS ONE 2016, 11, e0160989. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shen, Y.; Li, K.; Zhang, P.; Wang, G.; Gao, L.; Bai, C.; Elisa, M. Role of matrix metalloproteinase-9 in lipopolysaccharide-induced mucin production in human airway epithelial cells. Arch. Biochem. Biophys. 2009, 486, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hao, X.; Sun, B. Lipopolysaccharide regulates MMP-9 expression through TLR4/NF-κB signaling in human arterial smooth muscle cells. Mol. Med. Rep. 2012, 6, 774–778. [Google Scholar] [CrossRef]
- Wong, Y.; Sethu, C.; Louafi, F.; Hossain, P. Lipopolysaccharide Regulation of Toll-Like Receptor-4 and Matrix Metalloprotease-9 in Human Primary Corneal Fibroblasts. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2796–2803. [Google Scholar] [CrossRef][Green Version]
- Muthukuru, M.; Cutler, C.W. Resistance of MMP9 and TIMP1 to endotoxin tolerance. Pathog. Dis. 2019, 73, ftu003. [Google Scholar] [CrossRef]
- Dai, H.; Liu, X.; Yan, J.; Aabdin, Z.; Bilal, M.S.; Shen, X. Sodium Butyrate Ameliorates High-Concentrate Diet- Induced Inflammation in the Rumen Epithelium of Dairy Goats. J. Agric. Food Chem. 2017, 65, 596–604. [Google Scholar] [CrossRef]
- Wu, Y.; Sun, Y.; Zhang, Z.; Chen, J.; Dong, G. Effects of Peptidoglycan, Lipoteichoic Acid and Lipopolysaccharide on Inflammation, Proliferation and Milk Fat Synthesis in Bovine Mammary Epithelial Cells. Toxins 2020, 12, 497. [Google Scholar] [CrossRef]
- Bulgari, O.; Dong, X.; Roca, A.L.; Caroli, A.M.; Loor, J.J. Innate immune responses induced by lipopolysaccharide and lipoteichoic acid in primary goat mammary epithelial cells. J. Anim. Sci. Biotechnol. 2017, 8, 29. [Google Scholar] [CrossRef]
- Günther, J.; Esch, K.; Poschadel, N.; Petzl, W.; Zerbe, H.; Mitterhuemer, S.; Blum, H.; Seyfert, H. Comparative Kinetics of Escherichia coli- and Staphylococcus aureus- Specific Activation of Key Immune Pathways in Mammary Epithelial Cells Demonstrates That S. aureus Elicits a Delayed Response Dominated by Interleukin-6 (IL-6) but Not by IL-1A or tumor necrosis factor alpha. Infect. Immun. 2011, 79, 695–707. [Google Scholar] [PubMed]
- Tsugami, Y.; Wakasa, H.; Kawahara, M.; Nishimura, T. Lipopolysaccharide and lipoteichoic acid influence milk production ability via different early responses in bovine mammary epithelial cells. Exp. Cell Res. 2021, 400, 112472. [Google Scholar] [CrossRef]
- Petzl, W.; Zerbe, H.; Yang, W. Escherichia coli, but not Staphylococcus aureus triggers an early increased expression of factors contributing to the innate immune defense in the udder of the cow. Vet. Res. 2014, 39, 651–668. [Google Scholar]
- Wellnitz, O.; Arnold, E.T.; Bruckmaier, R.M. Lipopolysaccharide and lipoteichoic acid induce different immune responses in the bovine mammary gland. J. Dairy Sci. 2011, 94, 5405–5412. [Google Scholar] [CrossRef] [PubMed]
- Elena, A.; Giovannini, J.; Henricus, B.; Van Den Borne, P.; Wall, S.K.; Wellnitz, O.; Bruckmaier, R.M.; Spadavecchia, C. Experimentally induced subclinical mastitis: Are lipopolysaccharide and lipoteichoic acid eliciting similar pain responses? Acta Vet. Scand. 2017, 59, 40. [Google Scholar]
- Nocek, J.E. Bovine Acidosis: Implications on Laminitis. J. Dairy Sci. 1997, 80, 1005–1028. [Google Scholar] [CrossRef]
- Singh, S.S.; Murray, R.D.; Ward, W.R. Gross and Histopathological Study of Endotoxin-induced Hoof Lesions in Cattle. J. Comp. Pathol. 1994, 110, 103–115. [Google Scholar] [CrossRef]
- Leise, B.S.; Yin, C.; Pettigrew, A.; Belknap, J.K. Proinflammatory cytokine responses of cultured equine keratinocytes to bacterial pathogen-associated molecular pattern motifs. Equine Vet. J. 2010, 42, 294–303. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | 1 h | 2 h | 4 h | 24 h | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | LPS Low | LPS High | Control | LPS Low | LPS High | Control | LPS Low | LPS High | Control | LPS Low | LPS High | |
| TLR2 | 1.00 | −1.27 | −1.23 | 1.00 | 1.26 | 1.51 | 1.00 | 1.45 | 1.32 | 1.00 | 1.36 | 1.24 |
| TLR4 | 1.00 | −1.34 | −1.20 | 1.00 | −1.05 | −1.18 | 1.00 | 1.43 | 1.17 | 1.00 | 1.26 | 1.07 |
| MyD88 | 1.00 | 1.08 | 1.02 | 1.00 | 1.16 | −1.10 | 1.00 | 1.05 | −1.03 | 1.00 | 1.09 | 1.07 |
| TRAF6 | 1.00 | −1.20 | −1.24 | 1.00 | 1.01 | 1.02 | 1.00 | 1.06 | 1.03 | 1.00 | 1.01 | 1.02 |
| NFKB1 | 1.00 | 1.09 | 1.02 | 1.00 a | 2.39 b | 2.13 b | 1.00 a | 2.34 b | 2.23 b | 1.00 a | 2.09 b | 2.04 b |
| TNF-α | 1.00 a | 12.92 b | 8.04 ab | 1.00 a | 106.03 b | 87.13 b | 1.00 a | 8.11 b | 7.33 b | 1.00 a | 6.33 b | 7.63 b |
| IL1B | 1.00 | 1.90 | −1.05 | 1.00 a | 43.48 b | 46.93 b | 1.00 a | 96.98 b | 113.43 b | 1.00 a | 16.75 ab | 38.03 b |
| IL6 | 1.00 | 2.31 | 2.61 | 1.00 a | 26.55 b | 22.71 b | 1.00 a | 10.73 b | 10.14 b | 1.00 a | 6.75 b | 6.97 b |
| CXCL8 | 1.00 a | 24.87 b | 21.99 b | 1.00 a | 187.90 b | 158.33 b | 1.00 a | 29.47 b | 37.00 b | 1.00 a | 34.49 b | 42.50 b |
| MMP2 | 1.00 | 1.15 | 1.15 | 1.00 | 1.05 | −1.00 | 1.00 | 1.04 | 1.17 | 1.00 | 1.02 | −1.00 |
| MMP9 | 1.00 | 1.10 | −1.11 | 1.00 | 2.59 | 2.20 | 1.00 | 1.45 | 1.32 | 1.00 a | 14.32 b | 13.77 b |
| Genes | 1 h | 2 h | 4 h | 24 h | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | LTA Low | LTA High | Control | LTA Low | LTA High | Control | LTA Low | LTA High | Control | LTA Low | LTA High | |
| TLR2 | 1.00 | 1.34 | −1.23 | 1.00 | 1.01 | −1.08 | 1.00 | 1.26 | 1.13 | 1.00 | 1.20 | 1.75 |
| TLR4 | 1.00 | −1.23 | −1.10 | 1.00 | −1.03 | −1.17 | 1.00 | 1.01 | −1.02 | 1.00 | 1.01 | 1.17 |
| MyD88 | 1.00 | 1.06 | 1.03 | 1.00 | 1.05 | 1.05 | 1.00 | 1.01 | −1.01 | 1.00 | −1.05 | 1.03 |
| TRAF6 | 1.00 | −1.11 | −1.05 | 1.00 | −1.03 | −1.01 | 1.00 | −1.00 | 1.01 | 1.00 | 1.00 | −1.01 |
| NFKB1 | 1.00 | 1.01 | 1.01 | 1.00 | 1.57 | 2.06 | 1.00 | 1.15 | 1.27 | 1.00 | 1.21 | 1.52 |
| TNF-α | 1.00 | −1.24 | −1.28 | 1.00 a | 4.23 b | 14.45 b | 1.00 a | 3.93 b | 11.73 c | 1.00 a | 1.37 a | 2.86 b |
| IL1B | 1.00 | −1.02 | −1.05 | 1.00 a | 19.56 b | 60.72 b | 1.00 a | 1.36 ab | 2.95 b | 1.00 a | 1.28 a | 20.8 b |
| IL6 | 1.00 | 1.44 | 1.61 | 1.00 a | 2.72 a | 5.53 b | 1.00 | 1.01 | −1.02 | 1.00 | 1.05 | 1.47 |
| CXCL8 | 1.00 | 1.14 | 1.34 | 1.00 a | 12.67 a | 54.67 b | 1.00 | 1.26 | 1.13 | 1.00 a | 1.80 a | 7.07 b |
| MMP2 | 1.00 | 1.13 | 1.14 | 1.00 | −1.02 | 1.00 | 1.00 | −1.00 | 1.01 | 1.00 | 1.15 | 1.11 |
| MMP9 | 1.00 | −1.06 | 1.16 | 1.00 | 1.05 | 1.45 | 1.00 | 1.17 | −1.06 | 1.00 a | 2.57 b | 6.01 b |
| Genes | Start 0 h | Control 4 h | 1 µg/mL LPS 4 h | 5 µg/mL LPS 4 h | 10 µg/mL LPS 4 h |
|---|---|---|---|---|---|
| TLR2 | 1.00 a | 1.51 a | 2.52 b | 3.39 b | 3.0 b |
| TLR4 | 1.00 | −1.05 | −1.25 | 1.17 | 1.03 |
| MyD88 | 1.00 | −1.27 | −1.47 | −1.45 | −1.39 |
| TRAF6 | 1.00 | −1.14 | −1.28 | 1.00 | −1.05 |
| NFKB1 | 1.00 a | 2.83 b | 2.73 b | 4.13 b | 3.55 b |
| TNF-α | 1.00 a | 2.79 b | 1.31a | 4.70 b | 4.65 b |
| IL1B | 1.00 a | 110.92 b | 130.86 b | 197.04 b | 150.52 b |
| IL6 | 1.00 a | 117.49 b | 159.27 b | 255.75 b | 144.44 b |
| CXCL8 | 1.00 a | 68.71 b | 91.30 b | 162.57 b | 115.19 b |
| MMP2 | 1.00 | −1.25 | −1.64 | −1.19 | −1.25 |
| MMP9 | 1.00 | 1.97 | 3.89 | 1.77 | 3.72 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reisinger, N.; Wendner, D.; Schauerhuber, N.; Mayer, E. Effect of Lipopolysaccharides (LPS) and Lipoteichoic Acid (LTA) on the Inflammatory Response in Rumen Epithelial Cells (REC) and the Impact of LPS on Claw Explants. Animals 2021, 11, 2058. https://doi.org/10.3390/ani11072058
Reisinger N, Wendner D, Schauerhuber N, Mayer E. Effect of Lipopolysaccharides (LPS) and Lipoteichoic Acid (LTA) on the Inflammatory Response in Rumen Epithelial Cells (REC) and the Impact of LPS on Claw Explants. Animals. 2021; 11(7):2058. https://doi.org/10.3390/ani11072058
Chicago/Turabian StyleReisinger, Nicole, Dominik Wendner, Nora Schauerhuber, and Elisabeth Mayer. 2021. "Effect of Lipopolysaccharides (LPS) and Lipoteichoic Acid (LTA) on the Inflammatory Response in Rumen Epithelial Cells (REC) and the Impact of LPS on Claw Explants" Animals 11, no. 7: 2058. https://doi.org/10.3390/ani11072058
APA StyleReisinger, N., Wendner, D., Schauerhuber, N., & Mayer, E. (2021). Effect of Lipopolysaccharides (LPS) and Lipoteichoic Acid (LTA) on the Inflammatory Response in Rumen Epithelial Cells (REC) and the Impact of LPS on Claw Explants. Animals, 11(7), 2058. https://doi.org/10.3390/ani11072058