
Response of Reeves’s Pheasants Distribution to Human Infrastructure in the Dabie Mountains over the Last 20 Years





Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
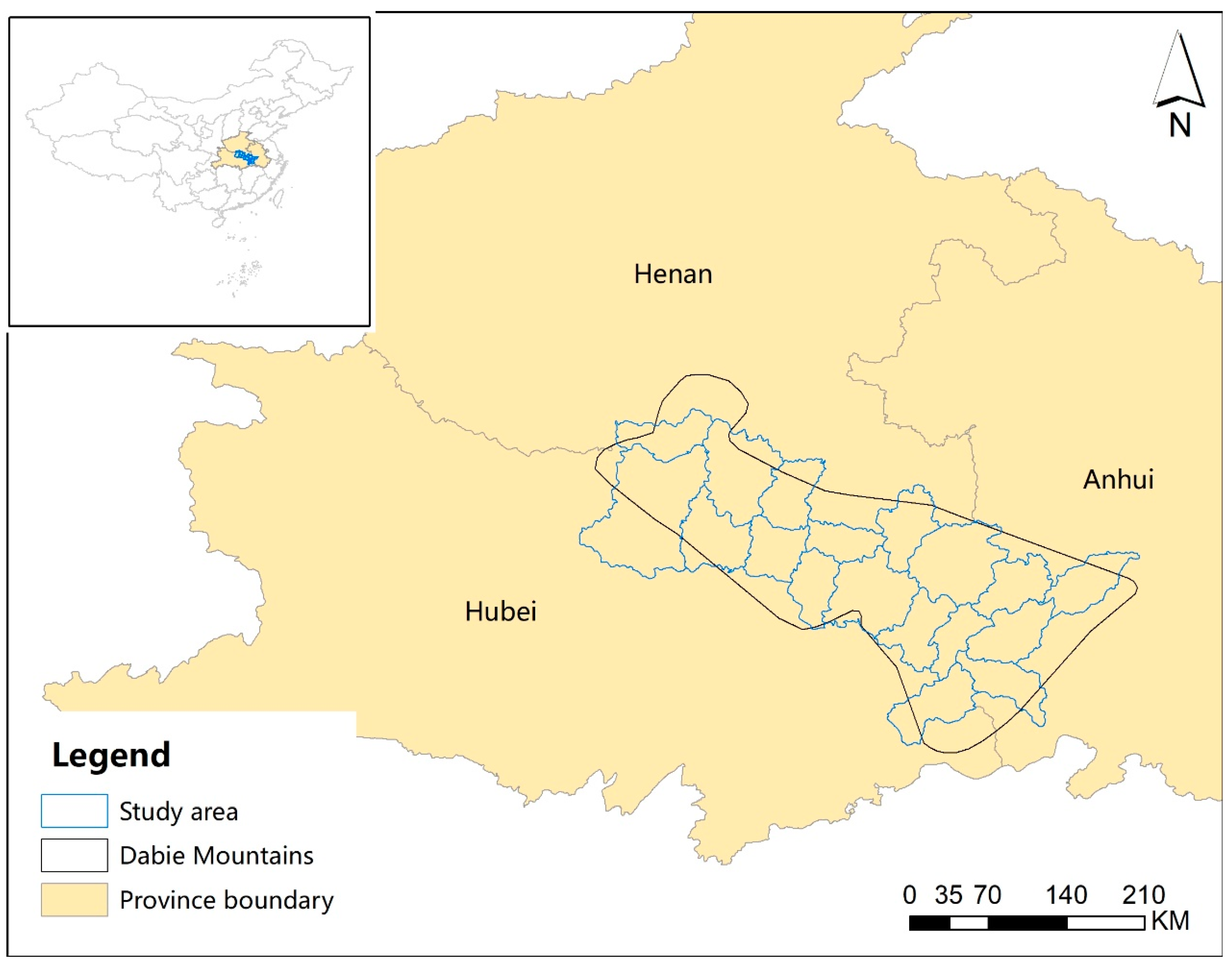
2.1. Study Area and Data Collection
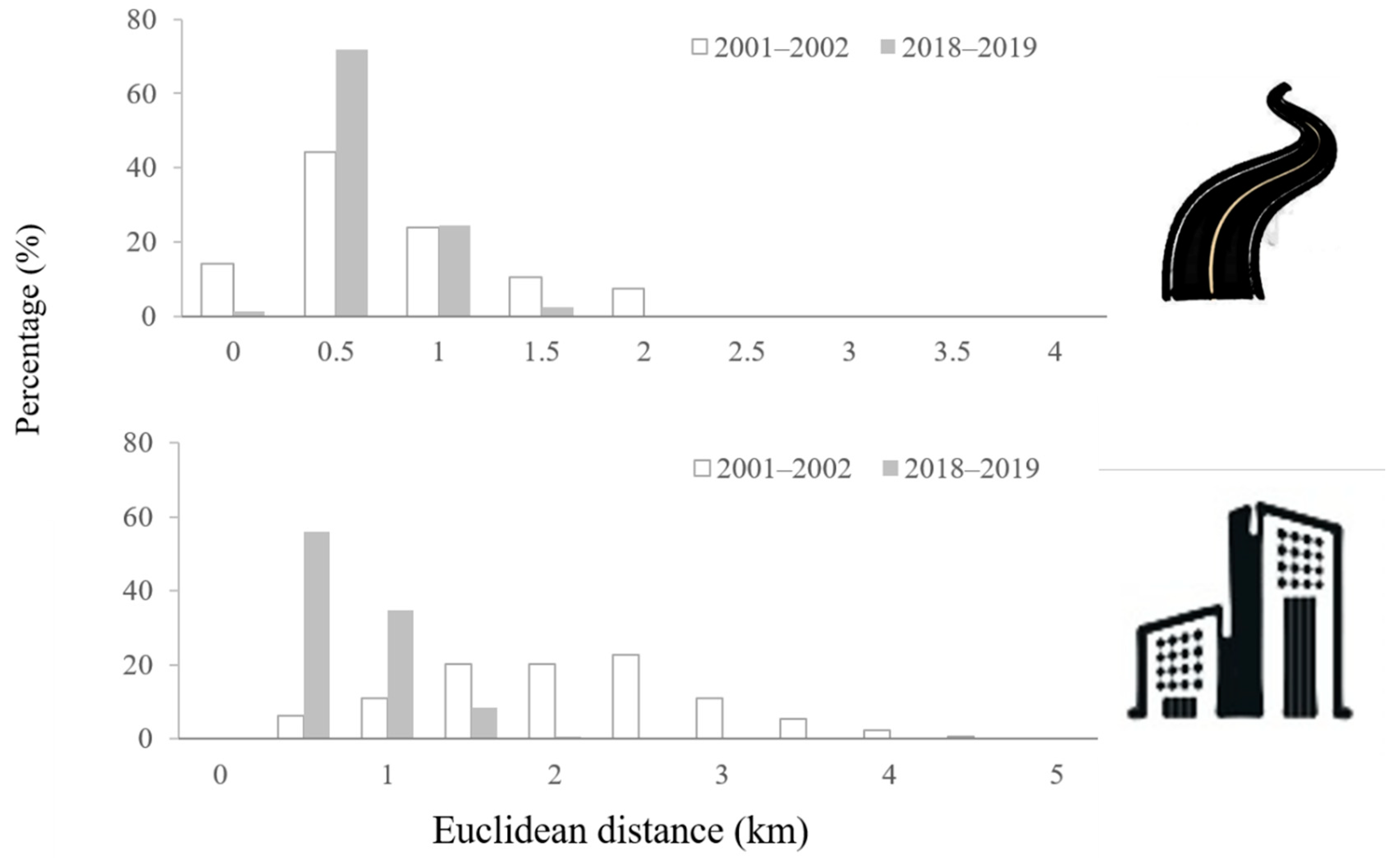
2.2. Euclidean Distance Analysis for Regional Infrastructure
2.3. Spatiotemporal Effects of Infrastructure on Reeves’s Pheasants
2.4. Data Analysis
3. Results
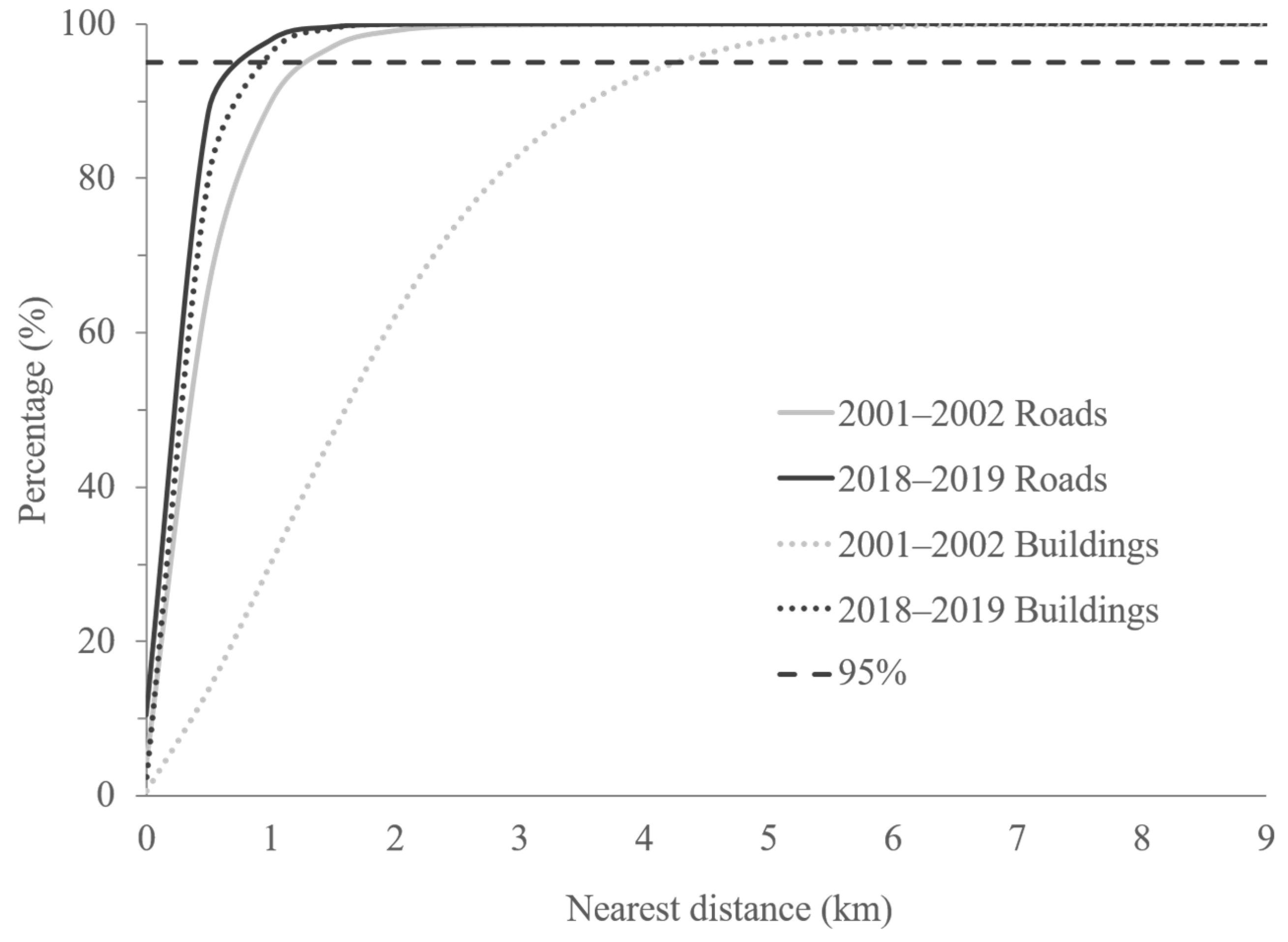
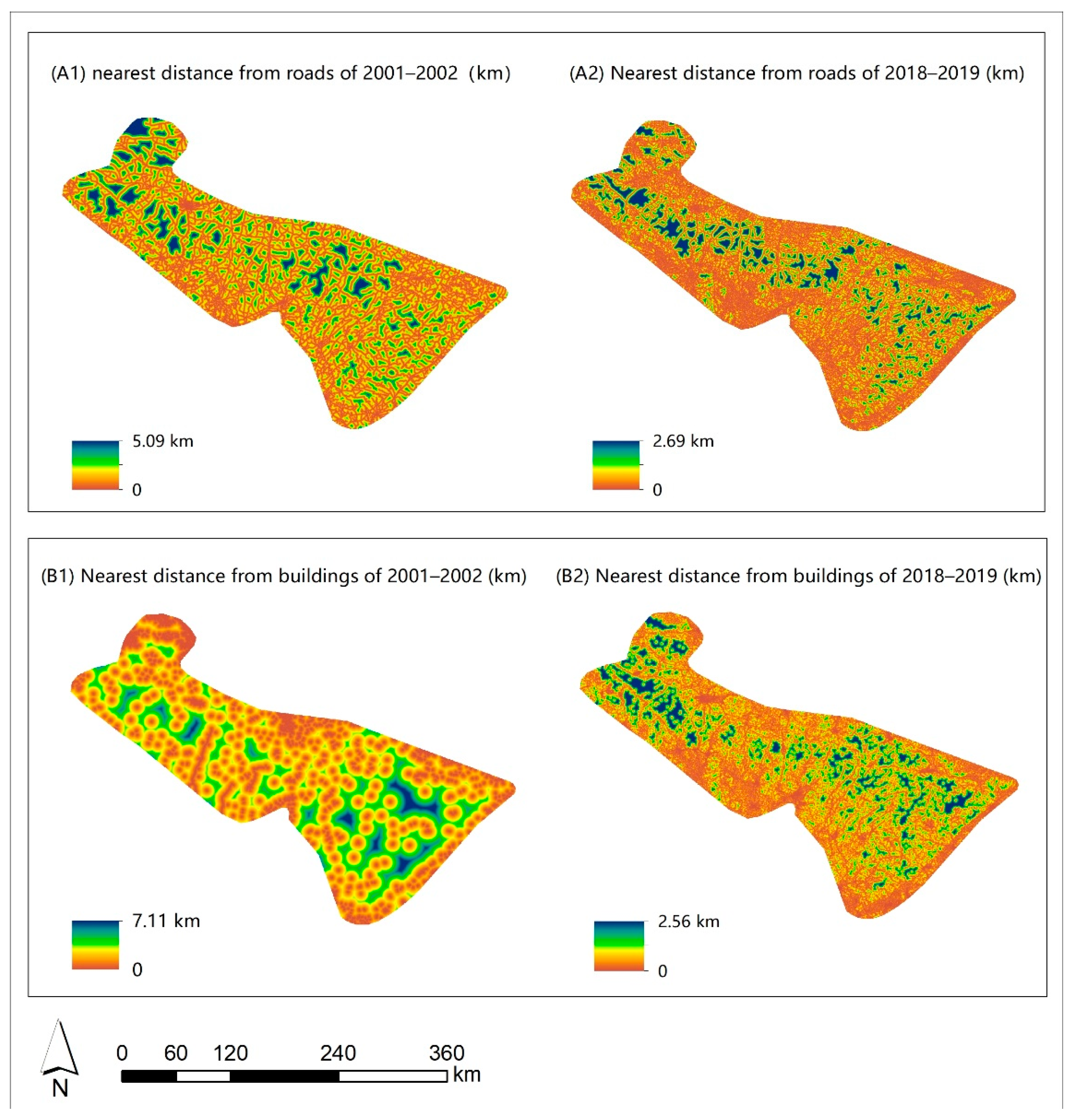
3.1. Distance Maps for Roads and Buildings
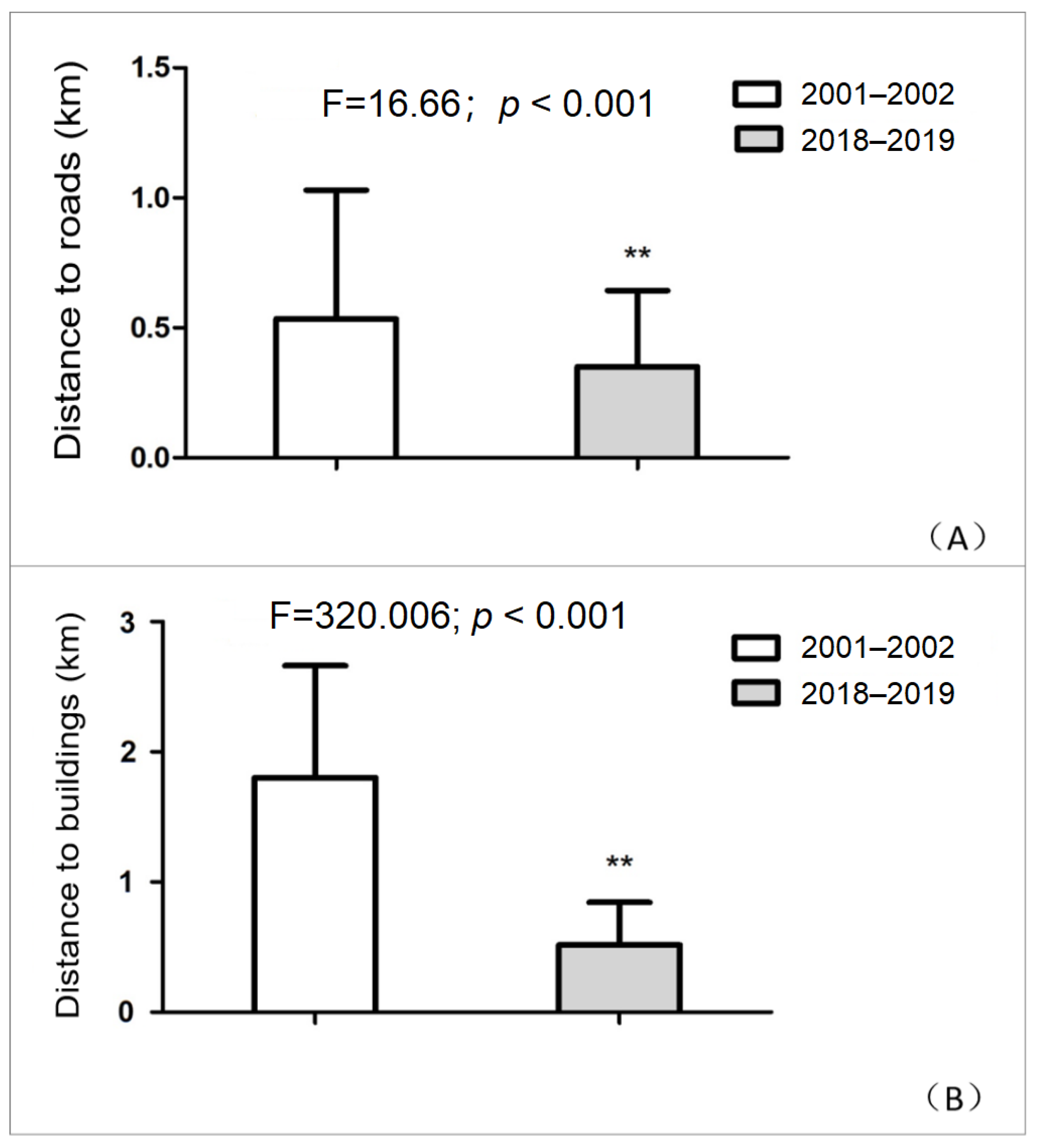
3.2. Spatiotemporal Effects of Infrastructure on Reeves’s Pheasants
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sih, A.; Ferrari, M.C.; Harris, D.J. Evolution and behavioural responses to human-induced rapid environmental change. Evol. Appl. 2011, 4, 367–387. [Google Scholar] [CrossRef]
- Torres, A.; Jaeger, J.A.G.; Alonso, J.C. Assessing large-scale wildlife responses to human infrastructure development. Proc. Natl. Acad. Sci. USA 2016, 113, 8472–8477. [Google Scholar] [CrossRef] [Green Version]
- Balotari-Chiebao, F.; Brommer, J.E.; Tikkanen, H.; Laaksonen, T. Habitat use by post-fledging white-tailed eagles shows avoidance of human infrastructure and agricultural areas. Eur. J. Wildl. Res. 2021, 67, 1–7. [Google Scholar] [CrossRef]
- Fernández-Juricic, E.; Venier, M.P.; Renison, D.; Blumstein, D.T. Sensitivity of wildlife to spatial patterns of recreationist behavior: A critical assessment of minimum approaching distances and buffer areas for grassland birds. Biol. Conserv. 2005, 125, 225–235. [Google Scholar] [CrossRef]
- Ahmed, S.E.; Lees, A.C.; Moura, N.G.; Gardner, T.A.; Barlow, J.; Ferreira, J.; Ewers, R.M. Road networks predict human influence on Amazonian bird communities. Proc. R. Soc. B 2014, 281, 20141742. [Google Scholar] [CrossRef] [Green Version]
- Konstantopoulos, K.; Moustakas, A.; Vogiatzakis, I.N. A spatially explicit impact assessment of road characteristics, road-induced fragmentation and noise on bird species in Cyprus. Biol. Conserv. 2020, 252, 108828. [Google Scholar] [CrossRef] [Green Version]
- Jin, Z.; Wang, J.; Kong, X. Combining habitat area and fragmentation change for ecological disturbance assessment in Jiangsu Province, China. Environ. Sci. Pollut. Res. 2020, 27, 20817–20830. [Google Scholar] [CrossRef]
- Di Marco, M.; Venter, O.; Possingham, H.P.; Watson, J.E.M. Changes in human footprint drive changes in species extinction risk. Nat. Commun. 2018, 9, 4621. [Google Scholar] [CrossRef]
- Sanderson, E.W.; Jaiteh, M.L.M.A.; Redford, K.H.; Wannebo, A.V.; Woolmer, G. The human footprint and the last of the wild. BioScience 2002, 52, 891–904. [Google Scholar] [CrossRef]
- Seto, K.C.; Guneralp, B.; Hutyra, L.R. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc. Natl. Acad. Sci. USA 2012, 109, 16083–16088. [Google Scholar] [CrossRef] [Green Version]
- Huera-Lucero, T.; Salas-Ruiz, A.; Changoluisa, D.; Bravo-Medina, C. Towards sustainable urban planning for Puyo (Ecuador): Amazon forest landscape as potential green infrastructure. Sustainability 2020, 12, 4768. [Google Scholar] [CrossRef]
- Weinzettel, J.; Hertwich, E.G.; Peters, G.P.; Steen-Olsen, K.; Galli, A. Affluence drives the global displacement of land use. Global Environ. Chang. 2013, 23, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Yao, M.; Zhang, Y. Evaluation and optimization of urban land-use efficiency: A case study in Sichuan Province of China. Sustainability 2021, 13, 1771. [Google Scholar] [CrossRef]
- Bennett, V.J.; Beard, M.; Zollner, P.A.; Fernández-Juricic, E.; Westphal, L.; LeBlanc, C.L. Understanding wildlife responses to human disturbance through simulation modelling: A management tool. Ecol. Complex. 2009, 6, 113–134. [Google Scholar] [CrossRef]
- Lee, J.; Arts, J.; Vanclay, F.; Ward, J. Examining the social outcomes from urban transport infrastructure: Long-term consequences of spatial changes and varied interests at multiple levels. Sustainability 2020, 12, 5907. [Google Scholar] [CrossRef]
- van der Ree, R.; Jaeger, J.A.G.; van der Grift, E.A.; Clevenger, A.P. Effects of roads and traffic on wildlife populations and landscape function: Road ecology is moving toward larger scales. Ecol. Soc. 2011, 16, 48. [Google Scholar] [CrossRef]
- Fernández-Juricic, E.; Vaca, R.; Schroeder, N. Spatial and temporal responses of forest birds to human approaches in a protected area and implications for two management strategies. Biol. Conserv. 2004, 117, 407–416. [Google Scholar] [CrossRef]
- Williams, C.K.; Lutz, R.S.; Applegate, R.D. Winter survival and additive harvest in northern bobwhite coveys in Kansas. J. Wildl. Manage. 2004, 68, 94–100. [Google Scholar] [CrossRef]
- Jones, D.N.; Bernede, L.; Bond, A.R.F.; Dexter, C.; Strong, C.L. Dust as a contributor to the road-effect zone: A case study from a minor forest road in Australia. Aust. J. Environ. Manage. 2016, 23, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Zheng, G.M. Pheasants in China; Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Xu, J.; Zhang, Z.; Zheng, G.; Zhang, X.; Sun, Q.; McGowan, P. Home range and habitat use of Reeves’s Pheasant Syrmaticus reevesii in the protected areas created from forest farms in the Dabie Mountains, central China. Bird Conserv. Internat. 2007, 17, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Zhang, Z. Home range and habitat composition of male Reeves’s Pheasants in an agricultural-forest plantation landscape in central China: A preliminary report. Chin. Birds 2011, 2, 53–58. [Google Scholar] [CrossRef]
- Lu, S.; Li, J.; Tian, S.; Hua, J.; Xu, J. Effects of tourism on nest survival rate of Galliformes based on artificial nest experiment. Acta Ecol. Sin. 2020, 40, 1–10. [Google Scholar]
- Zhou, C.; Xu, J.; Zhang, Z. Dramatic decline of the vulnerable Reeves’s pheasant Syrmaticus reevesii, endemic to central China. Oryx 2015, 49, 529–534. [Google Scholar] [CrossRef] [Green Version]
- IUCN. Syrmaticus reevesii. The IUCN Red List of Threatened Species 2018: E.T22679346A131873938. Available online: https://www.iucnredlist.org/species/22679346/131873938 (accessed on 13 March 2020).
- Zhang, Z.W.; Ding, C.Q.; Ding, P.; Zheng, G.M. The current status and a conservation strategy for species of Galliformes in China. Biodivers. Sci. 2003, 11, 414–421. [Google Scholar]
- Tian, S.; Xu, J.; Wang, Y. Human infrastructure development drives decline in suitable habitat for Reeves’s pheasant in the Dabie Mountains in the last 20 years. Glob. Ecol. Conserv. 2020, 22, e00940. [Google Scholar] [CrossRef]
- CITES. Convention on International Trade in Endangered Species of Wild Fauna and Flora. In Proceedings of the Twenty-fourth meeting of the Animals Committee, Geneva, Switzerland, 20–24 April 2009. [Google Scholar]
- NFGA. List of National Key Protected Wild Animals; National Forestry and Grassland Administration: Beijing, China, 2021. [Google Scholar]
- Forman, R.T.; Sperling, D.; Bissonette, J.A.; Clevenger, A.P.; Cutshall, C.D.; Dale, V.H.; Jones, J. Road Ecology: Science and Solutions; Island Press: Washington, DC, USA, 2003. [Google Scholar]
- Zhou, C.; Zhao, Y.; Connelly, J.W.; Li, J.; Xu, J. Current nature reserve management in China and effective conservation of threatened pheasant species. Wildl. Biol. 2017, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Grainger, M.J.; Garson, P.J.; Browne, S.J.; McGowan, P.J.K.; Savini, T. Conservation status of Phasianidae in southeast Asia. Biol. Conserv. 2018, 220, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Mateo, J.; Romero, M.; Urios, V. How can the home range of the Lesser Kestrel be affected by a large civil infrastructure? Chin. Birds 2019, 10, 138–147. [Google Scholar] [CrossRef]
- Chen, W.; Xie, Z.; Zhou, Y. Proximity to roads reduces acorn dispersal effectiveness by rodents: Implication for forest regeneration and management. For. Ecol. Manage. 2019, 433, 625–632. [Google Scholar] [CrossRef]
- Xu, W.; Xiao, Y.; Zhang, J.; Yang, W.; Zhang, L.; Hull, V.; Wang, Z.; Zheng, H.; Liu, J.; Polasky, S.; et al. Strengthening protected areas for biodiversity and ecosystem services in China. Proc. Natl. Acad. Sci. USA 2017, 114, 1601–1606. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Pimm, S.L.; Du, A.; Su, Y.; Fan, X.; An, L.; Liu, J.; Ouyang, Z. Transforming protected area management in China. Trends Ecol. Evol. 2019, 34, 762–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, S.; Lu, S.; Hua, J.; Chang, J.; Li, J.; Zhang, Z.; Wang, Y.; Xu, J.; Zhang, M. Integrating habitat suitability modelling and assessment of the conservation gaps of nature reserves for the threatened Reeves’s Pheasant. Bird Conserv. Internat. 2021, First View, 1–14. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Variables | Coefficients ± SE | df | F | p |
|---|---|---|---|---|---|
| 2001–2002 | Distance to nearest road | 1.274 ± 0.984 | 1, 321 | 1.19 | 0.196 |
| Distance to nearest building | −0.038 ± 0.281 | 1, 321 | 0.02 | 0.893 | |
| Distance to road × distance to building | 0.326 ± 0.445 | 1, 321 | 0.54 | 0.464 | |
| Coefficients±SE | Z | p | |||
| Random effect: county (city) | 4.794 ± 1.823 | 2.63 | 0.009 | ||
| 2018–2019 | Distance to nearest road | 1.664 ± 0.868 | 1, 324 | 3.68 | 0.056 |
| Distance to nearest building | 3.837 ± 0.754 | 1, 324 | 25.93 | < 0.001 | |
| Distance to road × Distance to building | −2.350 ± 0.128 | 1, 324 | 3.37 | 0.067 | |
| Coefficients±SE | Z | p | |||
| Random effect: county (city) | 1.545 ± 0.689 | 2.24 | 0.025 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, S.; Xu, J.; Li, J.; Zhang, M.; Wang, Y. Response of Reeves’s Pheasants Distribution to Human Infrastructure in the Dabie Mountains over the Last 20 Years. Animals 2021, 11, 2037. https://doi.org/10.3390/ani11072037
Tian S, Xu J, Li J, Zhang M, Wang Y. Response of Reeves’s Pheasants Distribution to Human Infrastructure in the Dabie Mountains over the Last 20 Years. Animals. 2021; 11(7):2037. https://doi.org/10.3390/ani11072037
Chicago/Turabian StyleTian, Shan, Jiliang Xu, Jianqiang Li, Mingxiang Zhang, and Yong Wang. 2021. "Response of Reeves’s Pheasants Distribution to Human Infrastructure in the Dabie Mountains over the Last 20 Years" Animals 11, no. 7: 2037. https://doi.org/10.3390/ani11072037
APA StyleTian, S., Xu, J., Li, J., Zhang, M., & Wang, Y. (2021). Response of Reeves’s Pheasants Distribution to Human Infrastructure in the Dabie Mountains over the Last 20 Years. Animals, 11(7), 2037. https://doi.org/10.3390/ani11072037